Biennial Mowing Maintains the Biomass and Functional Diversity of Semi-Arid Grassland

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Experiment Design
2.3. Sample Collection
2.4. Statistical Analysis
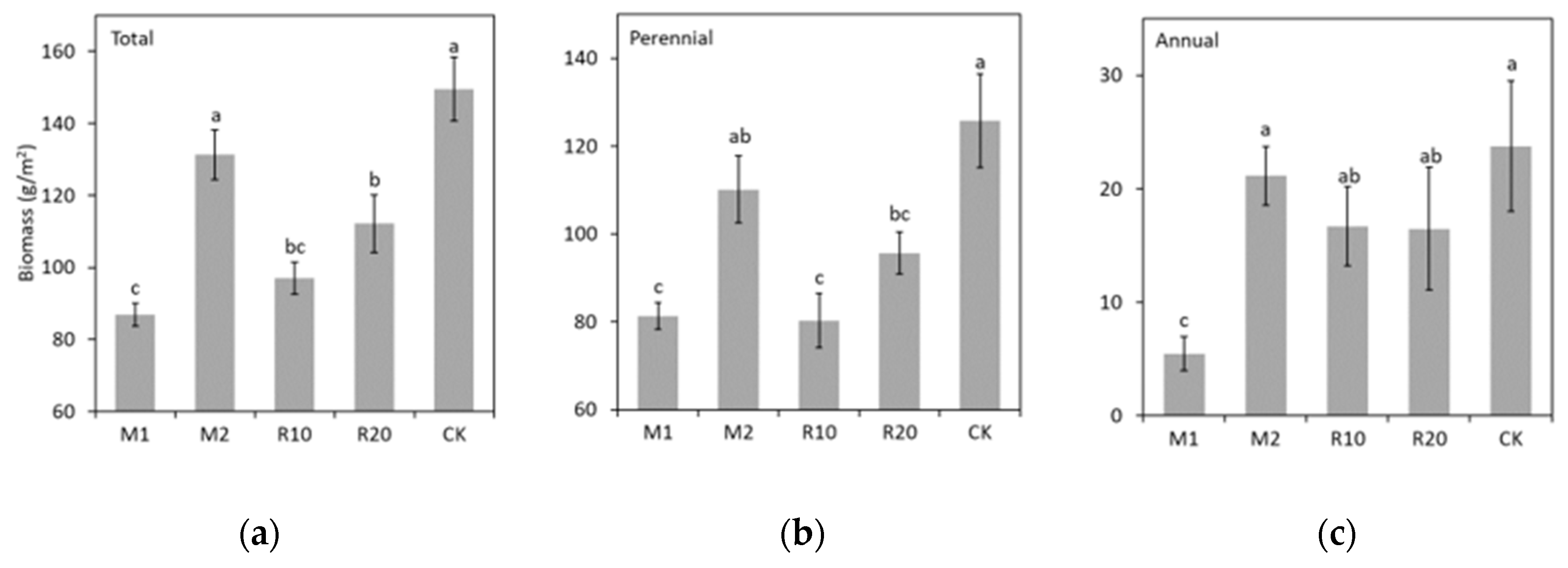
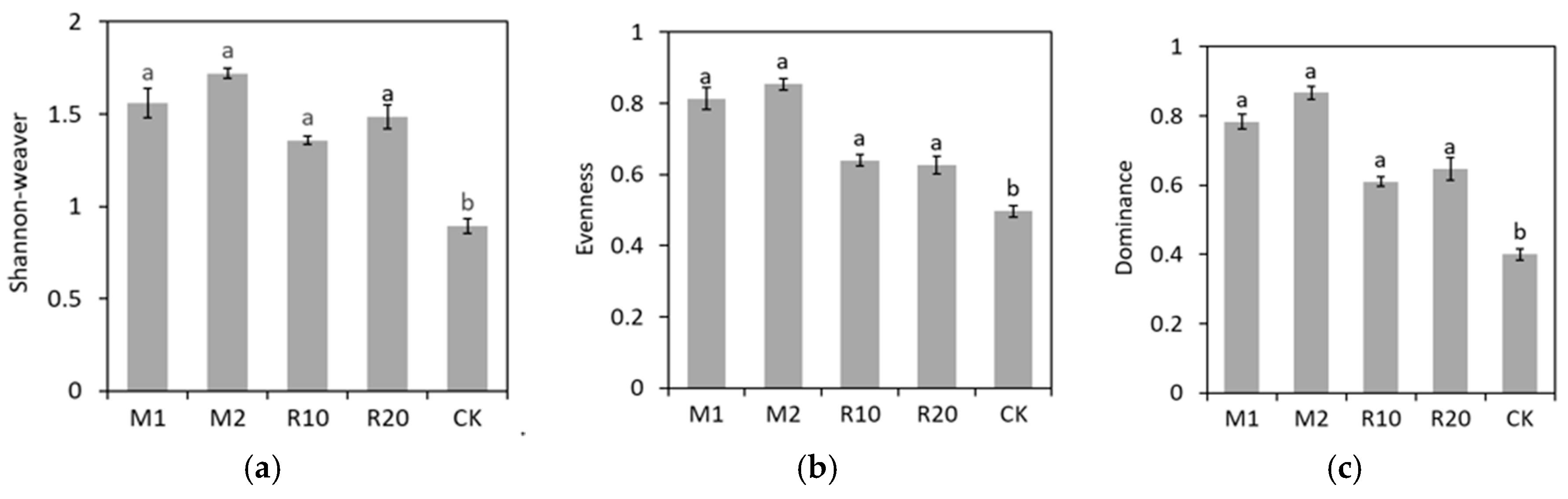
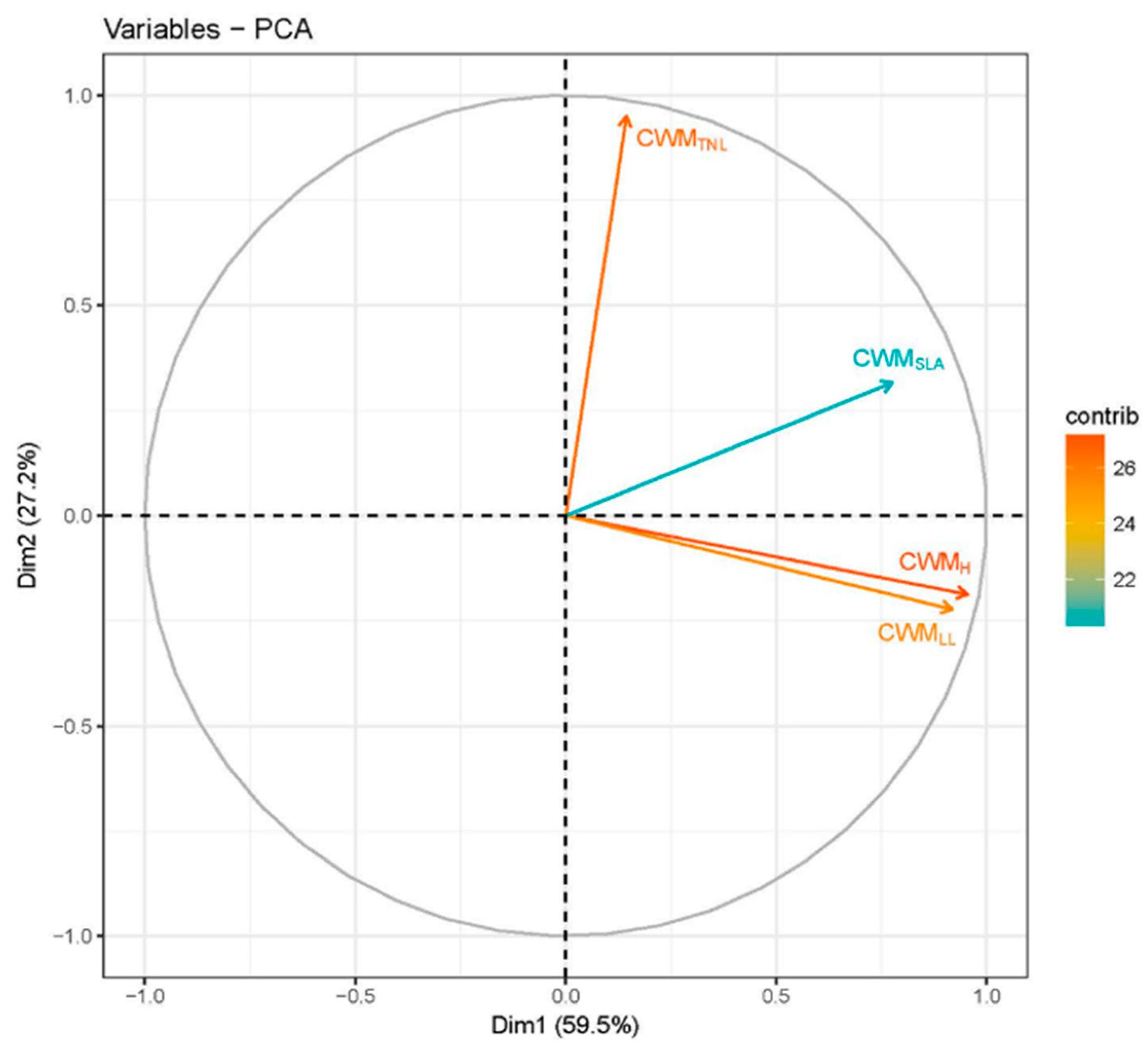
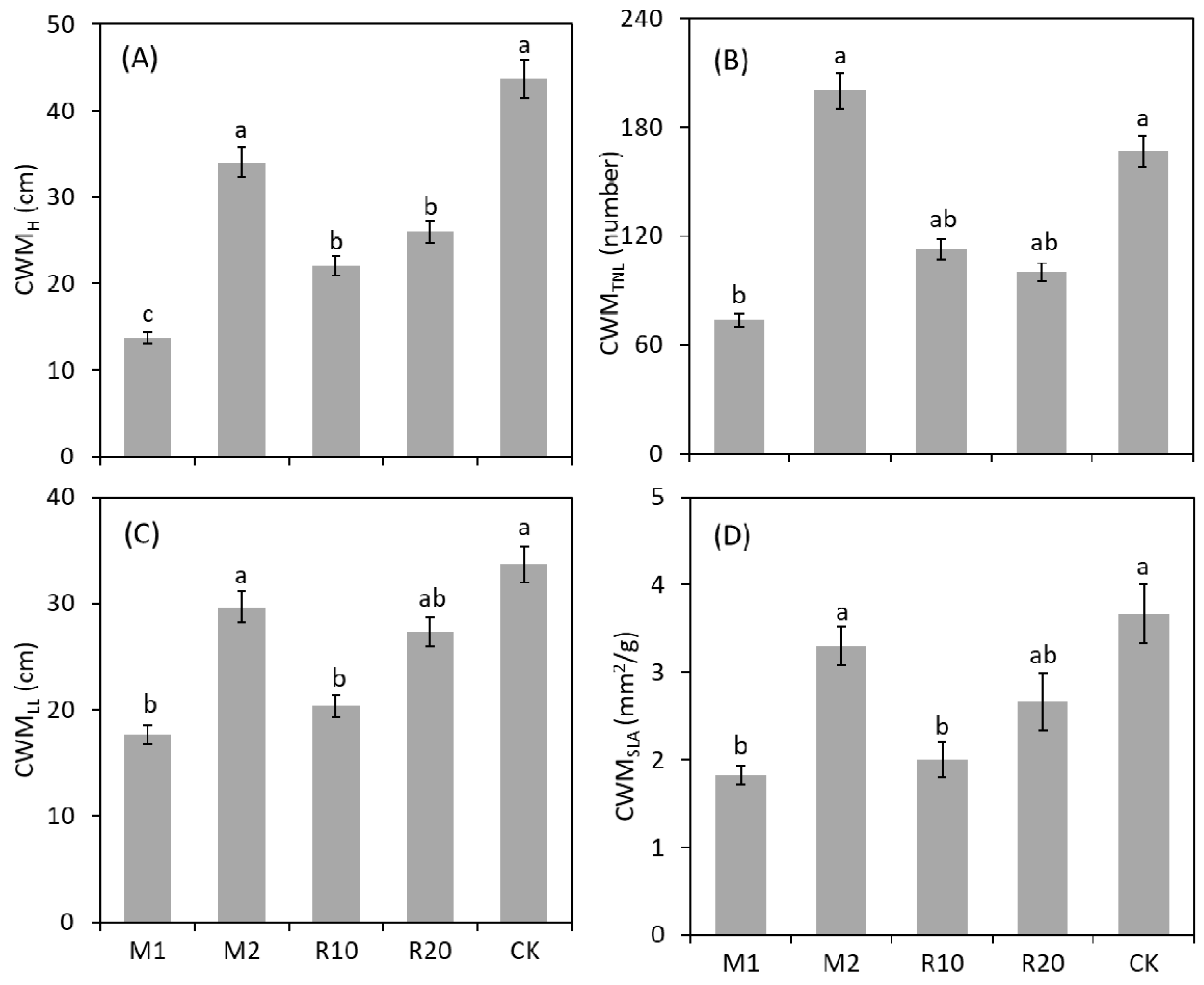
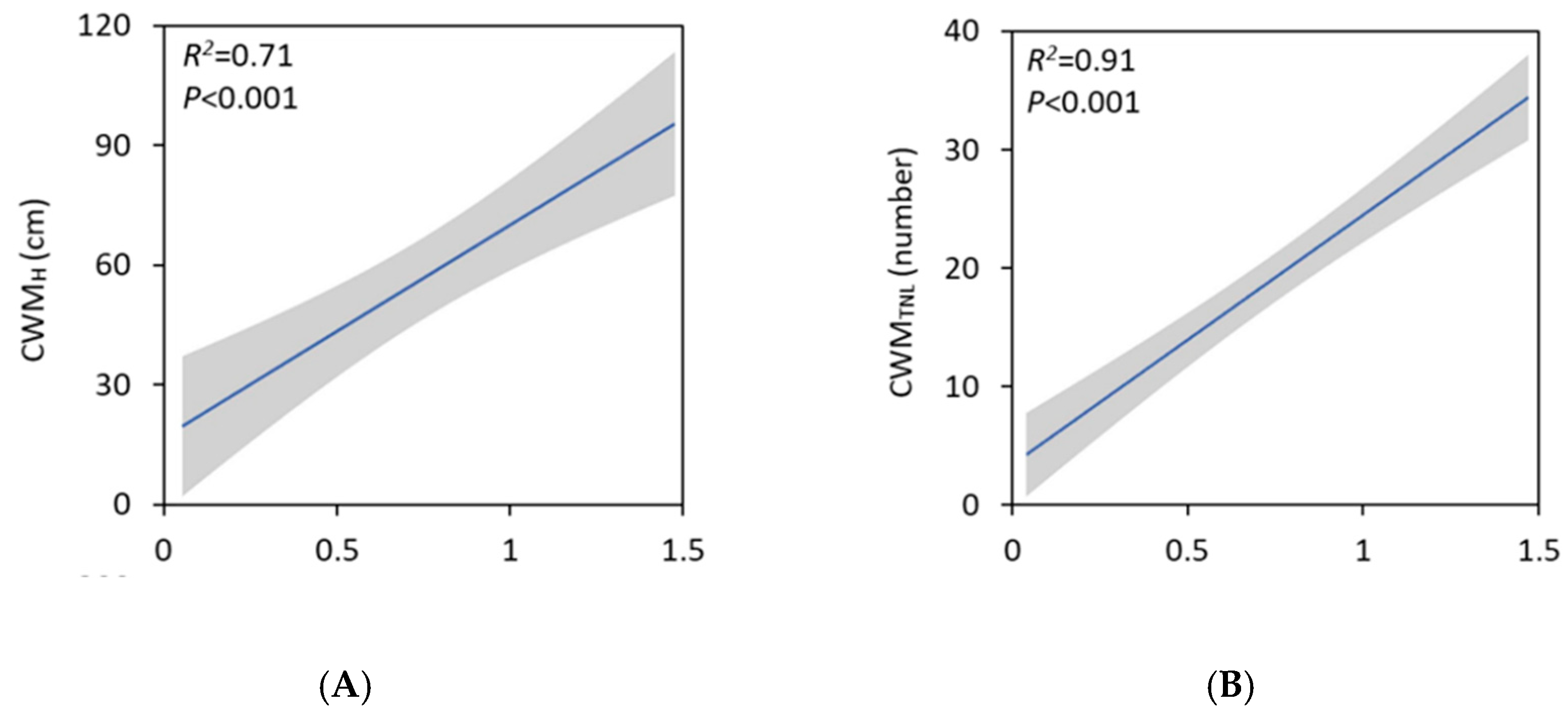
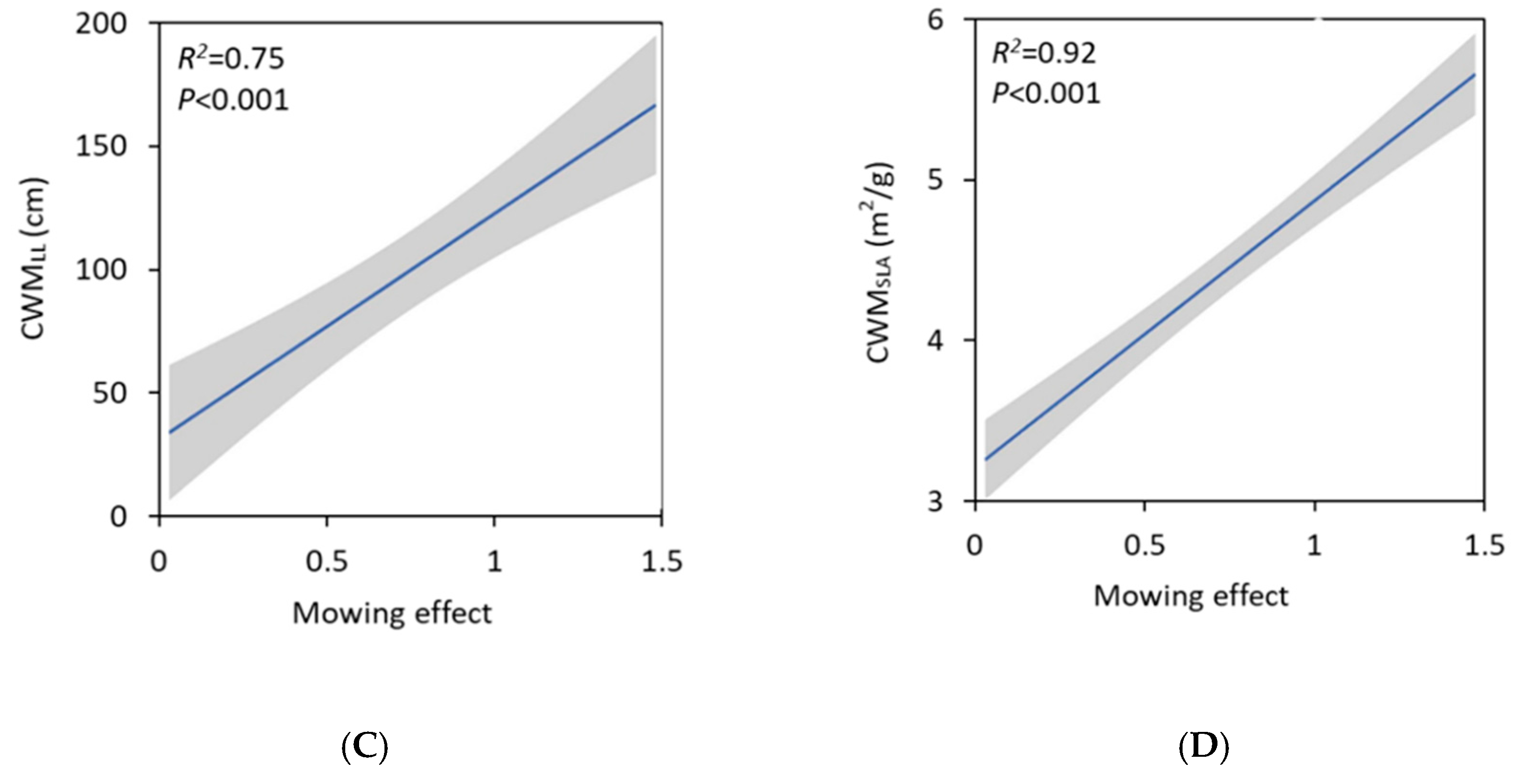
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Deléglise, C.; Meisser, M.; Mosimann, E.; Spiegelberger, T.; Signarbieux, C.; Jeangros, B. Drought-induced shifts in plants traits, yields and nutritive value under realistic grazing and mowing managements in a mountain grassland. Agr. Ecosyst. Environ. 2015, 213, 94–104. [Google Scholar] [CrossRef]
- Bai, Y.; Wu, J.; Clark, C.M.; Pan, Q.; Zhang, L.; Chen, S. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J. Appl. Ecol. 2012, 49, 1204–1215. [Google Scholar] [CrossRef]
- Li, X.; Wu, Z.; Liu, Z.; Hou, X.; Badgery, W.; Guo, H. Contrasting Effects of Long-Term Grazing and Clipping on Plant Morphological Plasticity: Evidence from a Rhizomatous Grass. PLoS ONE 2015, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Buri, P.; Arlettaz, R.; Humbert, J.Y. Delaying mowing and leaving uncut refuges boosts orthopterans in extensively managed meadows: Evidence drawn from field-scale experimentation. Agr. Ecosyst. Environ. 2013, 181, 22–30. [Google Scholar] [CrossRef]
