Stability of Ecosystem CO2 Flux in Response to Changes in Precipitation in a Semiarid Grassland


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Design
2.2. Measurement of Ecosystem CO2 Flux and Soil Temperature and Moisture
2.3. Data Analysis
3. Results
3.1. Changes in Microclimate
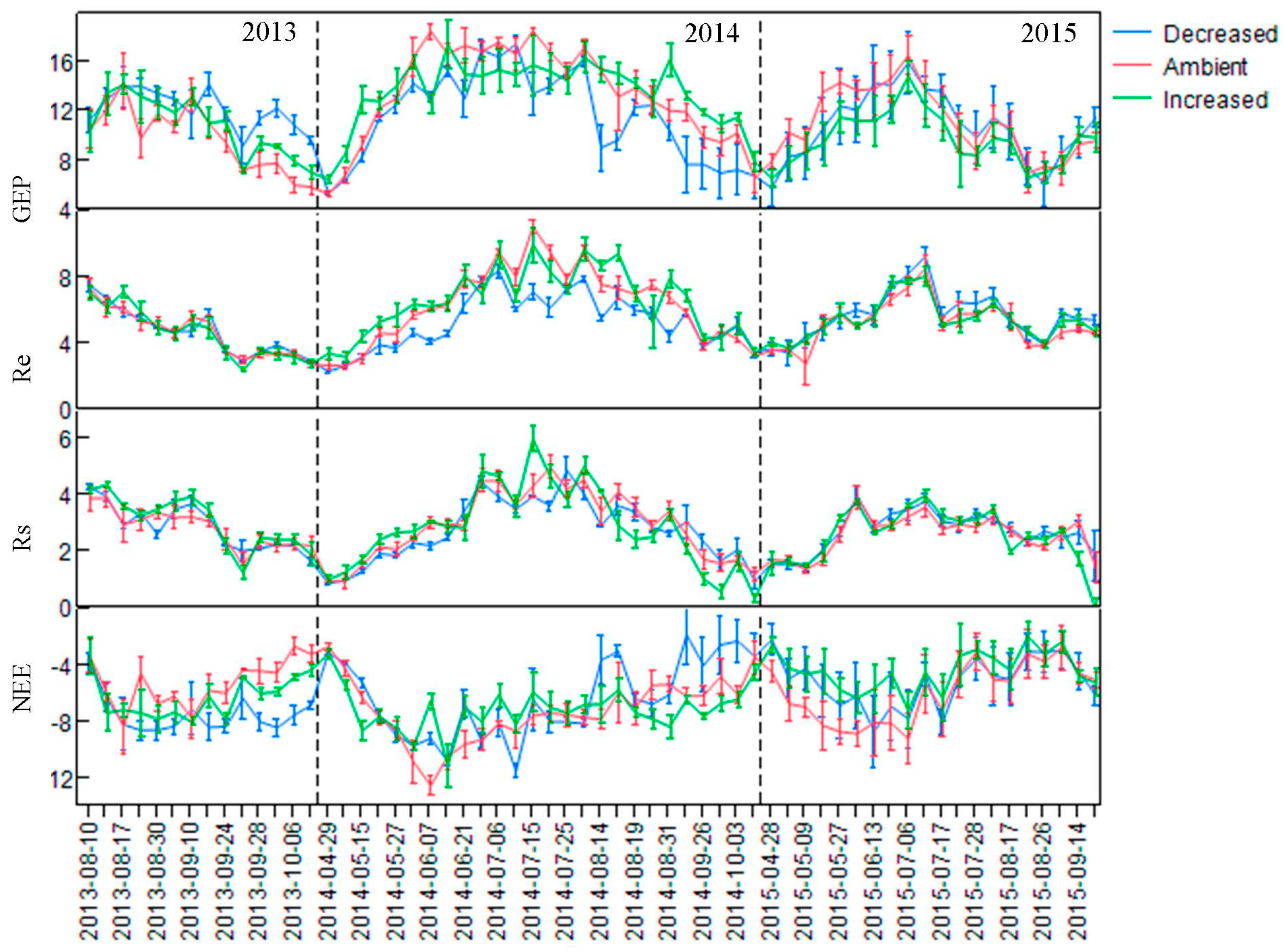
3.2. Seasonal and Diurnal Changes in Ecosystem CO2 Flux
3.3. Dependence of Ecosystem CO2 Flux Components on Soil Temperature and Moisture
4. Discussion
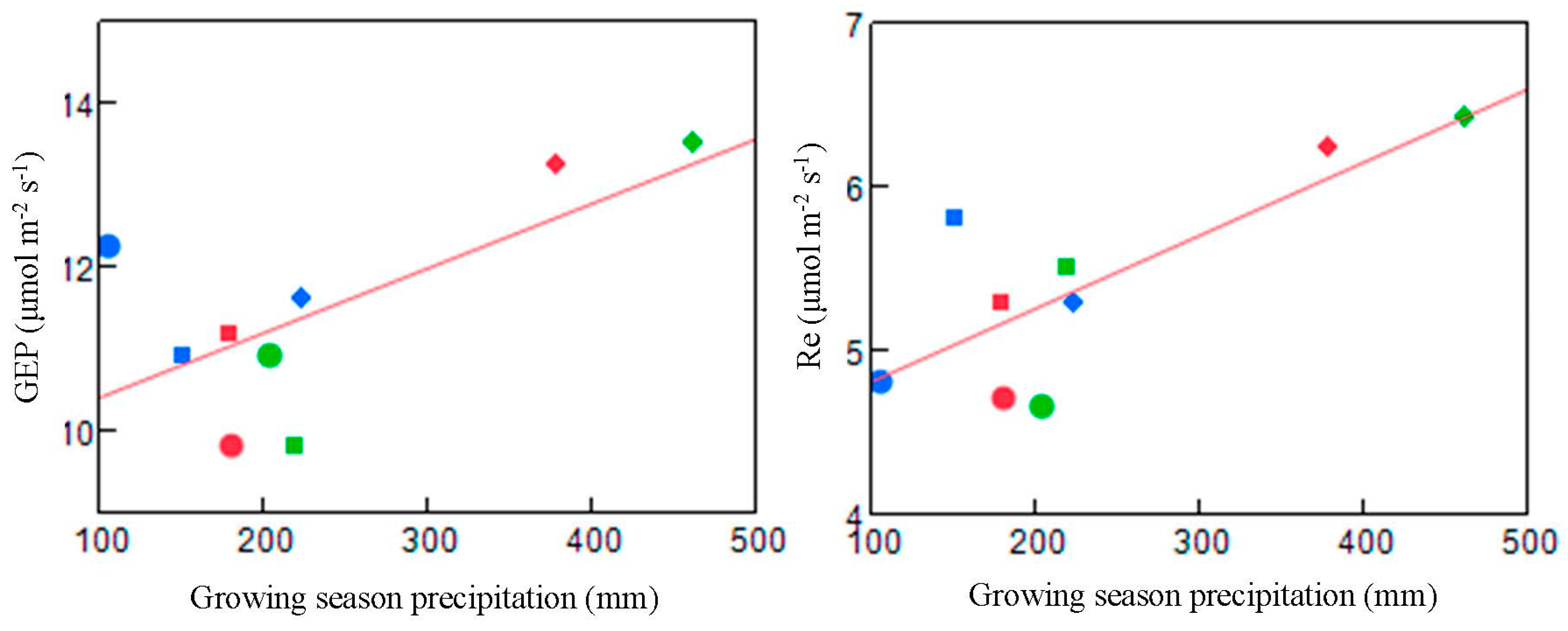
4.1. Response of CO2 Flux to the Precipitation Treatments
4.2. Dependence of CO2 Flux on Soil Temperature
4.3. Dependence of CO2 Flux on Soil Moisture
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carbone, M.S.; Still, C.J.; Ambrose, A.R.; Dawson, T.E.; Williams, A.P.; Boot, C.M.; Schaeffer, S.M.; Schimel, J.P. Seasonal and episodic moisture controls on plant and microbial contributions to soil respiration. Oecologia 2011, 167, 265–278. [Google Scholar] [CrossRef]
- Morgan, J.A.; LeCain, D.R.; Pendall, E.; Blumenthal, D.M.; Kimball, B.A.; Carrilli, Y.; Williams, D.G.; Heisler-White, J.; Dijkstra, F.A.; West, M. C4 grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland. Nature 2011, 476, 202–205. [Google Scholar] [CrossRef]
- Poulter, B.; Frank, D.; Ciais, P.; Myneni, R.B.; Andela, N.; Bi, J.; Broquet, G.; Canadell, J.G.; Chevallier, F.; Liu, Y.Y.; et al. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 2014, 509, 600–603. [Google Scholar] [CrossRef]
- Austin, A.T.; Yahdjian, L.; Stark, J.M.; Belnap, J.; Porporato, A.; Norton, U.; Ravetta, D.A.; Schaeffer, S.M. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 2004, 141, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfeld, J.G.; Ravit, B.; Elgersma, K. Feedback in the plant-soil system. Annu. Rev. Environ. Resour. 2005, 30, 75–115. [Google Scholar] [CrossRef]
- Rodríguez-Iturbe, I.; Porporato, A. Ecohydrology of Water-Controlled Ecosystems: Soil Moisture and Plant Dynamics; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Hovenden, M.J.; Newton, P.C.; Wills, K.E. Seasonal not annual rainfall determines grassland biomass response to carbon dioxide. Nature 2014, 511, 583–586. [Google Scholar] [CrossRef]
- Reich, P.B.; Hobbie, S.E.; Lee, T.D. Plant growth enhancement by elevated CO2 eliminated by joint water and nitrogen limitation. Nat. Geosci. 2014, 7, 920–924. [Google Scholar] [CrossRef]
- Knapp, A.K.; Fay, P.A.; Blair, J.M.; Collins, S.L.; Smith, M.D.; Carlisle, J.D.; Harper, C.W.; Danner, B.T.; Lett, M.S.; McCarron, J.K. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 2002, 298, 2202–2205. [Google Scholar] [CrossRef]
