Alpine Pioneer Plants in Soil Bioengineering for Slope Stabilization and Restoration: Results of a Preliminary Analysis of Seed Germination and Future Perspectives



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sampling Areas
2.2. Germination Tests
- Addition of 5 mL of distilled water (control);
- Addition of 1 g of CaCO3 and 5 mL of distilled water;
- Addition of 5 mL of a solution containing 100 mg L−1 of gibberellic acid (GA3);
- Addition of 1 g of CaCO3 and 5 mL of a solution containing 100 mg L−1 of GA3.
2.3. Data Analysis
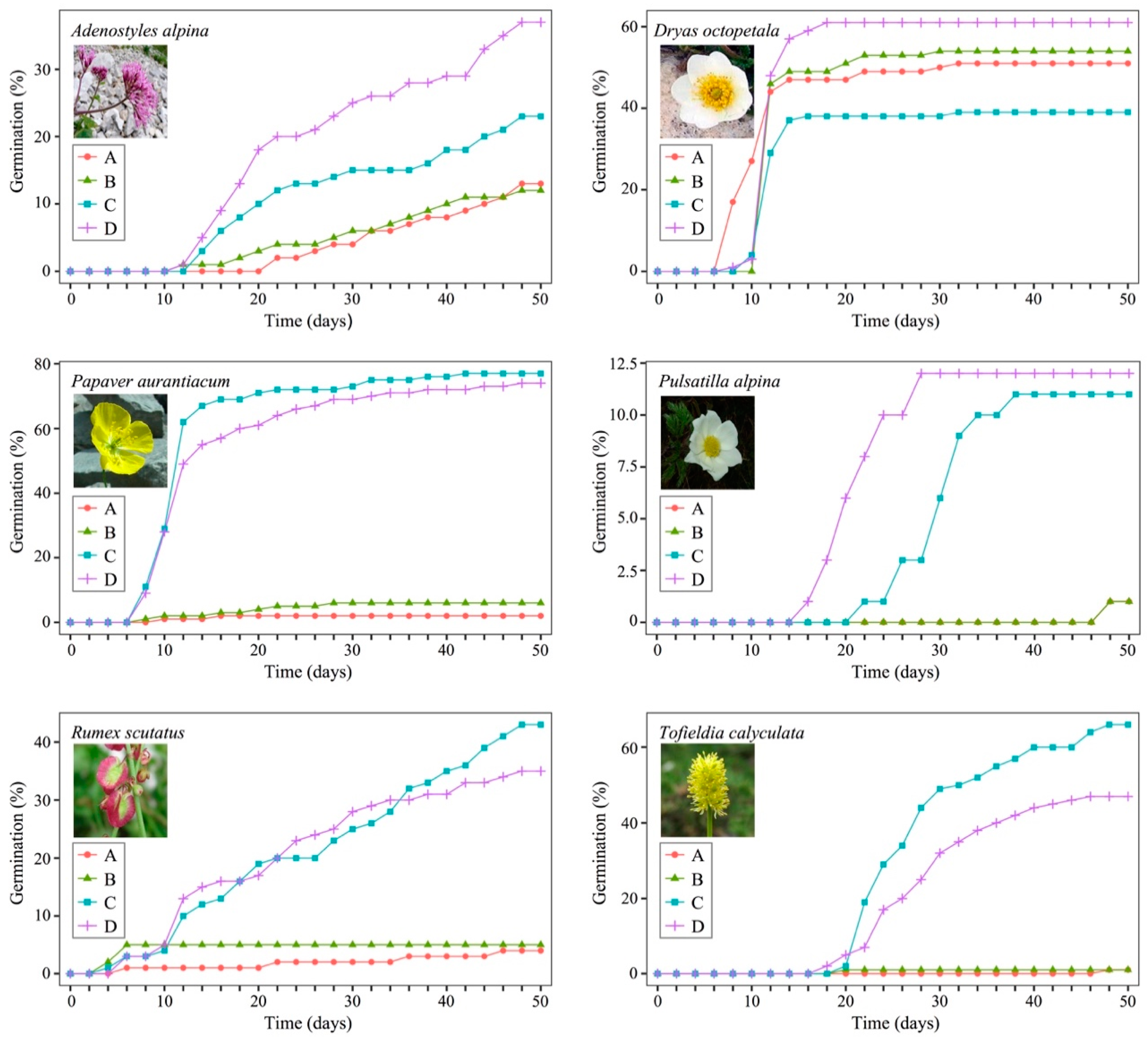
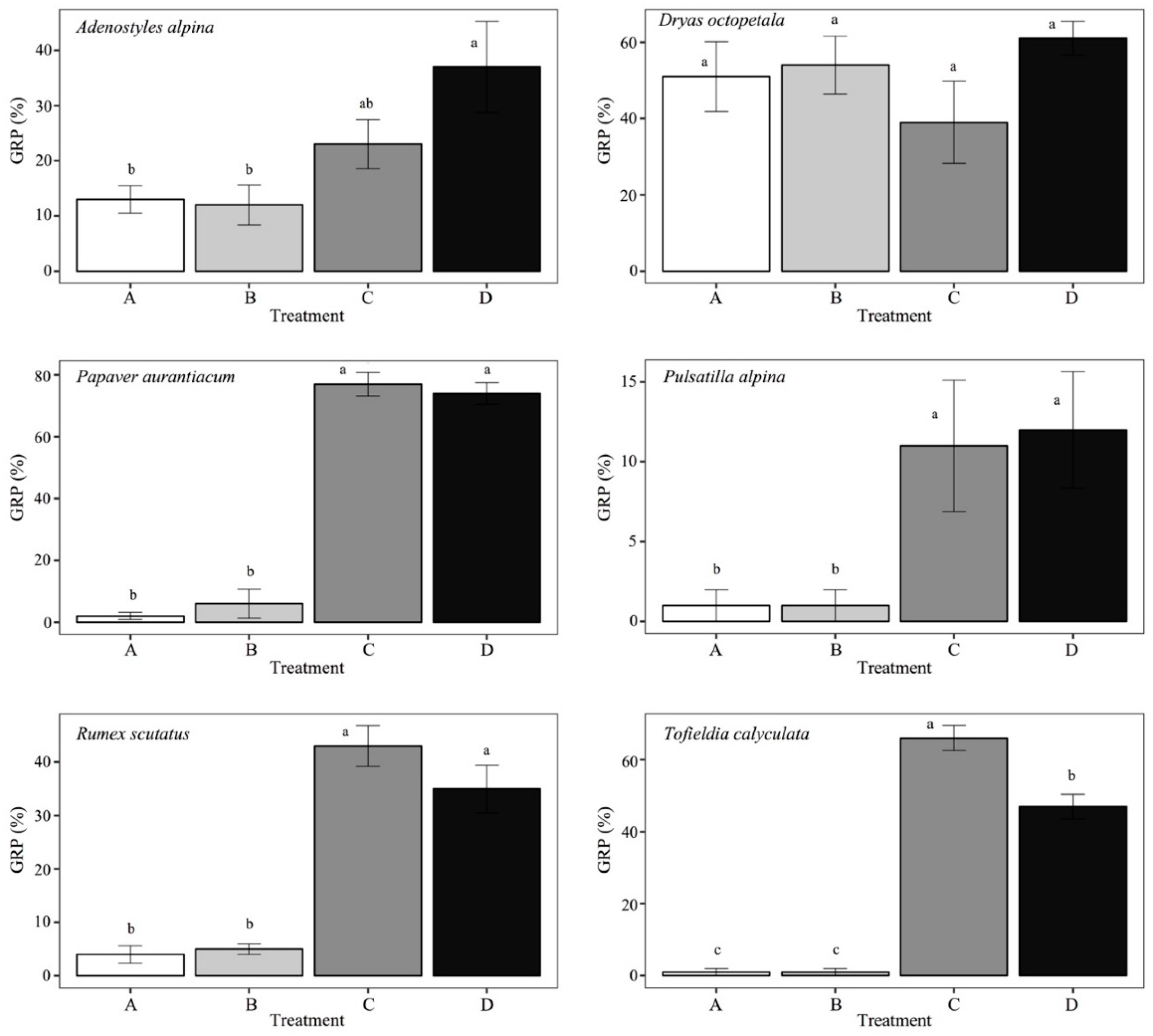
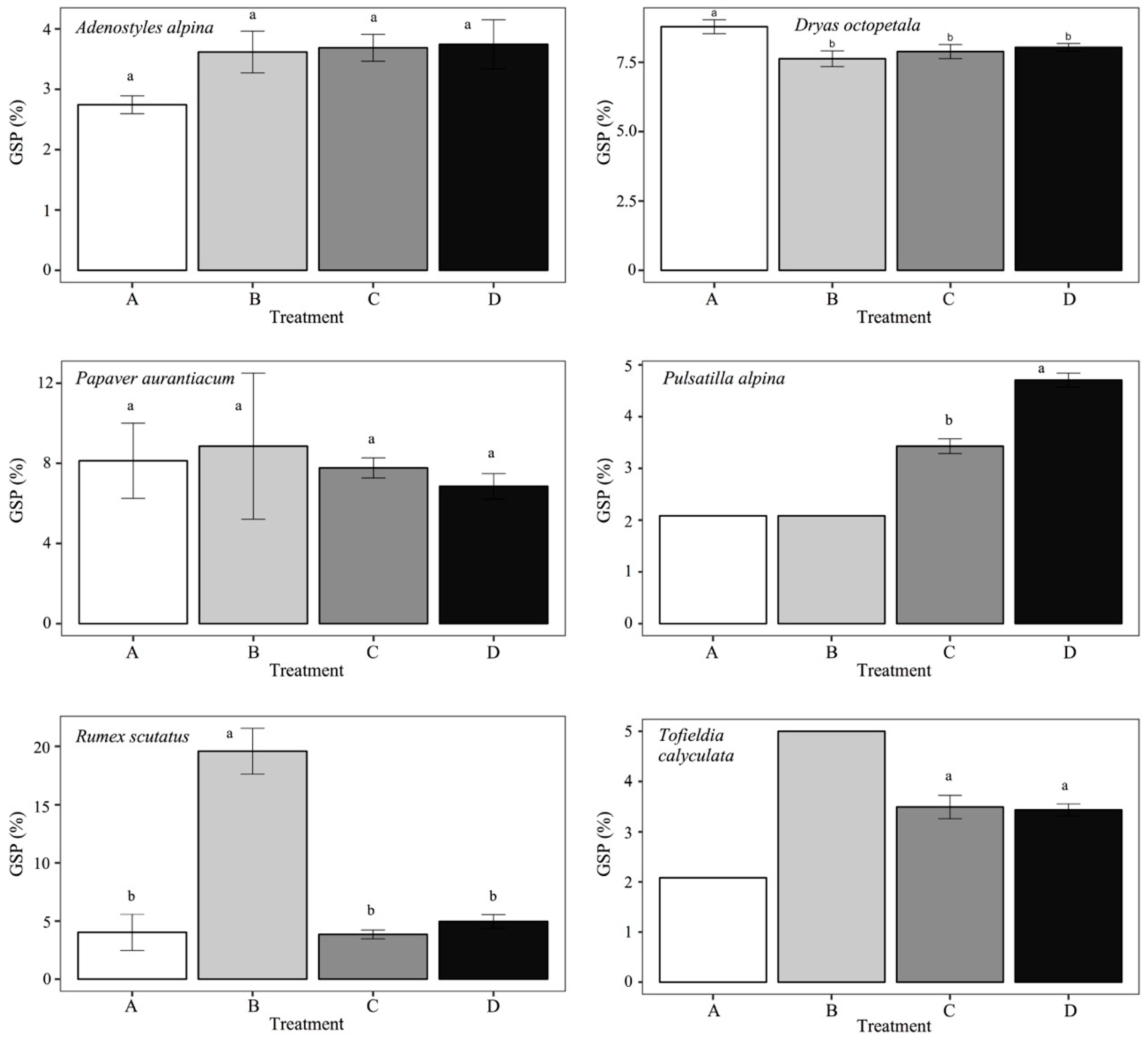
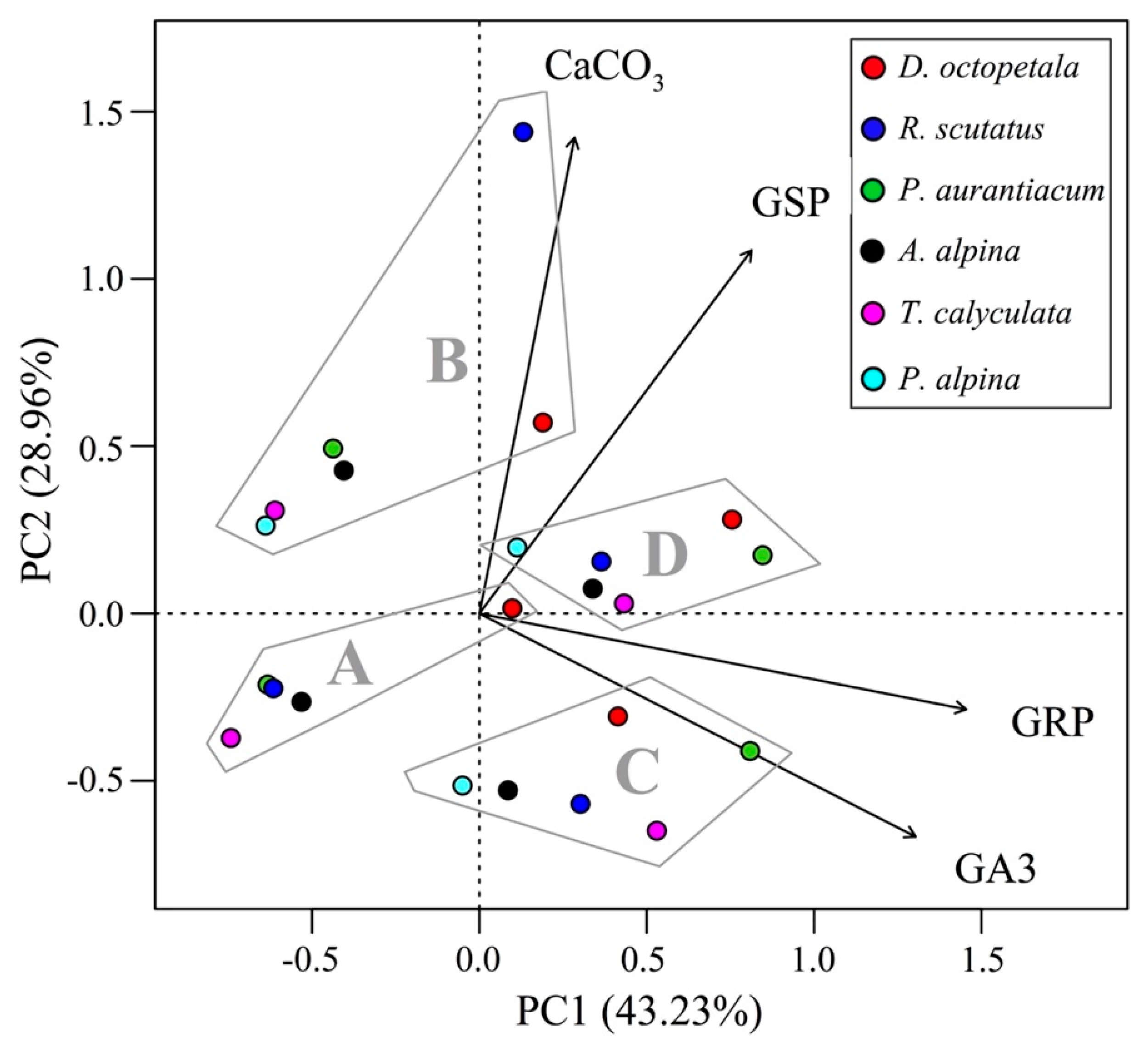