- Grime, J. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Wen, Z.; Zheng, H.; Smith, J.R.; Zhao, H.; Liu, L.; Ouyang, Z. Functional diversity overrides community-weighted mean traits in linking land-use intensity to hydrological ecosystem services. Sci. Total Environ. 2019, 682, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Mauchamp, L.; Mouly, A.; Badot, P.M.; Gillet, F. Impact of management type and intensity on multiple facets of grassland biodiversity in the F rench J ura M ountains. Appl. Veg. Sci. 2014, 17, 645–657. [Google Scholar] [CrossRef]
- Han, G.; Hao, X.; Zhao, M.; Wang, M.; Ellert, B.H.; Willms, W. Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agr. Ecosyst. Environ. 2008, 125, 21–32. [Google Scholar] [CrossRef]
- Qu, L.; Chen, J.; Dong, G.; Shao, C. Heavy mowing enhances the effects of heat waves on grassland carbon and water fluxes. Sci. Total Environ. 2018, 627, 561–570. [Google Scholar] [CrossRef]
- Wellstein, C.; Schröder, B.; Reineking, B.; Zimmermann, N.E. Understanding species and community response to environmental change-A functional trait perspective. Agr. Ecosyst. Environ. 2011, 145, 1–4. [Google Scholar] [CrossRef]
- Kleijn, D.; Sutherland, W.J. How effective are European agri-environment schemes in conserving and promoting biodiversity? J. Appl. Ecol. 2003, 40, 947–969. [Google Scholar] [CrossRef]
- Baoyin, T.; Li, F.Y.; Bao, Q.; Minggagud, H.; Zhong, Y. Effects of mowing regimes and climate variability on hay production of Leymus chinensis Trin. Tzvelev grassland in northern China. Rangeland J. 2014, 36, 593–600. [Google Scholar] [CrossRef]