- Doughty, C.E.; Metcalfe, D.; Girardin, C.; Amézquita, F.F.; Cabrera, D.G.; Huasco, W.H.; Silva-Espejo, J.E.; Araujo-Murakami, A.; da Costa, M.C.; Rocha, W.; et al. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 2015, 519, 78–82. [Google Scholar] [CrossRef]
- Lázaro, R.; Rodrigo, F.; Gutiérrez, L.; Domingo, F.; Puigdefábregas, J. Analysis of a 30-year rainfall record (1967–1997) in semi–arid SE Spain for implications on vegetation. J. Arid Environ. 2001, 48, 373–395. [Google Scholar] [CrossRef]
- Modarres, R.; da Silva, V.d.P.R. Rainfall trends in arid and semi-arid regions of Iran. J. Arid Environ. 2007, 70, 344–355. [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Sala, O.E. Long-term forage production of North American shortgrass steppe. Ecol. Appl. 1992, 2, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Sala, O.E.; Gherardi, L.A.; Reichmann, L.; Jobbagy, E.; Peters, D. Legacies of precipitation fluctuations on primary production: Theory and data synthesis. Philos. Trans. R. Soc. B 2012, 367, 3135–3144. [Google Scholar] [CrossRef]
- Oechel, W.C.; Vourlitis, G.L.; Hastings, S.J.; Zulueta, R.C.; Hinzman, L.; Kane, D. Acclimation of ecosystem CO2 exchange in the Alaskan Arctic in response to decadal climate warming. Nature 2000, 406, 978–981. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wan, S.; Hui, D.; Wallace, L.L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature 2001, 413, 622–625. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Sendall, K.M.; Stefanski, A.; Wei, X.; Rich, R.L.; Montgomery, R.A. Boreal and temperate trees show strong acclimation of respiration to warming. Nature 2016, 531, 633–636. [Google Scholar] [CrossRef]
- Atkin, O.K.; Tjoelker, M.G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci. 2003, 8, 343–351. [Google Scholar] [CrossRef]
- Shen, W.; Reynolds, J.F.; Hui, D. Responses of dryland soil respiration and soil carbon pool size to abrupt vs. gradual and individual vs. combined changes in soil temperature, precipitation, and atmospheric [CO2]: A simulation analysis. Glob. Chang. Biol. 2009, 15, 2274–2294. [Google Scholar] [CrossRef]
- Akiyama, T.; Kawamura, K. Grassland degradation in China: Methods of monitoring, management and restoration. Grassl. Sci. 2007, 53, 1–17. [Google Scholar] [CrossRef]
- Li, X.; Fang, X.; Wu, F.; Miao, Y. Pollen evidence from Baode of the northern Loess Plateau of China and strong East Asian summer monsoons during the Early Pliocene. Chin. Sci. Bull. 2011, 56, 64–69. [Google Scholar] [CrossRef]
