The Global Potential Distribution of Invasive Plants: Anredera cordifolia under Climate Change and Human Activity Based on Random Forest Models


Abstract
1. Introduction
2. Materials and Methods
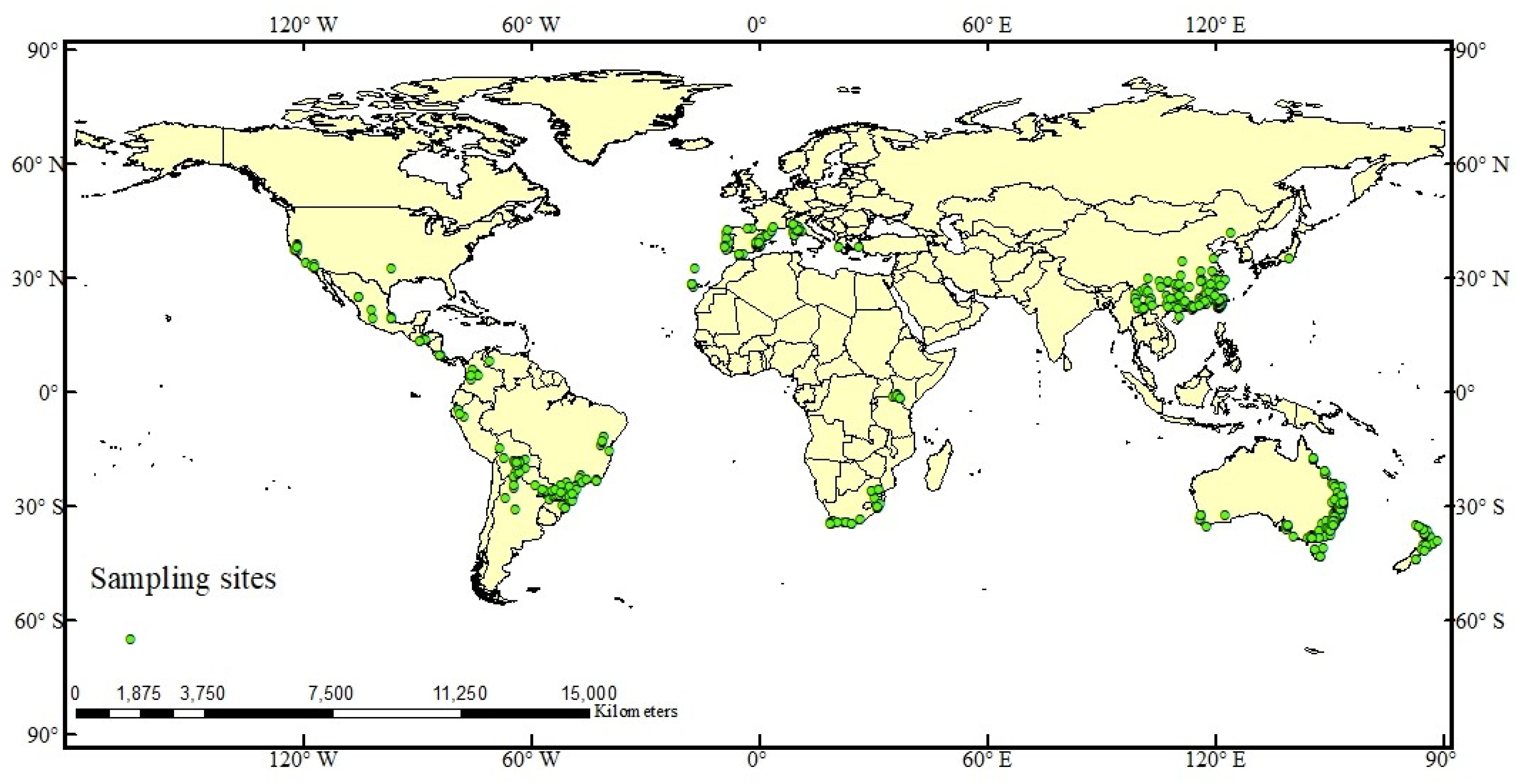
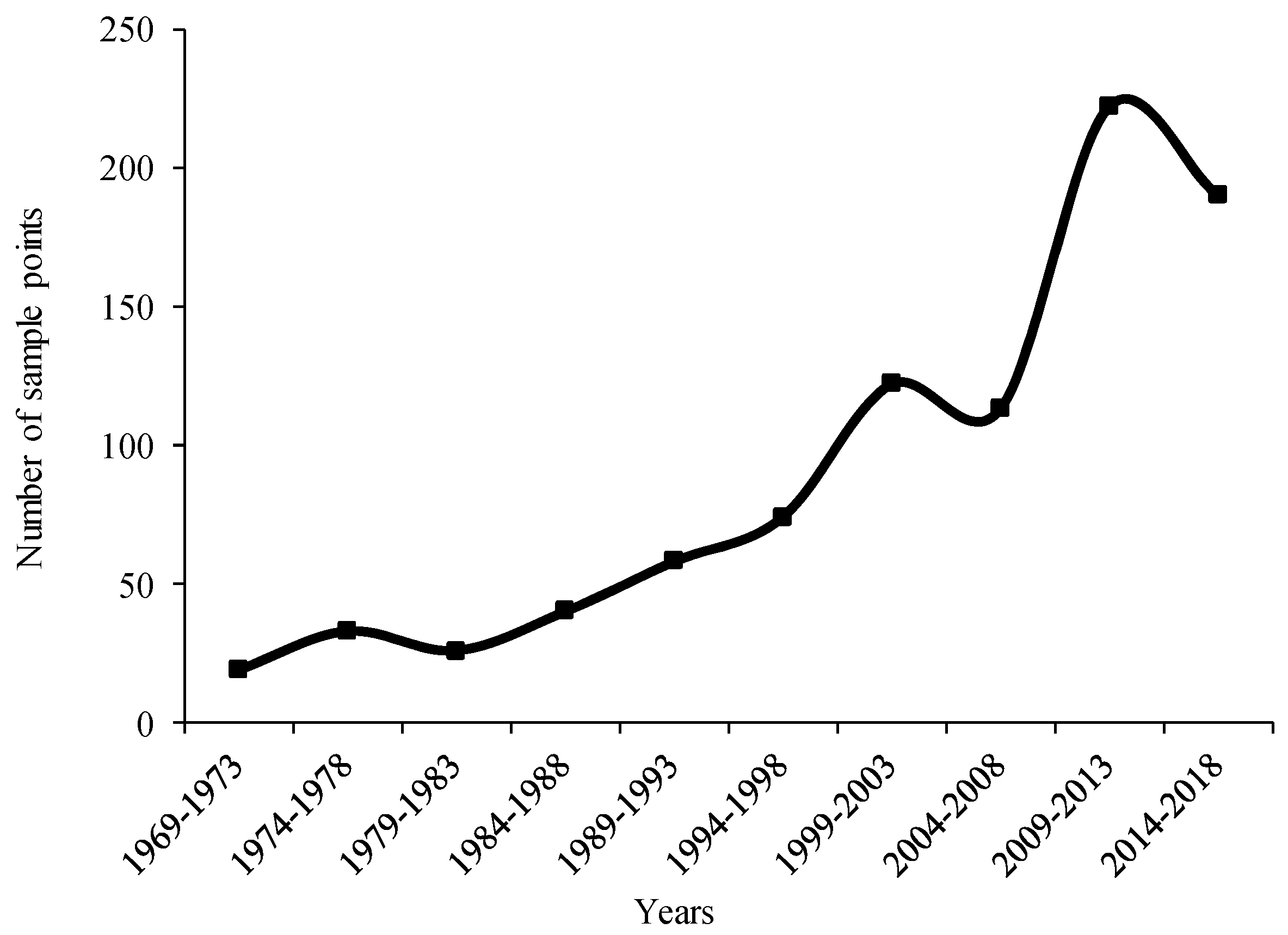
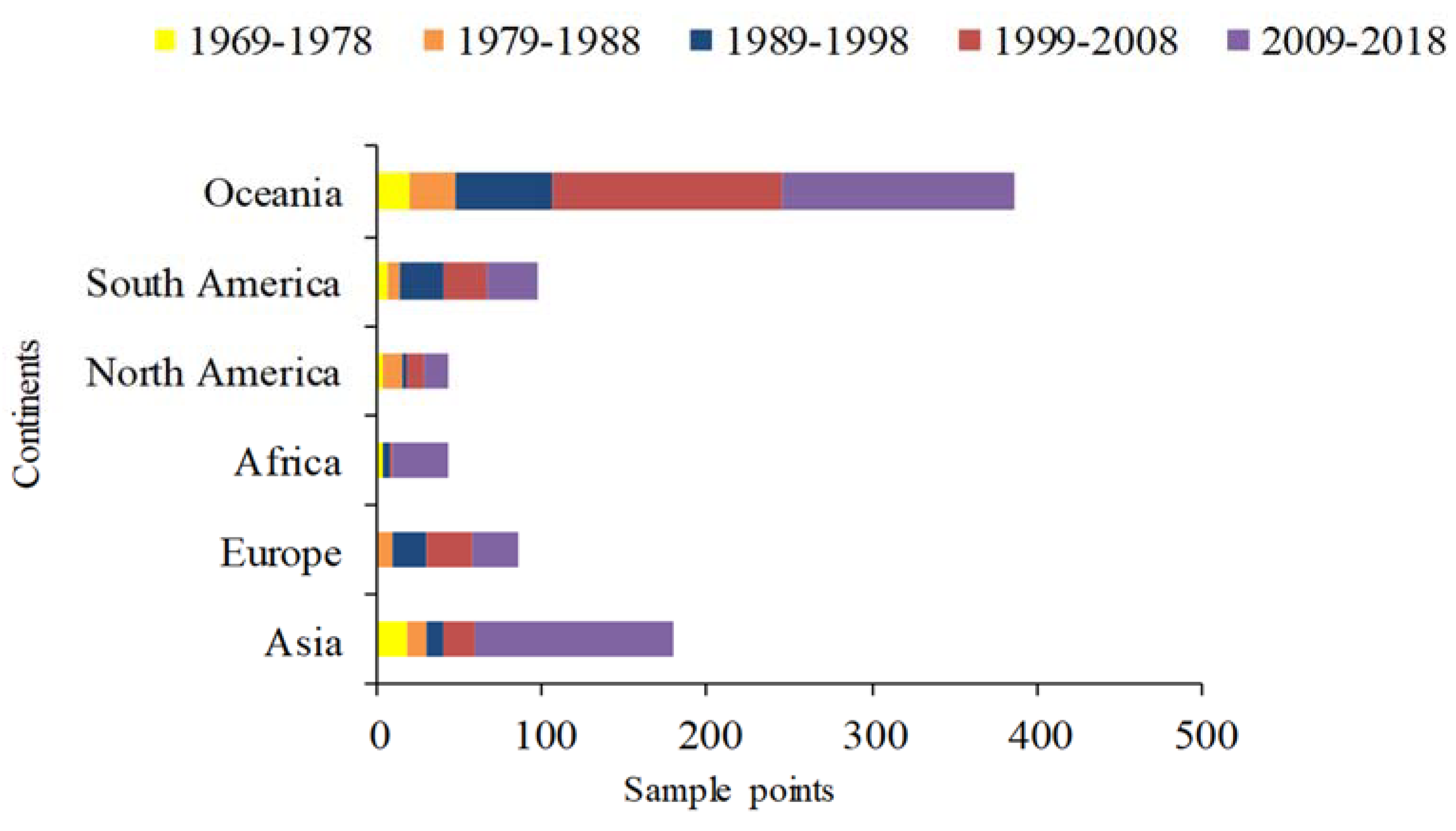
2.1. Species Distribution
2.2. Environmental Data
2.3. Random Forest Model
2.4. Classification of Predictive Distribution
3. Results
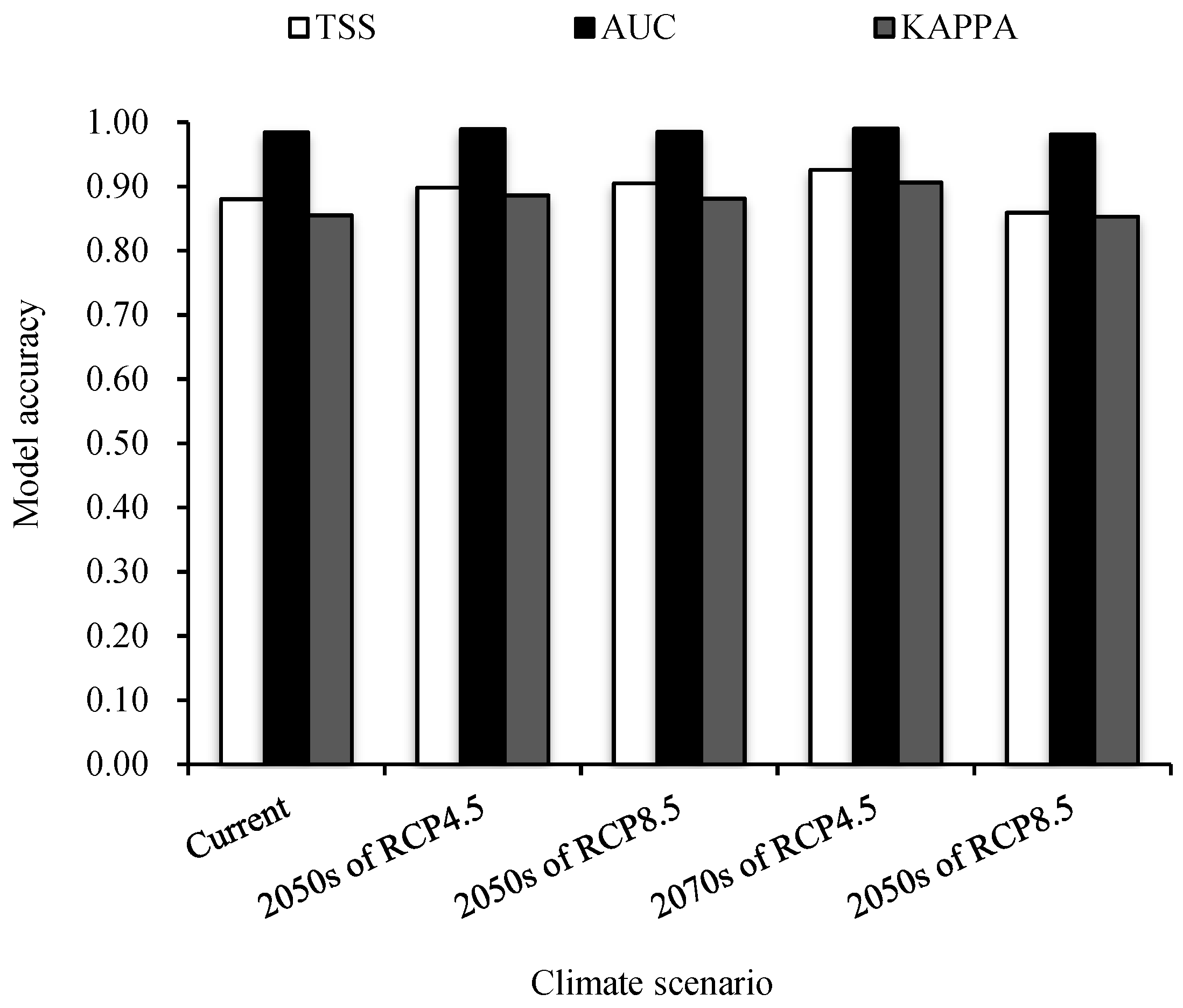