- Wu, N.; Liu, A.; Wang, Y.; Li, L.; Chao, L.; Liu, G. An Assessment Framework for Grassland Ecosystem Health with Consideration of Natural Succession: A Case Study in Bayinxile, China. Sustainability 2019, 11, 1096. [Google Scholar] [CrossRef]
- Piqueray, J.; Gilliaux, V.; Decruyenaere, V.; Cornelis, J.T.; Uyttenbroeck, R.; Mahy, G. Management of Grassland-like Wildflower Strips Sown on Nutrient-rich Arable Soils: The Role of Grass Density and Mowing Regime. Environ. Manag. 2019, 63, 647–657. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Römermann, C.; Sperlich, S.; Schmidt, W. Explaining grassland biomass-the contribution of climate.; species and functional diversity depends on fertilization and mowing frequency. J. Appl. Ecol. 2011, 48, 1088–1097. [Google Scholar] [CrossRef]
- Polley, H.W.; Isbell, F.I.; Wilsey, B.J. Plant functional traits improve diversity-based predictions of temporal stability of grassland productivity. Oikos 2013, 122, 1275–1282. [Google Scholar] [CrossRef]
- Saruul, K.; Jiangwen, L.; Jianming, N.; Qing, Z.; Xuefeng, Z.; Guodong, H.; Haifeng, B. Typical steppe ecosystems maintain high stability by decreasing the connections among recovery, resistance, and variability under high grazing pressure. Sci. Total Environ. 2019, 659, 1146–1157. [Google Scholar] [CrossRef]
- Yang, H.; Jiang, L.; Li, L.; Li, A.; Wu, M.; Wan, S. Diversity-dependent stability under mowing and nutrient addition: Evidence from a 7-year grassland experiment. Ecol. Lett. 2012, 15, 619–626. [Google Scholar] [CrossRef]