- Chen, S.; Lin, G.; Huang, J.; Jenerette, G.D. Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe. Glob. Chang. Biol. 2009, 15, 2450–2461. [Google Scholar] [CrossRef]
- Jasoni, R.L.; Smith, S.D.; Arnone, J.A. Net ecosystem CO2 exchange in Mojave Desert shrublands during the eighth year of exposure to elevated CO2. Glob. Chang. Biol. 2005, 11, 749–756. [Google Scholar] [CrossRef]
- Sendall, K.M.; Reich, P.B.; Zhao, C.; Hou, J.; Wei, X.; Stefanski, A.; Rice, K.; Rich, R.L.; Montgomery, R.A. Acclimation of photosynthetic temperature optima of temperate and boreal tree species in response to experimental forest warming. Glob. Chang. Biol. 2015, 21, 1342–1357. [Google Scholar] [CrossRef] [PubMed]
- Inglima, I.; Alberti, A.; Bertolini, T.; Vaccari, F.P.; Giolo, B.; Miglietta, F.; Cotrufo, M.F.; Peressotti, A. Precipitation pulses enhance respiration of Mediterranean ecosystems: The balance between organic and inorganic components of increased soil CO2 efflux. Glob. Chang. Biol. 2009, 15, 1289–1301. [Google Scholar] [CrossRef]
- Qi, Y.; Xu, M. Separating the effects of moisture and temperature on soil CO2 efflux in a coniferous forest in the Sierra Nevada mountains. Plant Soil 2001, 237, 15–23. [Google Scholar] [CrossRef]
- Yuste, J.C.; Janssens, I.; Carrara, A.; Meiresonne, L.; Ceulemans, R. Interactive effects of temperature and precipitation on soil respiration in a temperate maritime pine forest. Tree Physiol. 2003, 23, 1263–1270. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, J.; Wan, H.; Zhou, M.; Zhao, L.; Li, W.; Shi, H. The plant growth patterns in the natural grassland of Yunwu Mountain. Acta Ecol. Sin. 2008, 28, 3168–3175. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Wever, L.A.; Carlson, P.J. Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland. Glob. Chang. Biol. 2002, 8, 599–615. [Google Scholar] [CrossRef]
- Huxman, T.E.; Cable, J.M.; Ignace, D.D.; Eilts, J.A.; English, N.B.; Weltzin, J.; Williams, D.G. Response of net ecosystem gas exchange to a simulated precipitation pulse in a semi-arid grassland: The role of native versus non-native grasses and soil texture. Oecologia 2004, 141, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Verburg, P.S.; Arnone, J.A., III; Obrist, D.; Schorran, D.E.; Evans, R.D.; Leroux-swarthout, D.; Johnson, D.W.; Luo, Y.; Coleman, J.S. Net ecosystem carbon exchange in two experimental grassland ecosystems. Glob. Chang. Biol. 2004, 10, 498–508. [Google Scholar] [CrossRef]
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.S.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef]