3.1. Precision Test of Random Forest Model
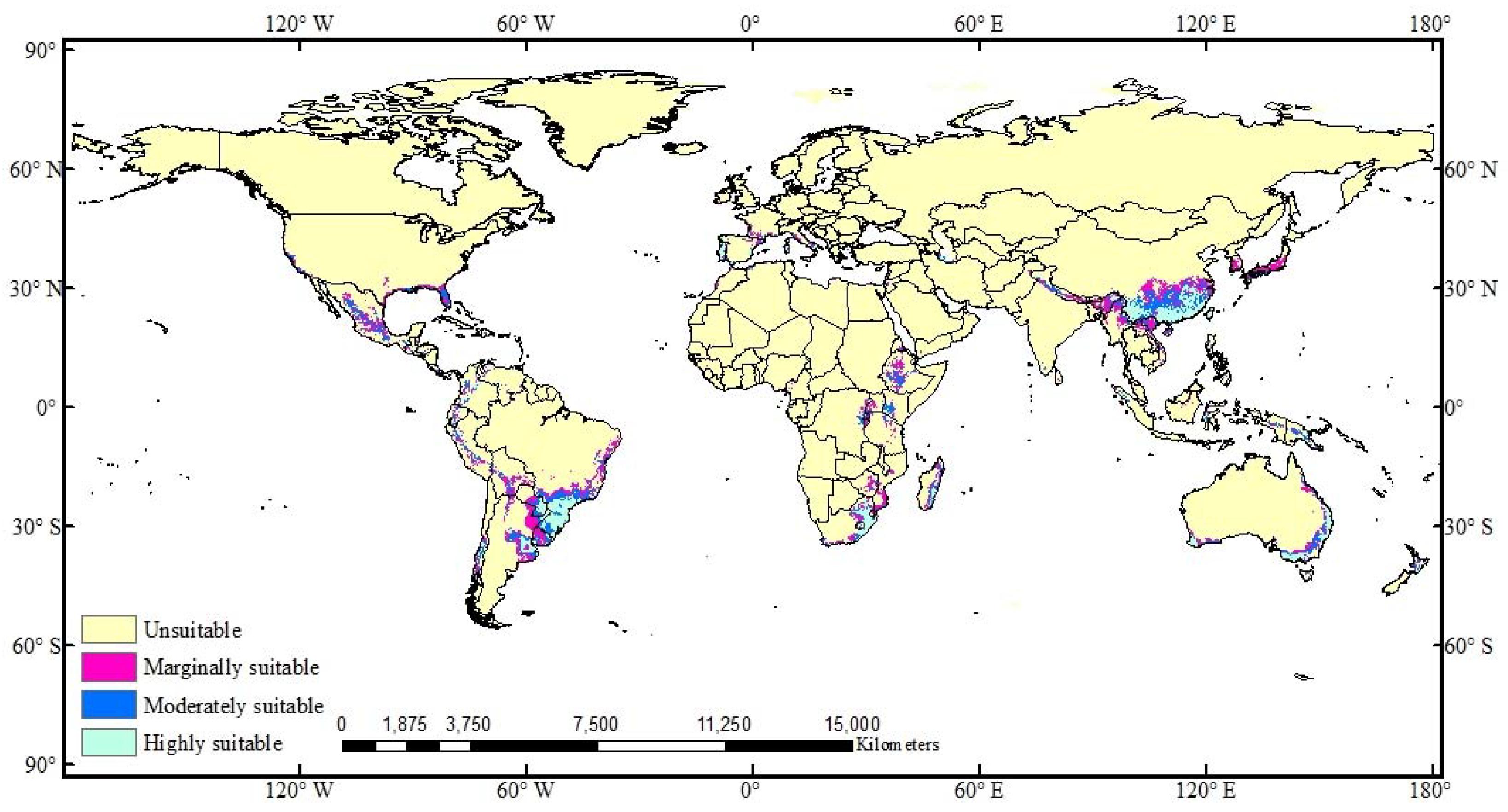
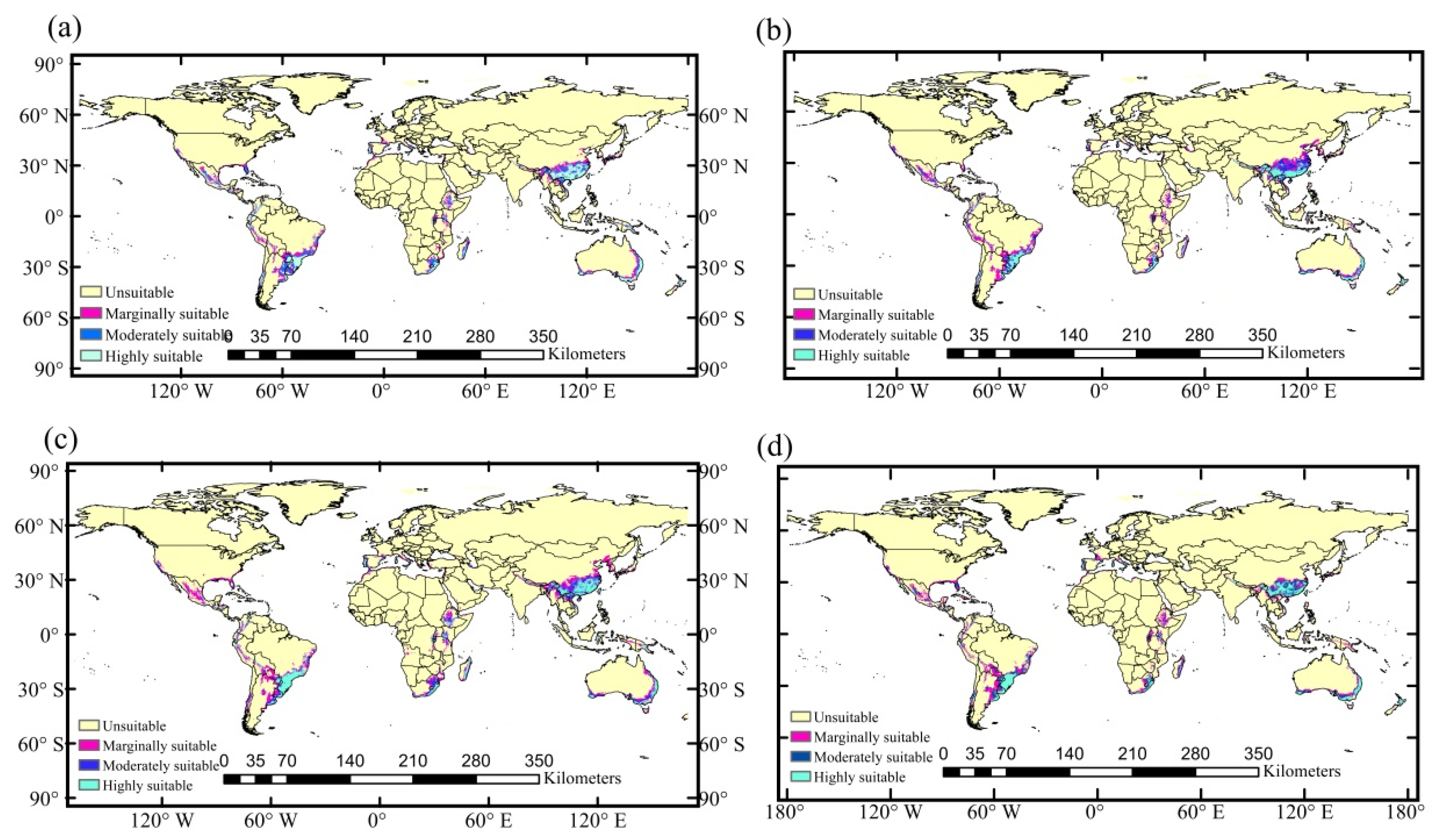
3.2. Potential Global Distribution
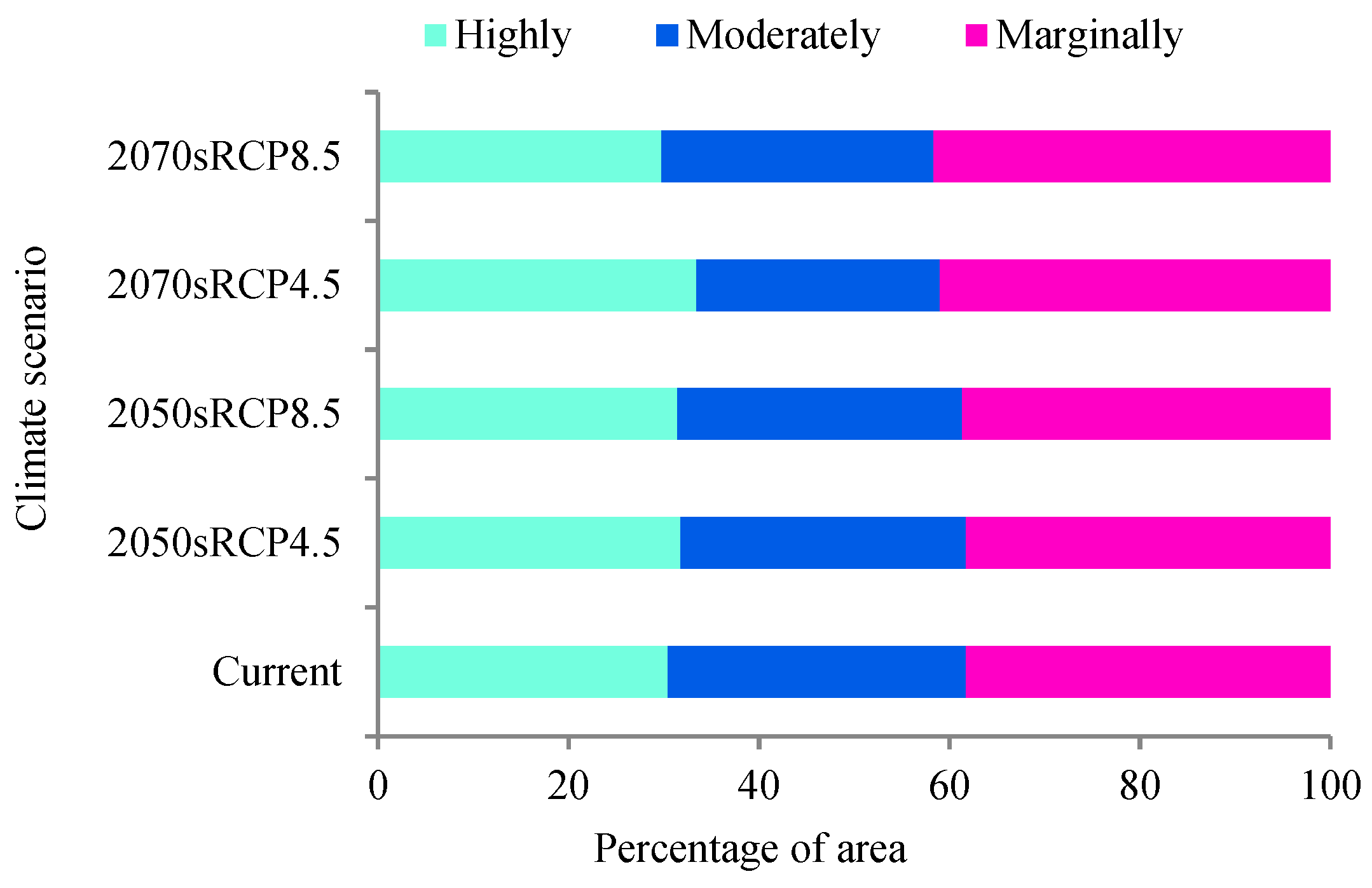
3.3. Predicting Potential Suitable Habitats under Future Climate Scenarios
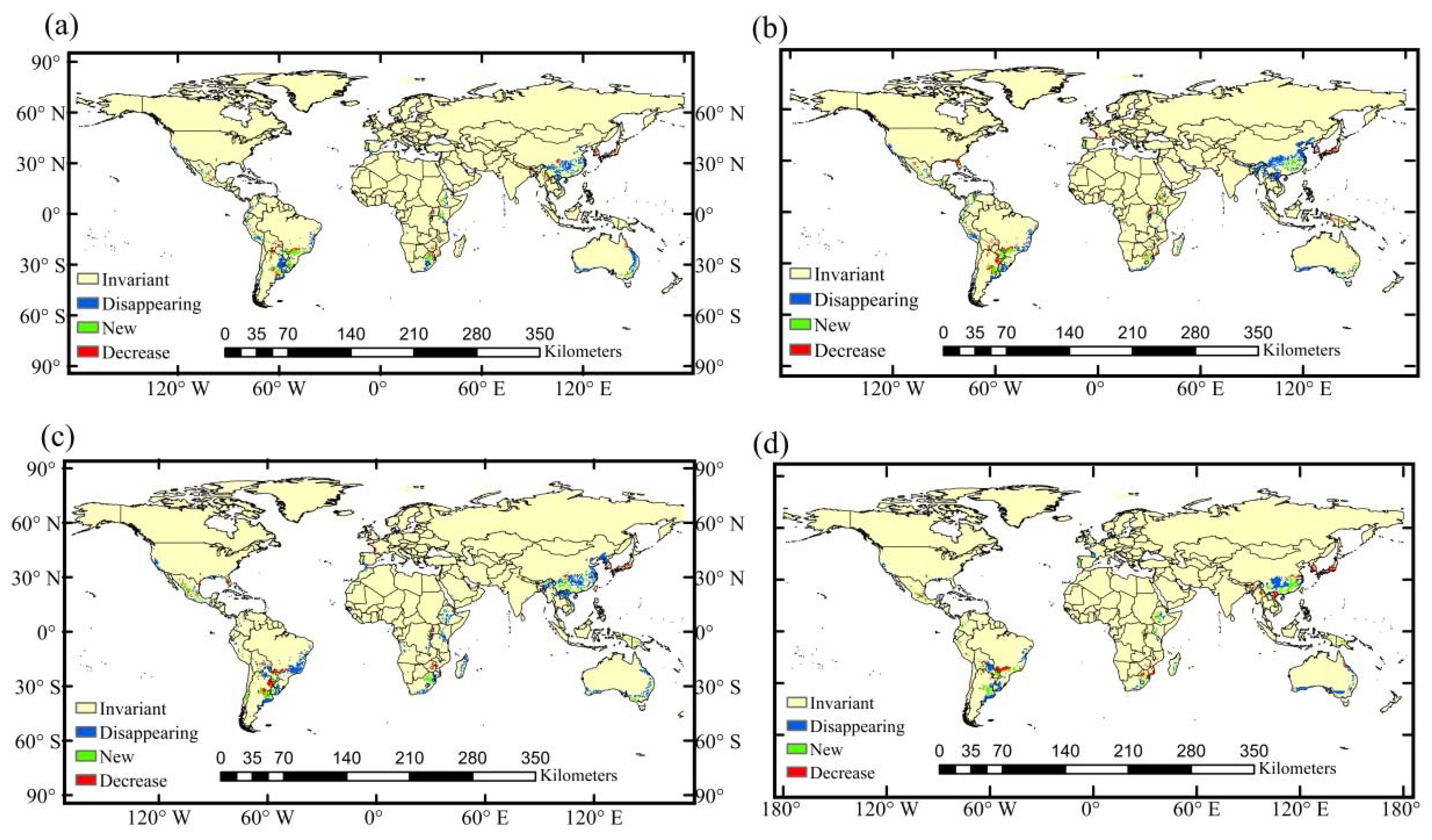
3.4. Potential Suitable Habitat Change in the Future
3.5. Relative Importance of Climatic Factors
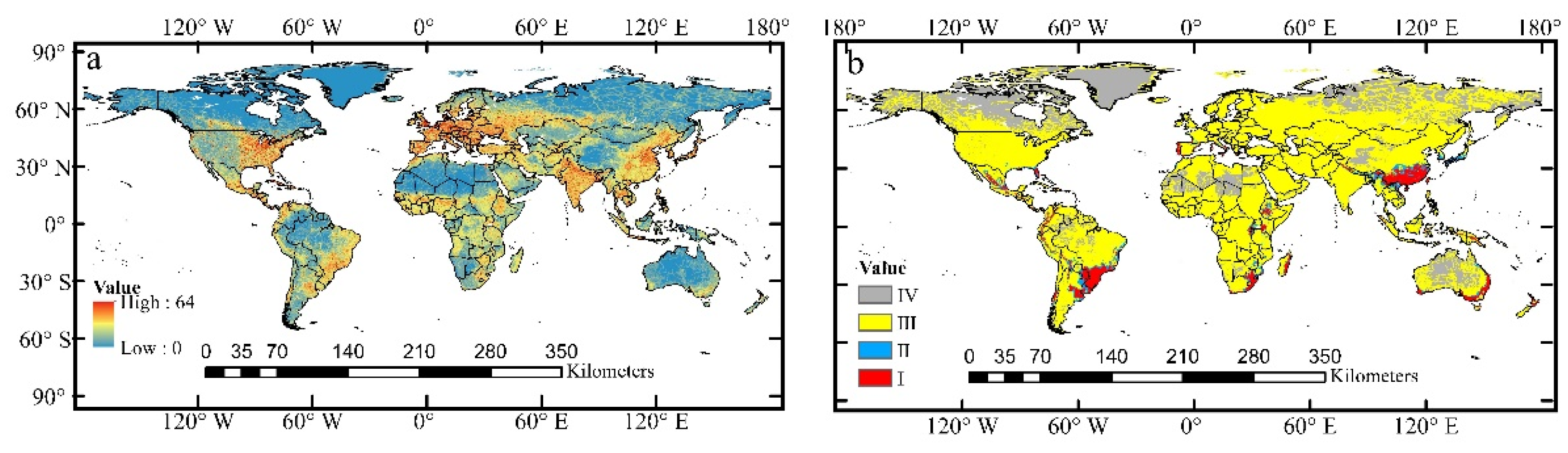
3.6. Relationships between Human Activities and Distribution