- Leegood, R.C. Strategies for engineering C4 photosynthesis. J. Plant. Physiol. 2013, 170, 378–388. [Google Scholar] [CrossRef]
- Brett Mattingly, W.; Hewlate, R.; Reynolds, H.L. Species evenness and invasion resistance of experimental grassland communities. Oikos 2007, 116, 1164–1170. [Google Scholar] [CrossRef]
- Chanteloup, P.; Bonis, A. Functional diversity in root and above-ground traits in a fertile grassland shows a detrimental effect on productivity. Basic Appl. Ecol. 2013, 14, 208–216. [Google Scholar] [CrossRef]
- Van Klink, R.; Boch, S.; Buri, P.; Rieder, N.S.; Humbert, J.Y.; Arlettaz, R. No detrimental effects of delayed mowing or uncut grass refuges on plant and bryophyte community structure and phytomass production in low-intensity hay meadows. Basic Appl. Ecol. 2017, 20, 1–9. [Google Scholar] [CrossRef]
- Doležal, J.; Lanta, V.; Mudrák, O.; Lepš, J.; Wilson, S. Seasonality promotes grassland diversity: Interactions with mowing, fertilization and removal of dominant species. J. Ecol. 2018, 107, 203–215. [Google Scholar] [CrossRef]
- Denelle, P.; Violle, C.; Munoz, F. Distinguishing the signatures of local environmental filtering and regional trait range limits in the study of trait-environment relationships. Oikos 2019, 128, 960–971. [Google Scholar] [CrossRef]
- Finegan, B.; Peña-Claros, M.; de Oliveira, A.; Ascarrunz, N.; Bret-Harte, M.S.; Carreño-Rocabado, G. Does functional trait diversity predict above-ground biomass and productivity of tropical forests? Testing three alternative hypotheses. J. Ecol. 2015, 103, 191–201. [Google Scholar] [CrossRef]