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gariano, S.L.; Guzzetti, F. Landslides in a changing climate. Earth Sci. Rev. 2016, 162, 227–252. [Google Scholar] [CrossRef]
- Yuan, X.C.; Wei, Y.M.; Wang, B. Risk management of extreme events under climate change. J. Clean. Prod. 2017, 166, 1169–1174. [Google Scholar] [CrossRef]
- Einhorn, B.; Eckert, N.; Chaix, C.; Ravanel, L.; Deline, P.; Gardent, M.; Boudières, V.; Richard, D.; Vengeon, J.-M.; Schoeneich, G.G.E.P. Climate change and natural hazards in the Alps. J. Alp. Res. 2015, 103, 1–38. [Google Scholar] [CrossRef]
- Kron, W.; Löw, P.; Kundzewicz, Z. Changes in risk of extreme weather events in Europe. Environ. Sci. Policy 2019, 100, 74–83. [Google Scholar] [CrossRef]
- Nordregio. Mountain Areas in Europe: Analysis of Mountain Areas in EU Member States, Acceding and Other European Countries; Commissioned Report by the European Commission–DG Regional Policy; Nordregio: Brussels, Belgium, 2004. [Google Scholar]
- Keenleyside, C.; Tucker, G.M. Farmland Abandonment in the EU: An Assessment of Trends and Prospects Report Prepared for WWF; Institute for European Environmental Policy: London, UK, 2010. [Google Scholar]
- Terres, J.M.; Nisini, L.; Anguiano, E. Assessing the Risk of Farmland Abandonment in the EU. Final Report; Joint Research Centre: Ispra, Italy, 2013. [Google Scholar]
- Cislaghi, A.; Giupponi, L.; Tamburini, A.; Giorgi, A.; Bischetti, G.B. The effects of mountain grazing abandonment on plant community, forage value and soil properties: Observations and field measurements in an alpine area. Catena 2019, 181, 104086. [Google Scholar] [CrossRef]
- Tanzini, M. Fenomeni Franosi e Opere di Stabilizzazione, 2nd ed.; Dario Flaccovio Editore: Palermo, Italy, 2011; pp. 1–35. [Google Scholar]
- Trigila, A.; Iadanza, C.; Bussettini, M.; Lastoria, B. Dissesto Idrogeologico in Italia: Pericolosità e Indicatori di Rischio–Ed. 2018; Rapporti 287/2018; ISPRA: Roma, Italy, 2018; pp. 3–26. [Google Scholar]
- EU Commission. EUROPE 2020 A Strategy for Smart, Sustainable and Inclusive Growth. 2010. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52010DC2020andfrom=it (accessed on 1 March 2020).
- EU Commission. Roadmap to a Resource Efficient Europe. 2011. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52011DC0571andfrom=IT (accessed on 1 March 2020).
