
Archaeogenetics and Landscape Dynamics in Sicily during the Holocene: A Review



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Ethnic and Demographic Dynamics of Sicily: From Prehistory to History
- (i)
- The application of genomic technologies to the analysis of ancient samples and extant populations;
- (ii)
- The selection of samples based on the careful integration of data from the material culture, literary sources, linguistics, and historical demography;
- (iii)
- The ascertainment of the place of birth of ancestors of blood/DNA donors from a given rural site (village, town) for at least three generations;
- (iv)
- The use of robust statistical techniques for data analysis.
3. The Archaeogenetics of Sicily: A Long Journey Lasted Forty Years
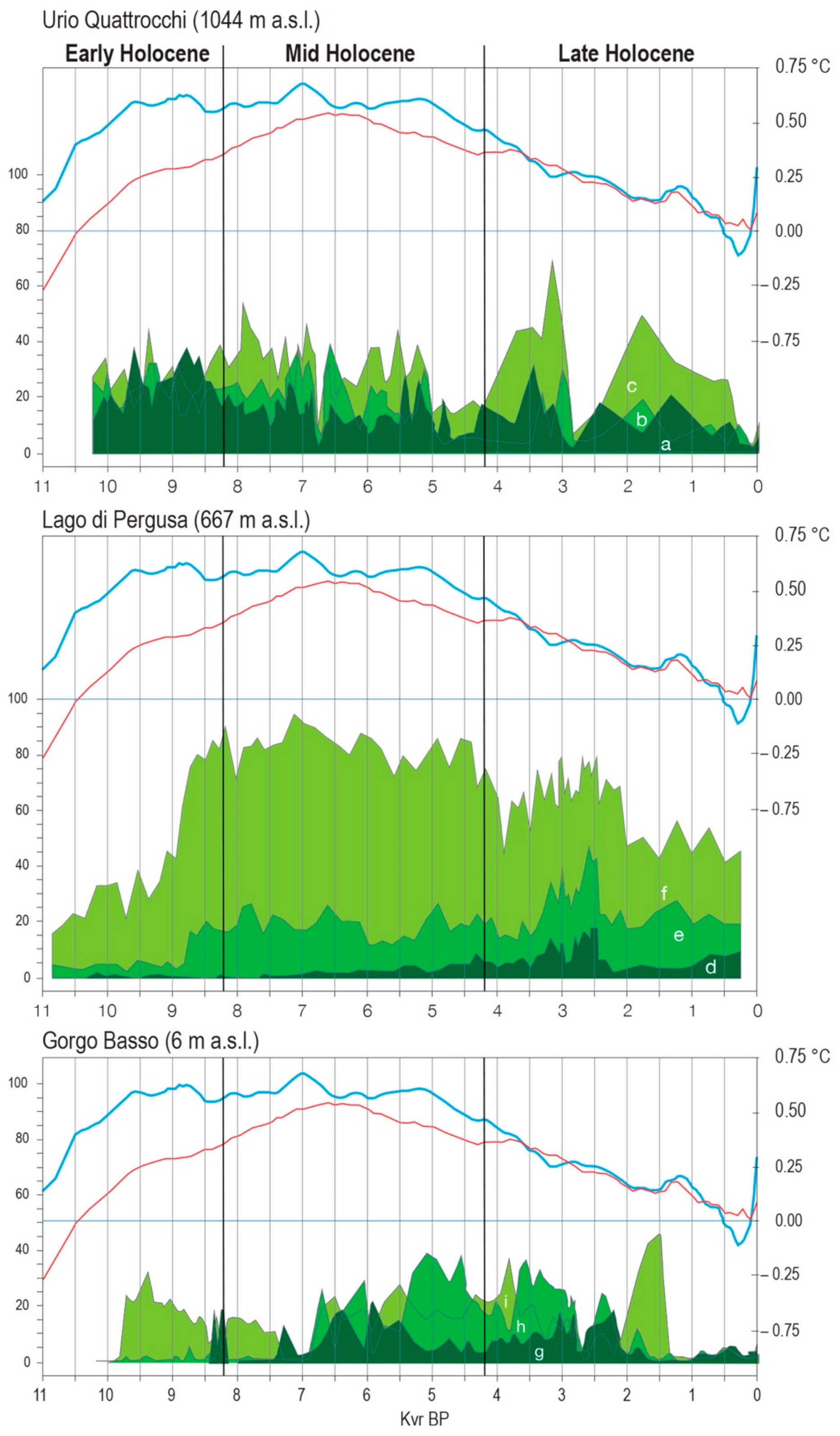
4. Past Vegetation, Climate and Landscape Dynamics during the Holocene

5. Discussion
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eisenmann, S.; Bánffy, E.; van Dommelen, P.; Hofmann, K.P.; Maran, J.; Lazaridis, I.; Mittnik, A.; McCormick, M.; Krause, J.; Reich, D.; et al. Reconciling material cultures in archaeology with genetic data: The nomenclature of clusters emerging from archaeogenomic analysis. Sci. Rep. 2018, 8, 13003. [Google Scholar] [CrossRef]
- Mannino, M.A.; Di Salvo, R.; Schimmenti, V.; Di Patti, C.; Incarbona, A.; Sineo, L.; Richards, M.P. Upper Palaeolithic hunter-gatherer subsistence in Mediterranean coastal environments: An isotopic study of the diets of the earliest directly-dated humans from Sicily. J. Archaeol. Sci. 2011, 38, 3094–3100. [Google Scholar] [CrossRef]
- Finley, M.I. A History of Sicily, Vol 1: Ancient Sicily to the Arab Conquest; Chatto & Windus: London, UK, 1968; p. 222. [Google Scholar]
- Romano, V.; Ayala, G.F. Genetic and Population history of Sicily. J. Cult. Herit. 2000, 1 (Suppl. S2), 1–2. [Google Scholar] [CrossRef]
- Crumley, C.L. Historical Ecology: A Robust Bridge between Archaeology and Ecology. Sustainability 2021, 13, 8210. [Google Scholar] [CrossRef]
- Sineo, L.; Petruso, D.; Forgia, V.; Messina, A.D.; D’Amore, G. Human Peopling of Sicily During Quaternary. In Geological Epochs; Fernandez, L.D., Ed.; AcademyPublish.org: Cheyenne, WY, USA, 2015; pp. 25–68. [Google Scholar]