- Tang, J.; Baldocchi, D.D.; Xu, L. Tree photosynthesis modulates soil respiration on a diurnal time scale. Glob. Chang. Biol. 2005, 11, 1298–1304. [Google Scholar] [CrossRef]
- Suyker, A.E.; Verma, S.B. Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie. Glob. Chang. Biol. 2001, 7, 279–289. [Google Scholar] [CrossRef]
- Xu, L.; Baldocchi, D.D. Seasonal variation in carbon dioxide exchange over a Mediterranean annual grassland in California. Agric. Forest Meteorol. 2004, 123, 79–96. [Google Scholar] [CrossRef]
- Hastings, S.J.; Oechel, W.C.; Muhlia-Melo, A. Diurnal, seasonal and annual variation in the net ecosystem CO2 exchange of a desert shrub community (Sarcocaulescent) in Baja California, Mexico. Glob. Chang. Biol. 2005, 11, 927–939. [Google Scholar] [CrossRef]
- Wohlfahrt, G.; Hammerle, A.; Haslwanter, A.; Bahn, M.; Tappeiner, U.; Cernusca, A. Seasonal and inter-annual variability of the net ecosystem CO2 exchange of a temperate mountain grassland: Effects of weather and management. J. Geophys. Res. Atmos. 2008, 113. [Google Scholar] [CrossRef]
- Scott, R.L.; Jenerette, G.D.; Potts, D.L.; Huxman, T.E. Effects of seasonal drought on net carbon dioxide exchange from a woody-plant-encroached semiarid grassland. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Altaf Arain, M.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef]
- Heisler-White, J.L.; Knapp, A.K.; Kelly, E.F. Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia 2008, 158, 129–140. [Google Scholar] [CrossRef]
- Niu, S.; Yang, H.; Zhang, Z.; Wu, M.; Lu, Q.; Li, L.; Han, X.; Wan, S. Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe. Ecosystems 2009, 12, 915–926. [Google Scholar] [CrossRef]
- Jia, X.; Shao, M.; Wei, X. Soil CO2 efflux in response to the addition of water and fertilizer in temperate semiarid grassland in northern China. Plant Soil 2013, 373, 125–141. [Google Scholar] [CrossRef]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; Ogée, J.; Allard, V.; et al. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Gilgen, A.; Buchmann, N. Response of temperate grasslands at different altitudes to simulated summer drought differed but scaled with annual precipitation. Biogeosciences 2009, 6, 2525–2539. [Google Scholar] [CrossRef]
- Shao, M.; Yang, W.; Li, Y. The availability of soil water to plant in Loess region. J. Hydrol. Eng. 1987, 18, 38–44. [Google Scholar]
- Liu, N.N.; Zhao, S.W.; Yong, Y.H.; Wang, H.J.; Zhao, Y.G.; Ji, X.Y.; Chao, L.H. Study on water-holding capacity of the top soil of a steppe reserve in the Yunwu Mountains, Guyuan, Ningxia Hui Autonomous Region. Acta Agrestia Sin. 2006, 14, 338–342. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef]