- EEA. Green Infrastructure and Territorial Cohesion. European Environment Agency; Technical Report 18/2011; EEA: Copenhagen, Denmark, 2011. [Google Scholar]
- Baró, F.; Bugter, R.; Gómez-Baggethun, E.; Kopperoinen, L.; Liquete, C.; Potschin, M. Green Infrastructure. In OpenNESS Ecosystem Service Reference Book; EC FP7 Grant Agreement No. 308428; Potschin, M., Jax, K., Eds.; OpenNESS: Belgium, Brussels, 2015. [Google Scholar]
- Schiechtl, H.M. Sicherungsarbeiten im Landschafsbau; Callwey: Munich, Germany, 1973. [Google Scholar]
- Schiechtl, H.M. Bioengineering for Land Reclamation and Conservation; University of Alberta Press: Edmonton, AB, Canada, 1980. [Google Scholar]
- Schiechtl, H.M. Bioingegneria Forestale-Biotecnica Naturalistica; Edizioni Castaldi: Feltre, Italy, 1991. [Google Scholar]
- Bischetti, G.B.; Di Fi Dio, M.; Florineth, F. On the origin of soil bioengineering. Landsc. Res. 2014, 39, 583–595. [Google Scholar] [CrossRef]
- Studer, R.; Zeh, H. Soil Bioengineering. Construction Type Manual; Hochschulverlag: Zurich, Switzerland, 2014. [Google Scholar]
- Giupponi, L.; Bischetti, G.B.; Giorgi, A. Ecological index of maturity to evaluate the vegetation disturbance of areas affected by restoration work: A practical example of its application in an area of the southern alps. Restor. Ecol. 2015, 23, 635–644. [Google Scholar] [CrossRef]
- Giupponi, L.; Bischetti, G.B.; Giorgi, A. A proposal for assessing the success of soil bioengineering work by analysing vegetation: Results of two case studies in the Italian Alps. Landsc. Ecol. Eng. 2017, 13, 305–318. [Google Scholar] [CrossRef]
- Arques, S. Géodynamique, Colonisation Végétale et Phytodiversité des talus D’éboulis dans le Massif de la Grande Chartreuse (Pre’alpes Fran¸caises du Nord). Caractéristiques Géo-Écologiques et Sensibilité aux Changements Environnementaux. Ph.D. Thesis, Joseph Fourier University, Institut de Géographie Alpine, Grenoble, France, 2006. [Google Scholar]
- Beaudiére, A. Modes de Perception de L’éboulis par les Botanistes; Bulletin de l’Association de Géographes FranÇais: Paris, France, 1983; Volume 491, pp. 25–32. [Google Scholar]
- Pérez, F.L. Geobotanical influence of talus movement on the distribution of caulescent Andean rosettes. Flora 1994, 189, 353–371. [Google Scholar] [CrossRef]
- Behre, C.H. Talus behavior above timber in the Rocky Mountains. J. Geol. 1933, 41, 622–635. [Google Scholar] [CrossRef]
- Rahn, P.H. The relationship between natural forested slopes and angles of repose for sand and gravel. Geol. Soc. Am. 1969, 80, 2123–2128. [Google Scholar] [CrossRef]
- Sigafoos, R. Soil instability in tundra vegetation. Ohio J. Sci. 1951, 51, 281–298. [Google Scholar]
- Smith, J. Some moving soils in Spitsbergen. J. Soil Sci. 1956, 7, 10–21. [Google Scholar] [CrossRef]
- Howard, J.A.; Mitchell, C.W. Phytogeomorphology; Wiley: New York, NY, USA, 1985. [Google Scholar]
- Hupp, C.R.; Osterkamp, W.R.; Howard, A.D. Preface (to Binghamton Symposium volume on Biogeomorphology). Geomorphology 1995, 13, Vii–Viii. [Google Scholar] [CrossRef]
- Phillips, J.D. Soils as extended phenotypes. Geoderma 2009, 149, 143–151. [Google Scholar] [CrossRef]
- Giupponi, L.; Borgonovo, G.; Giorgi, A.; Bischetti, G.B. How to renew soil bioengineering for slope stabilization: Some proposals. Landsc. Ecol. Eng. 2019, 15, 37–50. [Google Scholar] [CrossRef]
- Dierschke, H. Pflanzensoziologie: Grundlagen und Methoden; UTB: Stuttgart, Germany, 1994. [Google Scholar]
- Biondi, E. Phytosociology today: Methodological and conceptual evolution. Plant Biosyst. 2011, 145, 19–29. [Google Scholar] [CrossRef]
- Giupponi, L.; Bischetti, G.B.; Giorgi, A. Vegetation analysis and estimation of forest reconstitution time in protected areas of Val Camonica (Southern Alps) where a commercial mixture of seeds was sown. J. Prot. Mt. Areas Res. Manag. 2017, 9, 22–29. [Google Scholar] [CrossRef]
- Aavik, T.; Edwards, P.J.; Holderegger, R.; Graf, R.; Billeter, R. Genetic consequences of using seed mixtures in restoration: A case study of a wetland plant Lychnis flos-cuculi. Biol. Conserv. 2012, 145, 195–204. [Google Scholar] [CrossRef]
- Hagen, D.; Hansen, T.I.; Graae, B.J.; Rydgren, K. To seed or not to seed in alpine restoration: Introduced grass species outcompete rather than facilitate native species. Ecol. Eng. 2014, 64, 255–261. [Google Scholar] [CrossRef]
- Krautzer, B.; Hacker, E. Soil-Bioengineering: Ecological Restoration with Native Plant and Seed Material Conference. In Proceedings of the Soil-Bioengineering: Ecological Restoration with Native Plant and Seed Material Conference, Liezen, Austria, 5–9 September 2006. [Google Scholar]
- Elzenga, J.T.M.; Bekker, R.M.; Pritchard, H.W. Maximising the use of native seeds in restoration projects. Plant Biol. 2019, 21, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Landolt, E.; Baumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmle, R.W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.H.; Theurillat, J.-P.; et al. Flora Indicativa. Ecological Indicator Values and Biological Attributes of the Flora of Switzerland and the Alps; Haupt Verlag: Bern, Switzerland, 2010. [Google Scholar]
- Pignatti, E.; Pignatti, S. Plant. Life of the Dolomites. Vegetation Structure and Ecology; Springer Nature: Heidelberg/Berlin, Germany, 2014. [Google Scholar]
- Biondi, E.; Blasi, C. Prodromo Della Vegetazione d’Italia. Available online: http://www.prodromo-vegetazione-italia.org (accessed on 1 March 2020).