- Antonioli, F.; Lo Presti, V.; Morticelli, M.G.; Bonfiglio, L.; Mannino, M.A.; Palombo, M.R.; Sannino, G.; Ferranti, L.; Furlani, S.; Lambeck, K.; et al. Timing of the emergence of the Europe-Sicily bridge (40–17 cal ka BP) and its implications for the spread of modern humans. Geol. Soc. Spec. Publ. 2016, 411, 111–144. [Google Scholar] [CrossRef]
- D’Amore, G.; Di Marco, S.; Tartarelli, G.; Bigazzi, R.; Sineo, L. Late Pleistocene human evolution in Sicily: Comparative morphometric analysis of Grotta di San Teodoro craniofacial remains. J. Hum. Evol. 2009, 56, 537–550. [Google Scholar] [CrossRef]
- Galland, M.; D’Amore, G.; Friess, M.; Miccichè, R.; Pinhasi, R.; Sparacello, V.S.; Sineo, L. Morphological variability of Upper Paleolithic and Mesolithic skulls from Sicily. J. Anthr. Sci. 2019, 96, 151–172. [Google Scholar] [CrossRef]
- Mathieson, I.; Songül, A.R.; Posth, C.; Szécsényi-Nagy, A.; Rohland, N.; Mallick, S.; Olalde, I.; Broomandkhoshbacht, N.; Candilio, F.; Cheronet, O.; et al. The genomic history of southeastern Europe. Nature 2018, 555, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Catalano, G.; Lo Vetro, D.; Fabbri, P.F.; Mallick, S.; Reich, D.; Rohland, N.; Sineo, L.; Mathieson, I.; Martini, F. Late upper palaeolithic hunter-gatherers in the central mediterranean: New archaeological and genetic data from the late epigravettian burial Oriente C (Favignana, Sicily). Quat. Int. 2020, 537, 24–32. [Google Scholar] [CrossRef]
- Modi, A.; Catalano, G.; D’Amore, G.; Di Marco, S.; Lari, M.; Sineo, L.; Caramelli, D. Paleogenetic and morphometric analysis of a Mesolithic individual from Grotta d’Oriente: An oldest genetic legacy for the first modern humans in Sicily. Quat. Sci. Rev. 2020, 248, 106603. [Google Scholar] [CrossRef]
- Sparacello, V.S.; Panelli, C.; Rossi, S.; Dori, I.; Varalli, A.; Goude, G.; Starnini, E.; Biagi, P. The re-discovery of Arma dell’Aquila (Finale Ligure, Italy): New insights on Neolithic funerary behavior from the sixth millennium BCE in the north-western Mediterranean. Quat. Int. 2019, 512, 67–81. [Google Scholar] [CrossRef]