- Ripley, B.S.; Gilbert, M.E.; Ibrahim, D.G.; Osborne, C.P. Drought constraints on C4 photosynthesis: Stomatal and metabolic limitations in C3 and C4 subspecies of Alloteropsis semialata. J. Exp. Bot. 2007, 58, 1351–1363. [Google Scholar] [CrossRef]
- Wan, S.; Xia, J.; Liu, W.; Niu, S. Photosynthetic overcompensation under nocturnal warming enhances grassland carbon sequestration. Ecology 2009, 90, 2700–2710. [Google Scholar] [CrossRef]
- Jump, A.S.; Penuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A.; Luo, Y. On the variability of respiration in terrestrial ecosystems: Moving beyond Q10. Glob. Chang. Biol. 2006, 12, 154–164. [Google Scholar] [CrossRef]
- Ayub, G.; Smith, R.A.; Tissue, D.T.; Atkin, O.K. Impacts of drought on leaf respiration in darkness and light in Eucalyptus saligna exposed to industrial-age atmospheric CO2 and growth temperature. New Phytol. 2011, 190, 1003–1018. [Google Scholar] [CrossRef] [PubMed]
- Galmés, J.; Ribas-Carbó, M.; Medrano, H.; Flexas, J. Response of leaf respiration to water stress in Mediterranean species with different growth forms. J. Arid Environ. 2007, 68, 206–222. [Google Scholar] [CrossRef]
- Almagro, M.; López, J.; Querejeta, J.; Martínez-Mena, M. Temperature dependence of soil CO2 efflux is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biol. Biochem. 2009, 41, 594–605. [Google Scholar] [CrossRef]
- Jia, X.; Shao, M.; Wei, X. Responses of soil respiration to N addition, burning and clipping in temperate semiarid grassland in northern China. Agric. Forest Meteorol. 2012, 166, 32–40. [Google Scholar] [CrossRef]
- Schwalm, C.R.; Williams, C.A.; Schaefer, K.; Arneth, A.; Bonal, D.; Buchmann, N.; Chen, J.; Law, B.E.; Lindroth, A.; Luyssaert, S.; et al. Assimilation exceeds respiration sensitivity to drought: A FLUXNET synthesis. Glob. Chang. Biol. 2010, 16, 657–670. [Google Scholar] [CrossRef]
- Wan, S.; Hui, D.; Wallace, L.; Luo, Y. Direct and indirect effects of experimental warming on ecosystem carbon processes in a tallgrass prairie. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Oberbauer, S.F.; Tweedie, C.E.; Welker, J.M.; Fahnestock, J.T.; Henry, G.H.R.; Webber, P.J.; Hollister, R.D.; Walker, M.D.; Kuchy, A.; Elmore, E.; et al. Tundra CO2 fluxes in response to experimental warming across latitudinal and moisture gradients. Ecol. Monogr. 2007, 77, 221–238. [Google Scholar] [CrossRef]