4. Discussion
4.1. The Significance of Studying the Potential Distribution of A. Cordifolia
4.2. The Influence of Major Variables on Adaptation
4.3. The Dispersal of A. Cordifolia and Human Activities
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- González-Moreno, P.; Pino, J.; Carreras, D.; Basnou, C.; Fernández-Rebollar, I.; Vilà, M. Quantifying the landscape influence on plant invasions in Mediterranean coastal habitats. Landsc. Ecol. 2013, 28, 891–903. [Google Scholar] [CrossRef]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef]
- Sanjay, J.; Krishnan, R.; Shrestha, A.B.; Rajbhandari, R.; Ren, G.Y. Downscaled climate change projections for the Hindu Kush Himalayan region using CORDEX South Asia regional climate models. Adv. Clim. Chang. Res. 2017, 8, 185–198. [Google Scholar] [CrossRef]
- Hierro, J.L.; Villarreal, D.; Eren, Ö.; Graham, J.; Callaway, R.M. Disturbance facilitates invasion: The effects are stronger abroad than at home. Am. Nat. 2006, 168, 144–156. [Google Scholar] [CrossRef]
- Allen, S.K.; Plattner, G.K.; Nauels, A.; Xia, Y.; Stocker, T.F. Climate change 2013: The physical science basis. An overview of the working group 1 contribution to the fifth assessment report of the intergovernmental panel on climate change (IPCC). Comp. Geom. 2007, 18, 95–123. [Google Scholar] [CrossRef]
- Climate Change 2014 Synthesis Report. Available online: https://www.cornwallhousing.org.uk/media/10924486/IPCC-Climate-Change-2014-Synthesis-Report.pdf (accessed on 30 January 2020).