- Ali, A.; Yan, E.R.; Chang, S.X.; Cheng, J.Y.; Liu, X.Y. Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests. Sci. Total Environ. 2017, 574, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Buzzard, V.; Hulshof, C.M.; Birt, T.; Violle, C.; Enquist, B.J.; Larjavaara, M. Re-growing a tropical dry forest: Functional plant trait composition and community assembly during succession. Funct. Ecol. 2016, 30, 1006–1013. [Google Scholar] [CrossRef]
- Zhang, Q.; Buyantuev, A.; Li, F.Y.; Jiang, L.; Niu, J.; Ding, Y. Functional dominance rather than taxonomic diversity and functional diversity mainly affects community aboveground biomass in the Inner Mongolia grassland. Ecol. Evol. 2017, 7, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, M.; Qi, Y.; Dong, Y.; He, X.; Li, J.; Sun, L. Differential responses of soil microbial community to four-decade long grazing and cultivation in a semi-arid grassland. Sustainability 2017, 9, 128. [Google Scholar] [CrossRef]
- Liang, E.; Vennetier, M.; Lin, J.; Shao, X. Relationships between tree increment, climate and above-ground biomass of grass: A case study in the typical steppe, north China. Acta Oecologica 2003, 24, 87–94. [Google Scholar] [CrossRef]
- Wu, Z. Vegetation of China; Science Press: Beijing, China, 1980. (In Chinese) [Google Scholar]
- Jiang, S. Setting up the experimental sites for grassland ecosystem research and vegetation status. Res. Grassland Ecosyst. 1988, 3, 1–12. [Google Scholar]