- Fernandes, D.M.; Mittnik, A.; Olalde, I.; Lazaridis, I.; Cheronet, O.; Rohland, N.; Mallick, S.; Bernardos, R.; Broomandkhoshbacht, N.; Carlsson, J.; et al. The spread of steppe and Iranian-related ancestry in the islands of the western Mediterranean. Nat. Ecol. Evol. 2020, 4, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Amadasi Guzzo, M.G. Phoenician and Punic in Sicily. In Language and Linguistic Contact in Ancient Sicily; Tribulato, O., Ed.; Cambridge University Press: New York, NY, USA, 2012; pp. 115–131. [Google Scholar] [CrossRef]
- Shepherd, G. Greek “Colonisation” in Sicily and the West. Some Problems of Evidence and Interpretation Twety-Five Years On. Pallas 2009, 79, 15–25. [Google Scholar] [CrossRef]
- Bradley, K.R. Slave kingdoms and slave rebellions in ancient Sicily. Hist. Refl. 1984, 10, 435–451. [Google Scholar]
- Korhonen, K. Sicily in the Roman Imperial Period from Part III. In Language and Linguistic Contact in Ancient Sicily; Tribulato, O., Ed.; Cambridge University Press: New York, NY, USA, 2012; pp. 326–369. [Google Scholar] [CrossRef]
- Chowaniec, R. Vandals, Ostrogoths and the Byzantine footprints in Sicily: An archaeological-historical review. Institute of Archaeology, University of Warsaw. Med. Archaeol. Archaeom. 2019, 19, 51–61. [Google Scholar] [CrossRef]
- Mandalà, G. The Sicilian Questions. J. Transc. Med. Stud. 2015, 3, 1–2. [Google Scholar] [CrossRef]
- Lo Bue, L.; Lo Bue, F. Da Al Shāqquiyyin a Al Shāqqah. Origini Della Città di Sciacca e Del Suo Toponimo; Sicilgrafica: Palermo, Italy, 2014; pp. 1–89. [Google Scholar]
- Pierre-Amédée, J. Géographie d’Édrisi; Imprimerie Royale: Paris, France, 1840; p. 86. [Google Scholar]
- Tusa, S. (Ed.) Prima Sicilia: Alle Origini della Società Siciliana; Ediprint: Siracusa, IT, USA, 1997. [Google Scholar]
- Regione Siciliana. Linee Guida del Piano Territoriale Paesistico Regionale; Assessorato regionale Beni Culturali e Ambientali e della Pubblica Istruzione: Palermo, Italy, 1996; pp. 76–91. Available online: https://www2.regione.sicilia.it/beniculturali/dirbenicult/bca/ptpr/02articolazione.pdf (accessed on 1 July 2021).