- Linares, J.C.; Covelo, F.; Carreira, J.A.; Merino, J. Phenological and water-use patterns underlying maximum growing season length at the highest elevations: Implications under climate change. Tree Physiol. 2012, 32, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Vasseur, L.; You, M. Potential distribution of the invasive loblolly pine mealybug, Oracella acuta (Hemiptera: Pseudococcidae), in Asia under future climate change scenarios. Clim. Chang. 2017, 141, 719–732. [Google Scholar] [CrossRef]
- Jiang, F. Bioclimatic and altitudinal variables influence the potential distribution of canine parvovirus type 2 worldwide. Ecol. Evol. 2018, 8, 4534–4543. [Google Scholar] [CrossRef]
- Zhao, Z.F.; Wei, H.Y.; Guo, Y.L.; Gu, W. Potential distribution of Panax ginseng and its predicted responses to climate change. Chin. J. Appl. Ecol. 2016, 27, 3607–3615. (In Chinese) [Google Scholar] [CrossRef]
- Guo, Y.; Wei, H.; Lu, C.; Gao, B.; Gu, W. Predictions of potential geographical distribution and quality of Schisandra sphenanthera under climate change. Peerj 2016, 4, e2554. [Google Scholar] [CrossRef]
- Zhang, L.P.; Li, L.C.; Xia, J.; Wang, R.C. Quantitative assessment of the impact of climate variability and human activities on runoff change in the Luanhe river catchme. J. Nat. Resour. 2015, 30, 664–672. (In Chinese) [Google Scholar]
- Wauchope, H.S.; Shaw, J.D.; Varpe, Ø.; Lappo, E.G.; Boertmann, D.; Lanctot, R.B.; Fuller, R.A. Rapid climate-driven loss of breeding habitat for arctic migratory birds. Glob. Chang. Biol. 2016, 23, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.; Behera, M.D.; Jacob, J. Predicting the distribution of rubber trees (Hevea brasiliensis) through ecological niche modelling with climate, soil, topography and socioeconomic factors. Ecol. Res. 2016, 31, 75–91. [Google Scholar] [CrossRef]
- O’Donnel, J.; Gallagher, R.; Wilson, P.D.; Downey, P.O.; Hughes, L.; Leishman, M. Invasion hotspots for non-native plants in Australia under current and future climates. Glob. Chang. Biol. 2012, 18, 617–629. [Google Scholar] [CrossRef]
- Rossiter-Rachor, N.A.; Setterfield, S.A.; Douglas, M.M.; Hutley, L.B.; Cook, G.D.; Schmidt, S. Invasive, Andropogon gayanus, (gamba grass) is an ecosystem transformer of nitrogen relations in Australian savanna. Ecol. Appl. 2009, 19, 1546–1560. [Google Scholar] [CrossRef] [PubMed]
- Giakoumi, S.; Guilhaumon, F.; Kark, S.; Terlizzi, A.; Claudet, J.; Felline, S.; Cerrano, C.; Coll, M.; Danovaro, R.; Fraschetti, S.; et al. Space invaders; biological invasions in marine conservation planning. Divers. Distrib. 2016, 22, 1–12. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Walker, L.R.; Whiteaker, L.D.; Mueller-Dombois, D.; Matson, P.A. Biological invasion by Myrica faya alters ecosystem development in Hawaii. Science 1987, 238, 802–804. [Google Scholar] [CrossRef]
- Whitney, K.; Gabler, C. Rapid evolution in introduced species, ’invasive traits’ and recipient communities: Challenges for predicting invasive potential. Divers. Distrib. 2008, 14, 569–580. [Google Scholar] [CrossRef]
- González-Moreno, P.; Diez, J.M.; Ibáñez, I.; Castell, X.F.; Vilà, M. Plant invasions are context-dependent: Multiscale effects of climate, human activity and habitat. Divers. Distrib. 2014, 20, 720–731. [Google Scholar] [CrossRef]
- Hulme, P.E. Relative roles of life-form, land use and climate in recent dynamics of alien plant distributions in the British Isles. Weed Res. 2009, 49, 19–28. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.; Ibañez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [PubMed]
- Shine, K.P.; Forster, P. The effect of human activity on radiative forcing of climate change: A review of recent developments. Glob. Planet. Chang. 1999, 20, 205–225. [Google Scholar] [CrossRef]
- Huang, J.P.; Yu, H.P.; Guan, X.D.; Wang, G.Y.; Guo, R.X. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2015, 6, 166–171. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Richardson, D.M.; Holmes, P.M.; Esler, K.J.; Galatowitsch, S.M.; Stromberg, J.C.; Kirkman, S.P.; Pysek, P.; Hobbs, R.J. Riparian vegetation: Degradation, alien plant invasions, and restoration prospects. Divers. Distrib. 2007, 13, 126–139. [Google Scholar] [CrossRef]
- Weber, E.; Sun, S.G.; Li, B. Invasive alien plants in China: Diversity and ecological insights. Biol. Invasions 2008, 10, 1411–1429. [Google Scholar] [CrossRef]
- Lamsal, P.; Kumar, L.; Aryal, A.; Atreya, K. Invasive alien plant species dynamics in the Himalayan region under climate change. Ambio J. Hum. Environ. 2018, 47, 697–710. [Google Scholar] [CrossRef]
- Luizza, M.W.; Tewodros, W.; Evangelista, P.H.; Jarnevich, C.S. Integrating local pastoral knowledge, participatory mapping, and species distribution modeling for risk assessment of invasive rubber vine (Cryptostegia grandiflora) in Ethiopia’s Afar region. Ecol. Soc. 2016, 21, 22. [Google Scholar] [CrossRef]
- Guilherme, D.O.; Bruno, D.S.B.; Daniela, D.S.D.S.; Vinícius, Q.D.M.; Maria, C.S.S. Combining the effects of biological invasion and climate change into systematic conservation planning for the Atlantic forest. Biol. Invasions 2018, 20, 2753–2765. [Google Scholar] [CrossRef]
- Jing, L.; Yang, Y.; Wei, H.Y.; Zhang, Q.Z.; Zhang, X.H.; Zhang, X.Y.; Gu, W. Assessing habitat suitability of parasitic plant Cistanche deserticola in north China under future climate scenarios. Forests 2019, 10, 823. [Google Scholar] [CrossRef]
- Zhao, Z.F.; Guo, Y.L.; Wei, H.Y.; Qiao, R.; Gu, W. Prediction of potential geographical distribution and quality of a Gynostemma pentaphyllum base on the fuzzy matter element model in China. Sustainability 2017, 9, 1114. [Google Scholar] [CrossRef]
- Wang, R.; Jiang, Y.; Su, P.; Wang, J.A. Global spatial distribution of and trends in rice exposure to high temperature. Sustainability 2019, 11, 6271. [Google Scholar] [CrossRef]
- Ashraf, U.; Ali, H.; Chaudry, M.N.; Ashraf, I.; Batool, A.; Saqib, Z. Predicting the potential distribution of Olea ferruginea in Pakistan incorporating climate change by using Maxent model. Sustainability 2016, 8, 722. [Google Scholar] [CrossRef]
- Austin, M. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol. Model. 2007, 200, 1–19. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Wei, H.; Gao, B.; Gu, W. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom Tricholoma matsutake under multiple climate change scenarios. Sci. Rep. 2017, 7, 46221. [Google Scholar] [CrossRef]