- Olmo, M.; Lopez-Iglesias, B.; Villar, R. Drought changes the structure and elemental composition of very fine roots in seedlings of ten woody tree species. Implications for a drier climate. Plant Soil 2014, 384, 113–129. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef]
- Volf, M.; Redmond, C.; Albert, A.J.; Le Bagousse-Pinguet, Y.; Biella, P.; Gotzenberger, L. Effects of long and short term management on the functional structure of meadows through species turnover and intraspecific trait variability. Oecologia 2016, 180, 941–950. [Google Scholar] [CrossRef]
- Bennett, L.T.; Judd, T.S.; Adams, M.A. Growth and nutrient content of perennial grasslands following burning in semi-arid, sub-tropical Australia. Plant. Ecol. 2003, 164, 185–199. [Google Scholar] [CrossRef]
- Gaberščik, A.; Krek, J.L.; Zelnik, I. Habitat diversity along a hydrological gradient in a complex wetland results in high plant species diversity. Ecol. Eng. 2018, 118, 84–92. [Google Scholar] [CrossRef]
- Adler, P.B.; Seabloom, E.W.; Borer, E.T.; Hillebrand, H.; Hautier, Y.; Hector, A.; Harpole, W.S.; O’Halloran, L.R.; Grace, J.B.; Anderson, T.M.; et al. Productivity is a poor predictor of plant species richness. Science 2011, 333, 1750–1753. [Google Scholar] [CrossRef]
- Kemp, D.R.; Guodong, H.; Xiangyang, H.; Michalk, D.L.; Fujiang, H.; Jianping, W.; Yingjun, Z. Innovative grassland management systems for environmental and livelihood benefits. Proc. Natl. Acad. Sci. USA 2013, 110, 8369–8374. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Latin Name | Plants Type | M1 | M2 | R10 | R20 | CK |