- Giglioli, I. On not being European enough. Migration, crisis and precarious livelihoods on the periphery of Europe. Soc. Cult. Geogr. 2021, 22, 725–744. [Google Scholar] [CrossRef]
- Renfrew, C. From molecular genetics to archaeogenetics. Proc. Natl. Acad. Sci. USA 2001, 98, 4830–4832. [Google Scholar] [CrossRef]
- Piazza, A.; Cappello, N.; Olivetti, E.; Rendine, S. Genetic history of Italy A. Ann. Hum. Genet. 1988, 52, 203–313. [Google Scholar] [CrossRef]
- Cavalli-Sforza, L.L.; Menozzi, P.; Piazza, A. The History and Geography of Human Genes; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Guglielmino, C.R.; Zei, G.; Cavalli-Sforza, L.L. Genetic and Cultural Transmission in Sicily as Revealed by Names and Surnames. Hum. Biol. 1991, 63, 607–627. [Google Scholar]
- Piazza, A.; Rendine, S.; Zei, G.; Moroni, A.; Cavalli-Sforza, L.L. Migration rates of human populations from surname distributions. Nature 1987, 329, 714–771. [Google Scholar] [CrossRef]
- Rickards, O.; Biondi, G.; De Stefano, G.F.; Vecchi, F.; Walter, H. Genetic structure of the population of Sicily. Am. J. Phys. Anthr. 1992, 87, 395–406. [Google Scholar] [CrossRef]
- Vona, G.; Calì, C.M.; Autuori, L.; Mameli, G.E.; Lixi, M.F.; Ghiani, M.E.; Di Gaetano, C. Genetic structure of western Sicily. Int. J. Anthr. 1998, 13, 137–147. [Google Scholar] [CrossRef]
- Piazza, A. L’eredità genetica dell’Italia antica. Le Sci. 1991, 278, 62–69. [Google Scholar]
- Calì, F.; Dianzani, I.; Desviat, L.; Perez, B.; Ugarte, M.; Ozguc, M.; Seyrantepe, V.; Shiloh, Y.; Giannattasio, S.; Carducci, C.; et al. The STR252—IVS10nt546—VNTR7 phenylalanine hydroxylase minihaplotype in five Mediterranean samples. Hum. Gen. 1997, 100, 350–355. [Google Scholar] [CrossRef]
- Semino, O.; Torroni, A.; Scozzari, R.; Brega, A.; De Benedictis, G.; Santachiara Benerecetti, A.S. Mitochondrial DNA polymorphisms in Italy. III. Population data from Sicily: A possible quantitation of maternal African ancestry. Ann. Hum. Genet. 1989, 53, 193–202. [Google Scholar] [CrossRef]
- Vona, G.; Ghiani, C.M.; Calì, C.M.; Vacca, L.; Memmì, M.; Varesi, L. Mitochondrial DNA sequence analysis in Sicily. Am. J. Hum. Biol. 2001, 13, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Calì, C.M.; Garofano, L.; Mameli, A.; Pizzamiglio, M.; Vona, G. Genetic analysis of a Sicilian population using 15 short tandem repeats. Hum. Biol. 2003, 75, 163–178. [Google Scholar] [CrossRef]
- Forster, P.; Calì, F.; Röhl, A.; Metspalu, E.; D’Anna, R.; Mirisola, M.; De Leo, G.; Flugy, A.; Salerno, A.; Ayala, G.; et al. Continental and subcontinental distributions of mtDNA control region types. Int. J. Leg. Med. 2002, 116, 99–108. [Google Scholar] [CrossRef]
- Romano, V.; Calì, F.; Ragalmuto, A.; D’Anna, R.P.; Flugy, A.; De Leo, G.; Giambalvo, O.; Lisa, A.; Fiorani, O.; Di Gaetano, C.; et al. Autosomal Microsatellite and mtDNA Genetic Analysis in Sicily (Italy). Ann. Hum. Gen. 2003, 67, 42–53. [Google Scholar] [CrossRef]
- Capelli, C.; Redhead, N.; Romano, V.; Calì, F.; Lefranc, G.; Delague, V.; Megarbane, A.; Felice, A.E.; Pascali, V.L.; Neophytou, P.I.; et al. Population Structure in the Mediterranean Basin: A Y Chromosome Perspective. Ann. Hum. Gen. 2006, 70, 207–225. [Google Scholar] [CrossRef]