- Yvon-Durocher, G.; Jones, J.I.; Trimmer, M.; Woodward, G.; Montoya, J.M. Warming alters the metabolic balance of ecosystems. Philos. Trans. R. Soc. B 2010, 365, 2117–2126. [Google Scholar] [CrossRef]
- Jia, X.; Shao, M.; Wei, X.; Li, X. Response of soil CO2 efflux to water addition in temperate semiarid grassland in northern China: The importance of water availability and species composition. Biol. Fertil. Soils 2014, 50, 839–850. [Google Scholar] [CrossRef]
- Janssens, I.A.; Pilegaard, K. Large seasonal changes in Q10 of soil respiration in a beech forest. Glob. Chang. Biol. 2003, 9, 911–918. [Google Scholar] [CrossRef]
- Reichstein, M.; Subke, J.A.; Angeli, A.C.; Tenhunen, J.D. Does the temperature sensitivity of decomposition of soil organic matter depend upon water content, soil horizon, or incubation time? Glob. Chang. Biol. 2005, 11, 1754–1767. [Google Scholar] [CrossRef]
- Wan, S.; Norby, R.J.; Ledford, J.; Weltzin, J.F. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Glob. Chang. Biol. 2007, 13, 2411–2424. [Google Scholar] [CrossRef]
- Limousin, J.; Bickford, C.P.; Dickman, L.T.; Pangle, R.E.; Hudson, P.J.; Boutz, A.L.; Gehres, N.; Osuna, J.L.; Pockman, W.T.; McDowell, N.G. Regulation and acclimation of leaf gas exchange in a piñon–juniper woodland exposed to three different precipitation regimes. Plant Cell Environ. 2013, 36, 1812–1825. [Google Scholar] [CrossRef]
- Sanaullah, M.; Chabbi, A.; Rumpel, C.; Kuzyakov, Y. Carbon allocation in grassland communities under drought stress followed by 14C pulse labeling. Soil Biol. Biochem. 2012, 55, 132–139. [Google Scholar] [CrossRef]
- Sanaullah, M.; Blagodatskaya, E.; Chabbi, A.; Rumpel, C.; Kuzyakov, Y. Drought effects on microbial biomass and enzyme activities in the rhizosphere of grasses depend on plant community composition. Appl. Soil Ecol. 2011, 48, 38–44. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Van Dijk, A.I.; De Jeu, R.A.; Canadell, J.G.; McCabe, M.F.; Evans, J.P.; Wang, G. Recent reversal in loss of global terrestrial biomass. Nat. Clim. Chang. 2015, 5, 470–474. [Google Scholar] [CrossRef]
- Smith, N.G.; Dukes, J.S. Plant respiration and photosynthesis in global-scale models: Incorporating acclimation to temperature and CO2. Glob. Chang. Biol. 2013, 19, 45–63. [Google Scholar] [CrossRef]
- Bagley, J.; Rosenthal, D.M.; Ruiz-Vera, U.M.; Siebers, M.H.; Kumar, P.; Ort, D.R.; Bernacchi, C.J. The influence of photosynthetic acclimation to rising CO2 and warmer temperatures on leaf and canopy photosynthesis models. Glob. Biogeochem. Cycles 2015, 29, 194–206. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEP | Re | Rs | NEE | ||