|---|---|---|---|---|---|---|---|
| 1 | Stipa grandis | Perennial | + | + | + | + | + |
| 2 | Cleistogenes squarrosa | + | + | + | + | + | |
| 3 | Agropyron michnoi | Absent | + | absent | + | + | |
| 4 | Leymus chinensis | + | + | + | + | + | |
| 5 | Anemarrhena asphodeloides | + | + | + | + | + | |
| 6 | Allium tenuissimum | Absent | + | + | absent | + | |
| 7 | Thalictrum petaloideum | Absent | + | + | + | + | |
| 8 | Allium bidentatum | + | + | + | + | + | |
| 9 | Allium ramosum | Absent | absent | absent | + | absent | |
| 10 | Astragalus galactites | Absent | + | absent | + | absent | |
| 11 | Convolvulus ammannii | + | absent | + | absent | absent | |
| 12 | Scutellaria viscidula | Absent | + | absent | absent | absent | |
| 13 | Carex korshinskyi | + | + | absent | absent | absent | |
| 14 | Caragana microphylla | Absent | + | absent | + | + | |
| 15 | Androsace umbellata | + | absent | absent | + | absent | |
| 16 | Parthenocissus tricuspidata | + | + | + | absent | absent | |
| 17 | Gueldenstaedtia | + | absent | + | absent | absent | |
| 18 | Ephedra sinica | Absent | absent | + | absent | absent | |
| 19 | Artemisia frigida | + | absent | absent | absent | absent | |
| 20 | Cymbaria dahurica | Absent | + | absent | + | + | |
| 21 | Gentiana scabra | Absent | + | absent | absent | absent | |
| 22 | Polygala tenuifolia | + | absent | absent | absent | + | |