- Di Gaetano, C.; Cerutti, N.; Crobu, F.; Robino, C.; Inturri, S.; Gino, S.; Guarrera, S.; Underhill, P.A.; King, R.J.; Romano, V.; et al. Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome. Eur. J. Hum. Gen. 2009, 17, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Sarno, S.; Boattini, A.; Carta, M.; Ferri, G.; Alù, M.; Yang Yao, D.; Ciani, G.; Pettener, D.; Luiselli, D. An ancient Mediterranean melting pot: Investigating the uniparental genetic structure and population history of Sicily and Southern Italy. PLoS ONE 2014, 9, e96074. [Google Scholar] [CrossRef] [PubMed]
- Lazaridis, I.; Patterson, N.; Mittnik, A.; Renaud, G.; Mallick, S.; Kirsanow, K.; Sudmant, P.H.; Schraiber, J.G.; Castellano, S.; Lipson, M.; et al. Ancient human genomes suggest three ancestral populations for present-day Europeans. Nature 2014, 513, 409–413. [Google Scholar] [CrossRef]
- Busby, G.B.J.; Hellenthal, G.; Montinaro, F.; Tofanelli, S.; Bulayeva, K.; Rudan, I.; Zemunik, T.; Hayward, C.; Toncheva, D.; Karachanak-Yankova, S.; et al. The Role of Recent Admixture in Forming the Contemporary West Eurasian Genomic Landscape. Curr. Biol. 2015, 25, 2518–2526. [Google Scholar] [CrossRef] [PubMed]
- Tofanelli, S.; Brisighelli, F.; Anagnostou, P.; Busby, G.B.J.; Ferri, G.; Thomas, M.G.; Taglioli, L.; Rudan, I.; Zemunik, T.; Hayward, C.; et al. The Greeks in the West: Genetic signatures of the Hellenic colonisation in southern Italy and Sicily. Eur. J. Hum. Gen. 2016, 24, 429–436. [Google Scholar] [CrossRef]
- Sarno, S.; Boattini, A.; Pagani, L.; Sazzini, M.; De Fanti, S.; Quagliariello, A.; Gnecchi Ruscone, G.A.; Guichard, E.; Ciani, G.; Bortolini, E.; et al. Ancient and recent admixture layers in Sicily and Southern Italy trace multiple migration routes along the Mediterranean. Sci. Rep. 2017, 7, 1984. [Google Scholar] [CrossRef]
- Higuchi, R.; Bowman, B.; Freiberger, M.; Ryder, O.A.; Wilson, A. DNA sequences from the quagga, an extinct member of the horse family. Nature 1984, 312, 282–284. [Google Scholar] [CrossRef]
- Pääbo, S. Molecular cloning of Ancient Egyptian mummy DNA. Nature 1985, 314, 644–645. [Google Scholar] [CrossRef] [PubMed]
- Modi, A.; Vai, S.; Posth, C.; Vergata, C.; Zaro, V.; Diroma, M.A.; Boschin, F.; Capecchi, G.; Ricci, S.; Ronchitelli, A.; et al. More data on ancient human mitogenome variability in Italy: New mitochondrial genome sequences from three Upper Palaeolithic burials. Ann. Hum. Biol. 2021, in press. [Google Scholar]
- van de Loosdrecht, M.S.; Mannino, M.A.; Talamo, S.; Villalba-Mouco, V.; Posth, C.; Aron, F.; Burri, M.; Brandt, G.; Freund, C.; Radzeviciute, R.; et al. Genomic and Dietary Transitions during the Mesolithic and Early Neolithic in Sicily. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mannino, M.A.; Catalano, G.; Talamo, S.; Mannino, G.; Di Salvo, R.; Schimmenti, V.; Lalueza-Fox, C.; Messina, A.; Petruso, D.; Caramelli, D.; et al. Origin and diet of the prehistoric hunter-gatherers on the mediterranean island of Favignana (Egadi islands, Sicily). PLoS ONE 2012, 7, e49802. [Google Scholar] [CrossRef]
- Olalde, I.; Brace, S.; Allentoft, M.E.; Armit, I.; Kristiansen, K.; Booth, T.; Rohland, R.; Mallick, S.; Szécsényi-Nagy, A.; Mittnik, A.; et al. The Beaker phenomenon and the genomic transformation of northwest Europe. Nature 2018, 555, 190–196. [Google Scholar] [CrossRef]
- Marino, P.; Castiglia, G.; Bazan, G.; Domina, G.; Guarino, R. Tertiary relict laurophyll vegetation in the Madonie mountains (Sicily). Acta Bot. Gal. 2014, 161, 47–61. [Google Scholar] [CrossRef][Green Version]