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italia, 2017. [Google Scholar]
- Giupponi, L.; Giorgi, A. Mount Cavallo Botanical Path: A proposal for the valorization of an area of the Orobie Bergamasche Regional Park (Southern Alps). J. Prot. Mt. Areas Res. Manag. 2017, 9, 5–15. [Google Scholar] [CrossRef]
- Martiónez-Dióaz, E.; Martiónez-Saónchez, J.J.; Conesa, E.; Franco, J.A.; Vicente, M.J. Germination and morpho-phenological traits of Allium melananthum, a rare species from south-eastern Spain. Flora 2018, 249, 16–23. [Google Scholar] [CrossRef]
- Cerabolini, B.; De Andreis, R.; Ceriani, R.M.; Pierce, S.; Raimondi, B. (2004) Seed germination and conservation of endangered species from the Italian Alps: Physoplexis comosa and Primula glaucescens. Biol. Conserv. 2004, 117, 351–356. [Google Scholar] [CrossRef]
- Yang, J.; Lovett-Doust, J.; Lovett-Doust, L. Seed germination patterns in green dragon (Arisaema dracontium, Araceae). Am. J. Bot. 1999, 86, 1160–1167. [Google Scholar] [CrossRef]
- Lozano-Isla, F.; Benites-Alfaro, O.; Pompelli, M.F. GerminaR: An R package for germination analysis with the interactive web application “GerminaQuant for R”. Ecol. Res. 2018, 34, 339–346. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Wien, Austria, 2018; Available online: http://www.r-project.org (accessed on 1 March 2020).
- Dixon, P. Vegan, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Del Egido, L.L.; Toorop, P.E.; Lanfermeijer, F.C. Seed enhancing treatments: Comparative analysis of germination characteristics of 23 key herbaceous species used in European restoration programmes. Plant Biol. 2019, 21, 398–408. [Google Scholar] [CrossRef]
- Vishal, B.; Kumar, P.P. Regulation of Seed Germination and Abiotic Stresses by Gibberellins and Abscisic Acid. Front. Plant Sci. 2018, 9, 838. [Google Scholar] [CrossRef]
- Pedrini, S.; Lewandrowski, W.; Stevens, J.C.; Dixon, K.W. Optimising seed processing techniques to improve germination and sowability of native grasses for ecological restoration. Plant Biol. 2019, 21, 415–424. [Google Scholar] [CrossRef]
- Pierce, S.; Spada, A.; Caporali, E.; Ceriani, R.M.; Buffa, G. Enzymatic scarification of Anacamptis morio (Orchidaceae) seed facilitates lignin degradation, water uptake and germination. Plant Biol. 2019, 21, 409–414. [Google Scholar] [CrossRef]
- Schwienbacher, E.; Navarro-Cano, J.A.; Neuner, G.; Erschbamer, B. Correspondence of seed traits with niche position in glacier foreland succession. Plant Ecol. 2012, 213, 371–382. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life. Functional Plant Ecology of High Mountain Ecosystems; Springer Nature: Heidelberg/Berlin, Germany, 2003. [Google Scholar]
- Giupponi, L.; Giorgi, A. A contribution to the knowledge of Linaria tonzigii Lona, a steno-endemic species of the Orobie Bergamasche Regional Park (Italian Alps). J. Prot. Mt. Areas Res. Manag. 2019, 11, 16–24. [Google Scholar] [CrossRef]
- Walker, L.R.; del Moral, R. Primary Succession and Ecosystem Rehabilitation; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Walker, L.R.; del Moral, R. Lessons from primary succession for restoration of severely damaged habitats. Appl. Veg. Sci. 2008, 12, 56–67. [Google Scholar] [CrossRef]
- Andreis, C.; Caccianiga, M.; Cerabolini, B. Vegetation and environmental factors during primary succession on glacier forelands: Some outlines from the Italian Alps. Plant Biosyst. 2001, 135, 295–310. [Google Scholar] [CrossRef]
- Paiero, P.; Paiero, G. La Vegetazione Rivierasca Alpina. In Atti del 41° Corso di Cultura in Ecologia: Conoscere il Sistema Fiume Nell’ambiente Alpino; D’Agostino, V., Carraro, V., Eds.; Dipartimento TeSAF Università degli Studi di Padova: Padova, Italy, 2005; pp. 28–45. [Google Scholar]
- Caccianiga, M.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. The functional basis of a primary succession resolved by CSR classification. Oikos 2006, 112, 10–20. [Google Scholar] [CrossRef]
- Pierce, S.; Negreiros, D.; Cerabolini, B.E.L.; Kattge, J.; Díaz, S.; Kleyer, M.; Shipley, B.; Wright, S.J.; Soudzilovskaia, N.A.; Onipchenko, V.G.; et al. A global method for calculating plant CSR ecological strategies applied across biomes world-wide. Funct. Ecol. 2017, 31, 444–457. [Google Scholar] [CrossRef]