|---|---|---|---|---|---|
| SS | Stage of season | 1321.0 | 741.2 | 286.9 | 332.7 |
| Treatment | 6.2 | 12.5 | 0.1 | 7.6 | |
| Interaction | 120.5 | 19.8 | 6.0 | 114.3 | |
| Date | 346.9 | 5.6 | 26.9 | 440.4 | |
| F | Stage of season | 51.3 | 153.3 | 175.4 | 21.9 |
| Treatment | 0.2 | 2.6 | 0.0 | 0.5 | |
| Interaction | 2.3 | 2.1 | 1.8 | 3.8 | |
| Date | 27.0 | 2.3 | 32.9 | 58.1 | |
| P | Stage of season | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Treatment | 0.785 | 0.0755 | 0.9674 | 0.6047 | |
| Interaction | 0.0538 | 0.0857 | 0.1187 | 0.0048 | |
| Date | <0.0001 | 0.1292 | <0.0001 | <0.0001 |
| a | SE(a) | b | SE(b) | c | SE(c) | T0 (°C) | RMSE | R2 | P | |
| Across treatments | −0.065 | 0.006 | 2.419 | 0.198 | −8.190 | 1.478 | 18.61 | 2.943 | 0.386 | <0.01 |
| Ambient precipitation | −0.078 a | 0.014 | 2.861 a | 0.398 | −11.555 a | 2.782 | 18.34 | 3.003 | 0.476 | <0.01 |
| Decreased precipitation | −0.043 b | 0.010 | 1.684 b | 0.325 | −3.321 b | 2.571 | 19.58 | 3.152 | 0.250 | <0.01 |
| Increased precipitation | −0.088 a | 0.016 | 3.060 a | 0.458 | −11.862 a | 3.175 | 17.39 | 2.488 | 0.452 | <0.01 |
| a | SE(a) | b | SE(b) | Q10 | SE(Q10) | RMSE | R2 | P | ||
| Across treatments | 2.575 | 0.134 | 0.052 | 0.003 | 1.682 | 0.050 | 1.548 | 0.429 | <0.01 | |
| Ambient precipitation | 2.293 ab | 0.208 | 0.064 a | 0.005 | 1.896 a | 0.095 | 1.516 | 0.517 | <0.01 | |
| Decreased precipitation | 2.401 a | 0.158 | 0.046 b | 0.004 | 1.584 b | 0.063 | 1.048 | 0.566 | <0.01 | |
| Increased precipitation | 2.024 b | 0.194 | 0.073 a | 0.006 | 2.075 a | 0.125 | 1.437 | 0.554 | <0.01 | |
| a | SE(a) | b | SE(b) | Q10 | SE(Q10) | RMSE | R2 | P | ||
| Across treatments | 1.242 | 0.078 | 0.055 | 0.004 | 1.733 | 0.069 | 0.961 | 0.364 | <0.01 | |
| Ambient precipitation | 1.098 a | 0.113 | 0.067 a | 0.006 | 1.954 a | 0.117 | 0.867 | 0.472 | <0.01 | |
| Decreased precipitation | 1.204 a | 0.105 | 0.050 b | 0.005 | 1.649 b | 0.082 | 0.740 | 0.486 | <0.01 | |
| Increased precipitation | 1.017 a | 0.151 | 0.072 a | 0.009 | 2.054 a | 0.185 | 1.100 | 0.330 | <0.01 | |
| a | SE(a) | b | SE(b) | c | SE(c) | T0 (°C) | RMSE | R2 | P | |
| Across treatments | 0.045 | 0.005 | −1.469 | 0.161 | 4.443 | 1.199 | 16.32 | 2.388 | 0.175 | <0.01 |
| Ambient precipitation | 0.058 a | 0.011 | −1.889 a | 0.309 | 7.725 a | 2.160 | 16.28 | 2.331 | 0.254 | <0.01 |
| Decreased precipitation | 0.036 b | 0.009 | −1.207 b | 0.300 | 2.671 b | 2.370 | 16.76 | 2.905 | 0.122 | <0.01 |
| Increased precipitation | 0.070 a | 0.012 | −2.077 a | 0.341 | 7.647 a | 2.363 | 14.84 | 1.851 | 0.215 | <0.01 |