- Garfì, G.; Carimi, F.; Fazan, L.; Gristina, A.S.; Kozlowski, G.; Livreri Console, S.; Antonio Motisi Pasta, S. From glacial refugia to hydrological microrefugia: Factors and processes driving the persistence of the climate relict tree. Zelkova sicula. Ecol. Evol. 2021, 11, 2919–2936. [Google Scholar] [CrossRef]
- Magri, D.; Agrillo, E.; Di Rita, F.; Furlanetto, G.; Pini, R.; Ravazzi, C.; Spada, F. Holocene dynamics of tree taxa populations in Italy. Rev. Palaeob. Palyn. 2015, 218, 267–284. [Google Scholar] [CrossRef]
- Incarbona, A.; Zarcone, G.; Agate, M.; Bonomo, S.; Stefano, E.; Masini, F.; Russo, F.; Sineo, L. A multidisciplinary approach to reveal the Sicily Climate and Environment over the last 20000 years. Cent. Eur. J. Geosci. 2010, 2, 71–82. [Google Scholar] [CrossRef]
- Sadori, L.; Narcisi, B. The Postglacial record of environmental history from Lago di Pergusa, Sicily. Holocene 2001, 11, 655–671. [Google Scholar] [CrossRef]
- Sadori, L.; Giardini, M. Charcoal analysis, a method to study vegetation and climate of the Holocene: The case of Lago di Pergusa, Sicily (Italy). Geobios 2007, 40, 173–180. [Google Scholar] [CrossRef]
- Sadori, L.; Zanchetta, G.; Giardini, M. Last Glacial to Holocene palaeoenvironmental evolution at Lago di Pergusa (Sicily, Southern Italy) as inferred by pollen, microcharcoal, and stable isotopes. Quat. Int. 2008, 181, 4–14. [Google Scholar] [CrossRef]
- Sadori, L.; Jahns, S.; Peyron, O. Mid-Holocene vegetation history of the central Mediterranean. Holocene 2011, 21, 117–129. [Google Scholar] [CrossRef]
- Sadori, L.; Ortu, E.; Peyron, O.; Zanchetta, G.; Vanniere, B.; Desmet, M.; Magny, M. The last 7 millennia of vegetation and climate changes at Lago di Pergusa (central Sicily, Italy). Clim. Past. 2013, 9, 1969–1984. [Google Scholar] [CrossRef]
- Sadori, L.; Masi, A.; Ricotta, C. Climate driven past fires in central Sicily. Plant. Biosyst. 2015, 149, 166–173. [Google Scholar] [CrossRef]
- Zanchetta, G.; Borghini, A.; Fallick, A.E.; Bonadonna, F.P.; Leone, G. Late Quaternary palaeohydrology of Lake Pergusa (Sicily, southern Italy) as inferred by stable isotopes of lacustrine carbonates. J. Paleolimnol. 2006, 38, 227–239. [Google Scholar] [CrossRef]
- Noti, R.; van Leeuwen, J.F.N.; Colombaroli, D.; Vescovi, E.; Pasta, S.; La Mantia, T.; Tinner, W. Mid- and late- Holocene vegetation and fire history at Biviere di Gela, a coastal lake in southern Sicily, Italy. Veget. Hist. Archaeobot. 2009, 18, 371–387. [Google Scholar] [CrossRef]
- Tinner, W.; van Leeuwen, J.F.N.; Colombaroli, D.; Vescovi, E.; van der Knaap, W.O.; Henne, P.D.; Pasta, S.; D’Angelo, S.; La Mantia, T. Holocene environmental and climatic changes at Gorgo Basso, a coastal lake in southern Sicily, Italy. Quat. Sci. Rev. 2009, 28, 1498–1510. [Google Scholar] [CrossRef]
- Calò, C.; Henne, P.D.; Curry, B.; Magny, M.; Vescovi, E.; La Mantia, T.; Pasta, S.; Vanniere, B.; Tinner, W. Spatio-temporal patterns of Holocene environmental change in southern Sicily. Paleogeogr. Paleoclimatol. Paleoecol. 2012, 323, 110–122. [Google Scholar] [CrossRef]
- Bertolani Marchetti, D.; Accorsi, C.A.; Arobba, D.; Bandini Mazzanti, M.; Bertolani, M.; Biondi, E.; Braggio, G.; Ciuffi, C.; De Cunzo, T.; Della Ragione, S.; et al. Recherches geòobotaniques sur les Monts Madonie (Sicile du Nord). Webbia 1984, 38, 329–348. [Google Scholar] [CrossRef]