- Gao, B.; Wei, H.Y.; Guo, Y.L.; Gu, W. Using GIS and MaxEnt to analyze the potential distribution of Abies chensiensis. Chin. J. Ecol. 2015, 34, 843–852. (In Chinese) [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Bi, Y.F.; Xu, J.C.; Li, Q.H.; Antoine, G.; Wilfried, T.; Niklaus, E.Z.; Yang, Y.P.; Yang, X.F. Applying biomod for model-ensemble on species distribution: A case study for Tsuga chinensis in China. Plant Divers. Resour. 2013, 35, 647–655. (In Chinese) [Google Scholar] [CrossRef]
- Hill, L.; Hector, A.; Hemery, G.; Smart, S.; Tanadini, M.; Brown, N. Abundance distributions for tree species in Great Britain: A two-stage approach to modeling abundance using species distribution modeling and random forest. Ecol. Evol. 2017, 7, 1043–1056. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.L.; Zhang, X.; Liu, S.R.; Sun, P.S.; Wang, T.L. The basic principle of random forest and its applications in ecology: A case study of Pinus yunnanensis. Acta Ecol. Sin. 2014, 34, 650–659. [Google Scholar] [CrossRef]
- Jin, Y.; Zhou, K.X.; Fang, Y.; Liu, X. Assessment of effect of climate change on potential habitat of animal species based on random forest model. J. Ecol. Rural Environ. 2014, 30, 416–422. (In Chinese) [Google Scholar] [CrossRef]
- Stančić, Z.; Mihelj, D. Anredera cordifolia (Ten.) Steenis (Basellaceae), naturalised in south Croatia. Nat. Croat. 2010, 19, 273–279. [Google Scholar]
- Wang, Y.L.; Wei, M.Y.; Zhao, H. The biological characteristics of alien plant Anredera cordifolia and its control. J. Anhui Agric. Sci. 2008, 36, 5524–5526. (In Chinese) [Google Scholar] [CrossRef]
- Ji, G.X.; Tan, L.L.; Chu, Q.G. Anatomical studies on the distribution and development of mucilage cells in Anredera cordifolia. J. Trop. Subtrop. Bot. 2010, 18, 655–660. (In Chinese) [Google Scholar] [CrossRef]
- Wu, S.H.; Xie, C.F.; Rejmánek, M. Catalogue of the naturalized flora of Taiwan. Taiwania 2004, 49, 16–31. (In Chinese) [Google Scholar] [CrossRef]
- Timmins, S.M.; Reid, V. Climbing asparagus, Asparagus scandens Thunb.: A South African in your forest patch. Austral. Ecol. 2010, 25, 533–538. [Google Scholar] [CrossRef]
- Van Gennip, S.J.; Popova, E.E.; Yool, A.; Pecl, G.; Hobday, A.J.; Sorte, C.J.B. Going with the flow: The role of ocean circulation in global marine ecosystems under a changing climate. Glob. Chang. Biol. 2017, 23, 2602–2617. [Google Scholar] [CrossRef]
- Ling, Y.; Chen, J.L.; Wu, S.Y.; Chen, Y.Z. The test of Anredera cordifolia on nutrient quality and acute toxicity. Clin. Med. Eng. 2010, 17, 57–58. (In Chinese) [Google Scholar]
- Nuryantini, A.Y.; Edikresnha, D.; Munir, M.M. Electrospun polyvinylpyrrolidone as a carrier for leaves extracts of Anredera cordifolia (Ten.) Steenis. Mater. Sci. Forum 2015, 827, 91–94. [Google Scholar] [CrossRef]
- Vivian-Smith, G.; Lawson, B.E.; Turnbull, I.; Downey, P.O. The biology of Australian weeds. 46. Anredera cordifolia (Ten.) Steenis. Plant Protect. Quart. 2007, 22, 2–10. [Google Scholar]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. Biomod—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Bae, M.J.; Hwang, S.J.; Kim, S.H.; Park, Y.S. Predicting potential impacts of climate change on freshwater fish in Korea. Ecol. Inform. 2014, 29, 156–165. [Google Scholar] [CrossRef]
- Hipólito, J.; Hasui, É.; Viana, B.F. Solving problems involving the distribution of a species of unknown distribution via ecological niche modeling. Nat. Conserv. 2015, 13, 15–23. [Google Scholar] [CrossRef]
- Purves, R.D. Optimum numerical integration methods for estimation of area-under-the-curve (AUC) and area-under-the-moment-curve (AUMC). J. Pharmacokinet. Biop. 1992, 20, 211–226. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (Tss). J. Appl. Ecol. 2010, 43, 1223–1232. [Google Scholar] [CrossRef]
- Chu, J.; Li, Y.; Zhang, L.; Li, B.; Gao, M.; Tang, X.; Ni, J.; Xu, X. Potential distribution range and conservation strategies for the endangered species Amygdalus pedunculata. Biodivers. Sci. 2017, 25, 799–806. (In Chinese) [Google Scholar] [CrossRef][Green Version]
- Lu, C.Y.; Gu, W.; Dai, A.H.; Wei, H.Y. Assessing habitat suitability based on geographic information system (GIS) and fuzzy: A case study of Schisandra sphenanthera Rehd. et Wils. in Qinling Mountains, China. Ecol. Model. 2012, 242, 105–115. [Google Scholar] [CrossRef]
- Starr, F.; Starr, K.; Loope, L. Anredera cordifolia (Madeira vine), Basellaceae. Report from Biological Resources Division, United States Geological Survey, Maui, Hawaii, 2003. Available online: http://www.hear.org/starr/hiplants/reports/pdf/anredera_cordifolia.pdf (accessed on 30 January 2020).
- Keller, R.P.; Geist, J.; Jeschke, J.M.; Kühn, I. Invasive species in Europe: Ecology, status, and policy. Environ. Sci. Eur. 2011, 23, 23. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of aline plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Xifreda, C.C.; Argimon, S.; Wulff, A.F. Infraspecific characterization and chromosome numbers in Anredera cordifolia (Basellaceae). Thais. J. Bot. 1999, 9, 99–108. [Google Scholar] [CrossRef]
- Callaway, R.M.; Cipollini, D.; Barto, K.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Stinson, K.; Klironomos, J. Novel weapons: Invasive plant suppresses fungal mutualists in America but not in its native Europe. Ecology 2008, 89, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Mahall, B.E.; Wicks, C.; Zabinski, P.C. Soil fungi and the effects of an invasive forb on grasses: Neighbor identity matters. Ecology 2003, 84, 129–135. [Google Scholar] [CrossRef]
- Widrlechner, M.P.; Daly, C.; Keller, M.; Kaplan, K. Horticultural applications of a newly revised USDA Plant Hardiness Zone Map. HortTechnology 2012, 22, 6–19. [Google Scholar] [CrossRef]
- Mather, J.R.; Yoshioka, G.A. The role of climate in the distribution of vegetation. Ann. Assoc. Am. Geogr 1968, 58, 29–41. [Google Scholar] [CrossRef]
- Stephenson, N.L. Climatic control of vegetation distribution: The role of water balance. Am. Nat. 1990, 135, 649–670. [Google Scholar] [CrossRef]