- Eichel, J. Vegetation Succession and Biogeomorphic Interactions in Glacier Forelands. In Geomorphology of Proglacial Systems. Geography of the Physical Environment; Heckmann, T., Morche, D., Eds.; Springer Nature: Zürich, Switzerland, 2019; pp. 327–349. [Google Scholar]
- Schröter, C. Das Pflanzenleben der Alpen; Verlag von Albert Raustein: Zürich, Switzerland, 1926. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Non-Forest Vegetation: Coastal to Alpine, Natural to Man-MadeHabitats; Springer International Publishing: Zürich, Switzerland, 2017. [Google Scholar]
- Eichel, J.; Corenblit, D.; Dikau, R. (2016) Conditions for feedbacks between geomorphic and vegetation dynamics on lateral moraine slopes: A biogeomorphic feedback window. Earth Surf. Proc. Land. 2016, 41, 406–419. [Google Scholar] [CrossRef]
- Reisigl, H.; Keller, R. Alpenpflanzen im Lebensraum; Spektrum Akademischer Verlag: Heidelberg, Germany, 1994. [Google Scholar]
- Jones, C.G. Ecosystem engineers and geomorphological signatures in landscapes. Geomorphology 2012, 157–158, 75–87. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachack, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Boldrin, D.; Leung, A.K.; Bengough, A.G. Root biomechanical properties during establishment of woody perennials. Ecol. Eng. 2010, 109, 196–206. [Google Scholar] [CrossRef]
- Vergani, C.; Chiaradia, E.A.; Bischetti, G.B. Variability in the tensile resistance of roots in Alpine forest tree species. Ecol. Eng. 2012, 46, 43–56. [Google Scholar] [CrossRef]
- Pal, A.K.; Ahmed, A.; Panday, V.K. Soil Binding Capacity of Different Forage Grasses in Terms of Root. Indian J. Hill Farming 2019, 32, 137–143. [Google Scholar]
- Giadrossich, F.; Schwarz, M.; Cohen, D.; Cislaghi, A.; Vergani, C.; Hubble, T.; Phillips, C.; Stokes, A. Methods to measure the mechanical behaviour of tree roots: A review. Ecol. Eng. 2017, 109, 256–271. [Google Scholar] [CrossRef]
- De Vitis, M.; Mondoni, A.; Pritchard, H.W.; Laverack, G.; Bonomi, C. Native Seed Ecology, Production and Policy–Advancing Knowledge and Technology in Europe; MUSE: Trento, Italy, 2018. [Google Scholar]
- De Vitis, M.; Abbandonato, H.; Dixon, K.W.; Laverack, G.; Bonomi, C.; Pedrini, S. The European Native Seed Industry: Characterisation and Perspectives in Grassland Restoration. Sustainability 2017, 9, 1682. [Google Scholar] [CrossRef]
- Schmidt, I.B.; de Urzedo, D.I.; Pina-Rodrigues, F.C.M.; Vieira, D.L.M.; de Rezende, G.M.; Sampaio, A.B.; Junqueira, R.G.P. Community-based native seed production for restoration in Brazil–the role of science and policy. Plant Biol. 2019, 21, 389–397. [Google Scholar] [CrossRef]
- Nevill, P.G.; Tomlinson, S.; Elliott, C.P.; Espeland, E.K.; Dixon, K.W.; Merritt, D.J. Seed production areas for the global restoration challenge. Ecol. Evol. 2016, 6, 7490–7497. [Google Scholar] [CrossRef]
- Durka, W.; Michalski, S.G.; Berendzen, K.W.; Bossdorf, O.; Bucharova, A.; Hermann, J.; Hölzel, N.; Kollmann, J. Genetic differentiation within multiple common grassland plants supports seed transfer zones for ecological restoration. J. Appl. Ecol. 2017, 54, 116–126. [Google Scholar] [CrossRef]
- Prasse, R.; Kunzmann, D.; Schröder, R. Entwicklung und Praktische Umsetzung Naturschutzfachlicher Mindestanforderungen an Einen Herkunftsnachweis fuör Gebietseigenes Wildpflanzensaatgut Krautiger Pflanzen; Institute for Environmental Planning of the Gottfried Wilhelm Leibniz Universitaöt: Hannover, Germany, 2010. [Google Scholar]
- REWISA. Pruöfrichtlinie fuör die Gewinnung und den Vertrieb von Regionalen Wildgraösern und Wildkraöutern (REWISA®). Lehr- und Forschungszentrum fuör Landwirtschaft; Raumberg-Gumpenstein: Irdning, Austria, 2010. [Google Scholar]
- SKEW. Empfehlungen fuör den Anbau und Die Verwendung von Pflanz–Und Saatgut Einheimischer Wildpflanzen. Sekretariat; SKEW: Nyon, Switzerland, 2009. [Google Scholar]