- Bisculm, M.; Colombaroli, D.; Vescovi, E.; van Leeuwen, J.F.; Henne, P.D.; Rothen, J.; Procacci, G.; Pasta, S.; La Mantia, T.; Tinner, W. Holocene vegetation and fire dynamics in the supra-mediterranean belt of the Nebrodi Mountains (Sicily, Italy). J. Quat. Sci. 2012, 27, 687–698. [Google Scholar] [CrossRef]
- Tinner, W.; Vescovi, E.; van Leeuwen, J.F.; Colombaroli, D.; Henne, P.D.; Kaltenrieder, P.; Morales-Molino, C.; Beffa, G.; Gnaegi, B.; van der Knaap, W.O.; et al. Holocene vegetation and fire history of the mountains of Northern Sicily (Italy). Veg. Hist. Arch. 2016, 25, 499–519. [Google Scholar] [CrossRef]
- Combourieu Nebout, N.; Peyron, O.; Dormoy, I.; Desprat, S.; Beaudouin, C.; Kotthoff, U.; Marret, F. Rapid climatic variability in the west Mediterranean during the last 25,000 years from high resolution pollen data. Clim. Past 2009, 5, 503–521. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Sadori, L. Mediterranean culture and climatic change: Past patterns and future trends. In The Mediterranean Sea—Its History and Present Challenges; Goffredo, S., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 507–527. [Google Scholar]
- Alley, R.B.; Mayewski, P.A.; Sowers, T.; Stuiver, M.; Taylor, K.C.; Clark, P.U. Holocene climatic instability: A prominent, widespread event 8200 years ago. Geology 1997, 25, 483–486. [Google Scholar] [CrossRef]
- Zanchetta, G.; Regattieri, E.; Isola, I.; Drysdale, R.N.; Bini, M.; Baneschi, I.; Hellstrom, J.C. The so-called “4.2 event” in the central Mediterranean and its climatic teleconnections. Alp. Med. Quat. 2016, 29, 5–17. [Google Scholar]
- Gianguzzi, L.; Bazan, G. The Olea europaea L. var. sylvestris (Mill.) Lehr. forests in the Mediterranean area. Plant. Soc. 2019, 56, 3–34. [Google Scholar] [CrossRef]
- Gianguzzi, L.; Bazan, G. A phytosociological analysis of the Olea europaea L. var. sylvestris (Mill.) Lehr. forests in Sicily. Plant. Biosyst. 2020, 154, 705–725. [Google Scholar] [CrossRef]
- Bazan, G.; Marino, P.; Guarino, R.; Domina, G.; Schicchi, R. Bioclimatology and vegetation series in Sicily: A geostatistical approach. Ann. Bot. Fenn. 2015, 52, 1–18. [Google Scholar] [CrossRef]
- Crispino, A. Castelluccio (Noto, SR), Notiziario di Preistoria e Protostoria—II. Sard. Sicil. 2018, 5, 98–102. [Google Scholar]
- Leighton, R. Sicily before History: An Archaeological Survey from the Palaeolithic to the Iron Age; Cornell University Press: Ithaca, NY, USA, 1999. [Google Scholar]
- Tagliacozzo, A. Archeozoologia Della Grotta dell’Uzzo, Sicilia: Da Un’Economia di Caccia ad Un’Economia di Pesca ed Allevamento; Istituto poligrafico e zecca dello Stato: Roma, Italy, 1993. [Google Scholar]
- Marcott, S.A.; Shakun, J.D.; Clark, P.U.; Mix, A.C. A reconstruction of regional and global temperature for the past 11,300 years. Science 2013, 339, 1198–1201. [Google Scholar] [CrossRef]
- Kaufman, D.; McKay, N.; Routson, C.; Erb, M.; Dätwyler, C.; Sommer, P.S.; Heiri, O.; Davis, B. Holocene global mean surface temperature, a multi-method reconstruction approach. Sci. Data 2020, 7, 1–13. [Google Scholar] [CrossRef]
- Giannitrapani, E.; Pluciennik, M. La seconda campagna di ricognizione del progetto “Archeologia nella valle del Torcicoda”. Sicil. Archeol. 1997, 96, 59–69. [Google Scholar]
- Sadori, L.; Giraudi, C.; Masi, A.; Magny, M.; Ortu, E.; Zanchetta, G.; Izdebski, A. Climate, environment and society in southern Italy during the last 2000 years. A review of the environmental, historical and archaeological evidence. Quat. Sci. Rev. 2016, 136, 173–188. [Google Scholar] [CrossRef]