| 23 | Oxytropis myriophylla | + | absent | absent | + | absent | |
| 24 | Ruta graveolens | + | absent | + | absent | absent | |
| 25 | Euphorbia esula | Absent | + | absent | absent | + | |
| 26 | Asparagus cochinchinensis | Absent | + | absent | absent | absent | |
| 1 | Corispermum hyssopifolium | Annual | + | + | + | + | + |
| 2 | Chenopodium aristatum | + | + | + | + | + | |
| 3 | Salsola collina | + | + | + | + | + | |
| 4 | Oxybasis glauca | + | + | + | + | absent | |
| 5 | Chenopodium acuminatum | + | + | + | + | absent |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, T.; Zhang, F.; Suo, R.; Gu, C.; Chen, D.; Yang, T.; Zhao, M. Biennial Mowing Maintains the Biomass and Functional Diversity of Semi-Arid Grassland. Sustainability 2020, 12, 1507. https://doi.org/10.3390/su12041507
Zhao T, Zhang F, Suo R, Gu C, Chen D, Yang T, Zhao M. Biennial Mowing Maintains the Biomass and Functional Diversity of Semi-Arid Grassland. Sustainability. 2020; 12(4):1507. https://doi.org/10.3390/su12041507
Chicago/Turabian StyleZhao, Tianqi, Feng Zhang, Rongzhen Suo, Chen Gu, Daling Chen, Tony Yang, and Mengli Zhao. 2020. "Biennial Mowing Maintains the Biomass and Functional Diversity of Semi-Arid Grassland" Sustainability 12, no. 4: 1507. https://doi.org/10.3390/su12041507
APA StyleZhao, T., Zhang, F., Suo, R., Gu, C., Chen, D., Yang, T., & Zhao, M. (2020). Biennial Mowing Maintains the Biomass and Functional Diversity of Semi-Arid Grassland. Sustainability, 12(4), 1507. https://doi.org/10.3390/su12041507