- Giupponi, L.; Corti, C.; Manfredi, P. Onopordum acanthium subsp. acanthium in una ex-discarica della Pianura Padana (Piacenza). Ital. Bot. 2013, 45, 213–219. [Google Scholar]
- Giupponi, L.; Corti, C.; Manfredi, P. The vegetation of the Borgotrebbia landfill (Piacenza, Italy): Phytosociological and ecological characteristics. Plant Biosyst. 2015, 149, 865–874. [Google Scholar] [CrossRef]
- Giupponi, L.; Corti, C.; Manfredi, P.; Cassinari, C. Application of the floristic-vegetational indexes system for the evaluation of the environmental quality of a semi-natural area of the Po Valley (Piacenza, Italy). Plant Sociol. 2013, 50, 47–56. [Google Scholar] [CrossRef]
- Merrit, D.J.; Dixon, K.W. Restoration seed banks-A matter of scale. Science 2011, 332, 424–425. [Google Scholar] [CrossRef]
- Pagliacci, F.; Pavone, P.; Russo, M.; Giorgi, A. Regional structural heterogeneity: Evidence and policy implications for RIS3 in macro-regional strategies. Reg. Stud. 2019, 54, 765–775. [Google Scholar] [CrossRef]
- Pérez, F.L. Phytogeomorphic Influence of Stone Covers and Boulders on Plant Distribution and Slope Processes in High-Mountain Areas. Geogr. Compass 2009, 3, 1774–1803. [Google Scholar] [CrossRef]
- Cammeraat, E.; van Beek, R.; Kooijman, A. Vegetation succession and its consequences for slope stability in SE Spain. Plant Soil 2005, 278, 135–147. [Google Scholar] [CrossRef]
- Greenwood, J.R.; Norris, J.E.; Wint, J. Site investigation for the effects of vegetation on ground stability. J. Geotech. Geol. Eng. 2006, 24, 467–481. [Google Scholar] [CrossRef]
- Giupponi, L.; Pentimalli, D.; Manzo, A.; Panseri, S.; Giorgi, A. Effectiveness of fine root fingerprinting as a tool to identify plants of the Alps: Results of a preliminary study. Plant Biosyst. 2018, 152, 464–473. [Google Scholar] [CrossRef]
- Isselin-Nondedeu, F.; Bédécarrats, A. Influence of alpine plants growing on steep slopes on sediment trapping and transport by runoff. Catena 2007, 71, 330–339. [Google Scholar] [CrossRef]
- Baroni, C.; Armiraglio, S.; Gentili, R.; Carton, A. Landform-vegetation units for investigating the dynamics and geomorphologic evolution of alpine composite debris cones (Valle dell’Avio, Adamello Group, Italy). Geomorphology 2007, 84, 59–79. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Response to Treatment | Sum of Squares | Mean of Squares | F3,9 | p | |
|---|---|---|---|---|---|---|
| Adenostyles alpina | GRP | 1619.0 | 539.7 | 5.044 | 0.017 | * |
| GSP | 2.680 | 0.893 | 2.509 | 0.108 | ||
| Dryas octopetala | GRP | 1011.0 | 337.0 | 1.220 | 0.345 | |
| GSP | 2.933 | 0.978 | 4.302 | 0.028 | * | |
| Papaver aurantiacum | GRP | 20,499.0 | 6833.0 | 135.8 | <0.001 | * |
| GSP | 5.910 | 1.970 | 0.380 | 0.770 | ||
| Pulsatilla alpina | GRP | 433.0 | 147.7 | 4.567 | 0.024 | * |
| GSP | 9.565 | 3.188 | 41.55 | <0.001 | * | |
| Rumex scutatus | GRP | 4891.0 | 1630.3 | 43.28 | <0.001 | * |
| GSP | 687.5 | 229.2 | 37.50 | <0.001 | * | |
| Tofieldia calyculata | GRP | 13,043.0 | 4348.0 | 169.40 | <0.001 | * |
| GSP | 4.269 | 1.423 | 10.28 | 0.009 | * | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giupponi, L.; Leoni, V. Alpine Pioneer Plants in Soil Bioengineering for Slope Stabilization and Restoration: Results of a Preliminary Analysis of Seed Germination and Future Perspectives. Sustainability 2020, 12, 7190. https://doi.org/10.3390/su12177190
Giupponi L, Leoni V. Alpine Pioneer Plants in Soil Bioengineering for Slope Stabilization and Restoration: Results of a Preliminary Analysis of Seed Germination and Future Perspectives. Sustainability. 2020; 12(17):7190. https://doi.org/10.3390/su12177190
Chicago/Turabian StyleGiupponi, Luca, and Valeria Leoni. 2020. "Alpine Pioneer Plants in Soil Bioengineering for Slope Stabilization and Restoration: Results of a Preliminary Analysis of Seed Germination and Future Perspectives" Sustainability 12, no. 17: 7190. https://doi.org/10.3390/su12177190
APA StyleGiupponi, L., & Leoni, V. (2020). Alpine Pioneer Plants in Soil Bioengineering for Slope Stabilization and Restoration: Results of a Preliminary Analysis of Seed Germination and Future Perspectives. Sustainability, 12(17), 7190. https://doi.org/10.3390/su12177190