- Todaro, P.; Barbera, G.; Castrorao Barba, A.; Bazan, G. Qanāts and historical irrigated landscapes in Palermo’s suburban area (Sicily). Eur. J. Post Archaeol. 2020, 10, 335–370. [Google Scholar]
- Castrorao Barba, A.; Speciale, C.; Miccichè, R.; Pisciotta, F.; Aleo Nero, C.; Marino, P.; Bazan, G. The Sicilian Countryside in the Early Middle Ages: Human-Environment Interactions at Contrada Castro. Environ. Arch. 2021. [Google Scholar] [CrossRef]
- Bazan, G.; Castrorao Barba, A.; Rotolo, A.; Marino, P. Geobotanical approach to detect land-use change of a Mediterranean landscape: A case study in Central-Western Sicily. Geo J. 2019, 84, 795–811. [Google Scholar] [CrossRef]
- Bazan, G.; Castrorao Barba, A.; Rotolo, A.; Marino, P. Vegetation series as a marker of interactions between rural settlements and landscape: New insights from the archaeological record in Western Sicily. Landsc. Res. 2020, 45, 1–19. [Google Scholar] [CrossRef]
- Tusa, S. Selinunte; L’Erma di Bretschneider: Roma, Italy, 2010; pp. 7–231. [Google Scholar]
- Baiamonte, G.; Domina, G.; Raimondo, F.M.; Bazan, G. Agricultural landscapes and biodiversity conservation: A case study in Sicily (Italy). Biod. Cons. 2015, 24, 3201–3216. [Google Scholar] [CrossRef]
- Troia, A.; Bazan, G.; Schicchi, R. Micromorphological approach to the systematics of Mediterranean Isoëtes species (Isoëtaceae, Lycopodiophyta): Analysis of the megaspore surface. Grana 2012, 51, 35–43. [Google Scholar] [CrossRef]
- Marino, P.; Guarino, R.; Bazan, G. The Sicilian taxa of Genista sect. Voglera and their phytosociological framework. Flora Mediterr. 2012, 22, 169–190. [Google Scholar] [CrossRef]
- Tanasi, D.; Greco, E.; Noor, R.E.; Feola, S.; Kumar, V.; Crispino, A.; Gelis, I. 1 H NMR, 1 H-1 H 2D TOCSY and GC-MS analyses for the identification of olive oil in Early Bronze Age pottery from Castelluccio (Noto, Italy). Anal. Meth. 2018, 10, 2756–2763. [Google Scholar] [CrossRef]
- Tanasi, D.; Greco, E.; Di Tullio, V.; Capitani, D.; Gullì, D.; Ciliberto, E. 1H-1H NMR 2D-TOCSY, ATR FT-IR and SEM-EDX for the identification of organic residues on Sicilian prehistoric pottery. Microch. J. 2017, 135, 140–147. [Google Scholar] [CrossRef]
- Bazan, G.; Civantos, J.M. Agrobiodiversity as Mediterranean Agrarian Heritage. Memola Project European Policy Brief. 2017. Available online: https://www.open-heritage.eu/wp-content/uploads/2019/09/d9.5_3rd_policy_brief.pdf (accessed on 1 July 2021).
- Medail, F.; Quezel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, V.; Catalano, G.; Bazan, G.; Calì, F.; Sineo, L. Archaeogenetics and Landscape Dynamics in Sicily during the Holocene: A Review. Sustainability 2021, 13, 9469. https://doi.org/10.3390/su13179469
Romano V, Catalano G, Bazan G, Calì F, Sineo L. Archaeogenetics and Landscape Dynamics in Sicily during the Holocene: A Review. Sustainability. 2021; 13(17):9469. https://doi.org/10.3390/su13179469
Chicago/Turabian StyleRomano, Valentino, Giulio Catalano, Giuseppe Bazan, Francesco Calì, and Luca Sineo. 2021. "Archaeogenetics and Landscape Dynamics in Sicily during the Holocene: A Review" Sustainability 13, no. 17: 9469. https://doi.org/10.3390/su13179469
APA StyleRomano, V., Catalano, G., Bazan, G., Calì, F., & Sineo, L. (2021). Archaeogenetics and Landscape Dynamics in Sicily during the Holocene: A Review. Sustainability, 13(17), 9469. https://doi.org/10.3390/su13179469