- Song, S.; Li, F.D.; Lu, Y.L.; Khan, K.; Xue, J.F.; Leng, P.F. Spatio-temporal characteristics of the extreme climate events and their potential effects on crop yield in Ethiopia. J. Resour. Ecol. 2018, 9, 290–301. [Google Scholar] [CrossRef]
- Tandazo-Yunga, J.V.; Ruiz-González, M.X.; Rojas, J.R.; Capa-Mora, E.D.; Prohens, J.; Alejandro, J.D.; Acosta-Quezada, P.G. The impact of an extreme climatic disturbance and different fertilization treatments on plant development, phenology, and yield of two cultivar groups of Solanum betaceum Cav. PLoS ONE 2017, 12, e0190316. [Google Scholar] [CrossRef]
- Sirgedaite-Šežiene, V.; Baležentiene, L.; Varnagiryte-Kabašinskiene, I.; Stakėnas, V.; Baliuckas, V. Allelopathic effects of dominant ground vegetation species on initial growth of Pinus sylvestris L. seedlings in response to different temperature scenarios. iForest 2019, 12, 132–140. [Google Scholar] [CrossRef]
- Baležentienė, L. Allelopathic activity of two invasive Impatiens species in temperate climate of Lithuania. Allelopath. J. 2018, 45, 45–54. [Google Scholar] [CrossRef]
- Šežiene, V.; Baležentienė, L.; Maruška, A. Identification and allelochemic activity of phenolic compounds in extracts from the dominant plant species established in clear-cuts of Scots pine stands. iForest 2017, 10, 309–314. [Google Scholar] [CrossRef]
- Westhuizen, L.V.D. Initiation of a biological control programme against Madeira vine, Anredera cordifolia (Ten.) Steenis (Basellaceae), in South Africa. Afr. Entomol. 2011, 19, 217–222. [Google Scholar] [CrossRef]
- Cagnotti, C.; Kay, F.M.; Gandolfo, D. Biology and host specificity of Plectonycha correntina Lacordaire (Chrysomelidae), a candidate for the biological control of Anredera cordifolia (Tenore) Steenis (Basellaceae). Afr. Entomol. 2007, 15, 300–309. [Google Scholar] [CrossRef]
- Batianoff, G.N.; Butler, D.W. Assessment of invasive naturalized plants in south-east Queensland. Plant Prot. Q. 2002, 17, 27–34. [Google Scholar]
- Loope, L.; Starr, F.; Starr, K. Protecting endangered plant species from displacement by invasive plants on Maui, Hawaii. Weed Technol. 2004, 18, 1472–1474. [Google Scholar] [CrossRef]
- Armstrong, T.; Prior, S. Control of Madeira vine (Anredera cordifolia) using mechanical methods, herbicides and hot fire. In Alan Fletcher Research Station: Environmental Weed Management Research; Queensland Department of Natural Resources: Brisbane, Australia, 1997; pp. 37–43. [Google Scholar]
- Zhang, Y.T.; Huang, J.P.; Guan, X.D.; Guo, R.X. Quantitative assessment of global human activity’s influence on climate. J. Arid. Meteorol. 2017, 35, 182–189. (In Chinese) [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Abbreviation | Name | Selected |
|---|---|---|---|
| Climate variables | Bio1 | Annual mean air temperature | √ |
| Bio2 | Mean diurnal range (Mean of monthly (max temp-min temp)) | ||
| Bio3 | Isothermality (Bio2/Bio7) (*100) | √ | |
| Bio4 | Temperature seasonality (standard deviation *100) | √ | |
| Bio5 | Max temperature of the warmest month | √ | |
| Bio6 | Min temperature of coldest month | √ | |
| Bio7 | Temperature annual range (Bio5-Bio6) | √ | |
| Bio8 | Mean temperature of wettest quarter | ||
| Bio9 | Mean temperature of driest quarter | ||
| Bio10 | Mean temperature of warmest quarter | √ | |
| Bio11 | Mean temperature of coldest quarter | √ | |
| Bio12 | Annual precipitation | ||
| Bio13 | Precipitation of wettest month | ||
| Bio14 | Precipitation of driest month | √ | |
| Bio15 | Precipitation Seasonality (Coefficient of Variation) | ||
| Bio16 | Precipitation of the wettest quarter | √ | |
| Bio17 | Precipitation of the driest quarter | √ | |
| Bio18 | Precipitation of the warmest quarter | √ | |
| Bio19 | Precipitation of the coldest quarter | √ | |
| Human activity | HFP | Human footprint data set |
| Index | Extremely High | Very High | High | Average | Fail | References |
|---|---|---|---|---|---|---|
| AUC | 0.91–1 | 0.81–0.90 | 0.71–0.80 | 0.61–0.70 | <0.60 | [57] |
| TSS | 0.81–1 | 0.80–0.61 | 0.41–0.60 | 0.40–0.21 | <0.20 | [58] |
| Kappa | 0.86–1 | 0.71–0.85 | 0.56–0.70 | 0.41–0.55 | <0.40 | [59] |
| Periods | Climate Scenario | Area Percentage | |||
|---|---|---|---|---|---|
| New | Invariant | Decrease | Disappearing | ||
| Future (2050s) | RCP4.5 | 2.32 | 95.92 | 1.01 | 0.75 |
| RCP8.5 | 2.50 | 95.31 | 1.16 | 1.03 | |
| Future (2070s) | RCP4.5 | 2.75 | 95.52 | 1.01 | 0.72 |
| RCP8.5 | 2.21 | 95.43 | 1.28 | 1.08 | |
| Factor | Current | 2050sRCP4.5 | 2050sRCP8.5 | 2070sRCP4.5 | 2070sRCP8.5 |
|---|---|---|---|---|---|
| Bio11 | 22.66 | 23.04 | 22.87 | 23.46 | 14.43 |
| Bio5 | 14.03 | 11.16 | 19.45 | 18.41 | 14.43 |
| Bio1 | 13.31 | 12.64 | 8.86 | 13.00 | 20.43 |
| Bio7 | 11.15 | 17.84 | 14.68 | 13.00 | 14.79 |
| Bio19 | 11.15 | 8.92 | 4.09 | 6.50 | 4.93 |
| Bio4 | 7.55 | 7.43 | 6.82 | 6.13 | 3.87 |
| Bio6 | 5.40 | 4.46 | 9.55 | 6.13 | 10.92 |
| Bio3 | 3.95 | 3.72 | 3.42 | 1.80 | 2.81 |
| Bio18 | 2.52 | 1.49 | 2.74 | 2.53 | 2.11 |
| Bio16 | 2.52 | 2.22 | 2.06 | 1.81 | 1.41 |
| Bio10 | 2.16 | 1.49 | 2.05 | 2.18 | 2.11 |
| Bio14 | 1.80 | 3.34 | 1.36 | 2.52 | 5.28 |
| Bio17 | 1.80 | 2.25 | 2.05 | 2.53 | 2.48 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wei, H.; Zhao, Z.; Liu, J.; Zhang, Q.; Zhang, X.; Gu, W. The Global Potential Distribution of Invasive Plants: Anredera cordifolia under Climate Change and Human Activity Based on Random Forest Models. Sustainability 2020, 12, 1491. https://doi.org/10.3390/su12041491
Zhang X, Wei H, Zhao Z, Liu J, Zhang Q, Zhang X, Gu W. The Global Potential Distribution of Invasive Plants: Anredera cordifolia under Climate Change and Human Activity Based on Random Forest Models. Sustainability. 2020; 12(4):1491. https://doi.org/10.3390/su12041491
Chicago/Turabian StyleZhang, Xuhui, Haiyan Wei, Zefang Zhao, Jing Liu, Quanzhong Zhang, Xiaoyan Zhang, and Wei Gu. 2020. "The Global Potential Distribution of Invasive Plants: Anredera cordifolia under Climate Change and Human Activity Based on Random Forest Models" Sustainability 12, no. 4: 1491. https://doi.org/10.3390/su12041491
APA StyleZhang, X., Wei, H., Zhao, Z., Liu, J., Zhang, Q., Zhang, X., & Gu, W. (2020). The Global Potential Distribution of Invasive Plants: Anredera cordifolia under Climate Change and Human Activity Based on Random Forest Models. Sustainability, 12(4), 1491. https://doi.org/10.3390/su12041491