| a | SE(a) | b | SE(b) | RMSE | R2 | P | |
| Across treatments | 15.151 | 0.574 | −0.175 | 0.032 | 3.616 | 0.070 | <0.01 |
| Ambient precipitation | 17.845 a | 1.236 | −0.313 ab | 0.066 | 3.843 | 0.137 | <0.01 |
| Decreased precipitation | 15.769 b | 0.800 | −0.341 a | 0.057 | 3.186 | 0.227 | <0.01 |
| Increased precipitation | 18.124 a | 1.129 | −0.267 b | 0.055 | 3.095 | 0.145 | <0.01 |
| a | SE(a) | b | SE(b) | RMSE | R2 | P | |
| Across treatments | 6.293 | 0.323 | −0.043 | 0.018 | 2.033 | 0.014 | 0.02 |
| Ambient precipitation | 7.976 a | 0.675 | −0.121 a | 0.036 | 2.100 | 0.074 | <0.01 |
| Decreased precipitation | 5.528 b | 0.395 | −0.048 b | 0.028 | 1.572 | 0.023 | 0.09 |
| Increased precipitation | 8.973 a | 0.737 | −0.154 a | 0.036 | 2.021 | 0.117 | <0.01 |
| a | SE(a) | b | SE(b) | RMSE | R2 | P | |
| Across treatments | 3.497 | 0.188 | −0.039 | 0.010 | 1.185 | 0.033 | <0.01 |
| Ambient precipitation | 3.863 b | 0.375 | −0.052 b | 0.020 | 1.166 | 0.045 | 0.01 |
| Decreased precipitation | 3.084 c | 0.255 | −0.035 b | 0.018 | 1.017 | 0.029 | 0.06 |
| Increased precipitation | 5.200 a | 0.448 | −0.114 a | 0.022 | 1.227 | 0.165 | <0.01 |
| a | SE(a) | b | SE(b) | RMSE | R2 | P | |
| Across treatments | −8.858 | 0.399 | 0.132 | 0.022 | 2.516 | 0.081 | <0.01 |
| Ambient precipitation | −9.869 a | 0.811 | 0.192 b | 0.044 | 2.522 | 0.121 | <0.01 |
| Decreased precipitation | −10.240 a | 0.680 | 0.293 a | 0.049 | 2.707 | 0.231 | <0.01 |
| Increased precipitation | −9.151 a | 0.733 | 0.113 c | 0.036 | 2.011 | 0.067 | <0.01 |
| GEP | Re | Rs | NEE | ||
|---|---|---|---|---|---|
| SS | Moisture | 10 | 67 | 5 | 25 |
| Temperature | 856 | 982 | 246 | 4 | |
| Interaction | 863 | 126 | 46 | 330 | |
| F | Moisture | 1 | 34 | 6 | 3 |
| Temperature | 68 | 504 | 309 | 1 | |
| Interaction | 69 | 65 | 57 | 41 | |
| P | Moisture | 0.3647 | <0.0001 | 0.0124 | 0.0808 |
| Temperature | <0.0001 | <0.0001 | <0.0001 | 0.4658 | |
| Interaction | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, K.; Tian, H.; Su, M.; Qiu, L.; Wei, X.; Zhang, Y.; Liu, J.; Gao, H.; Cheng, J. Stability of Ecosystem CO2 Flux in Response to Changes in Precipitation in a Semiarid Grassland. Sustainability 2019, 11, 2597. https://doi.org/10.3390/su11092597
Bao K, Tian H, Su M, Qiu L, Wei X, Zhang Y, Liu J, Gao H, Cheng J. Stability of Ecosystem CO2 Flux in Response to Changes in Precipitation in a Semiarid Grassland. Sustainability. 2019; 11(9):2597. https://doi.org/10.3390/su11092597
Chicago/Turabian StyleBao, Kaiqiang, Haifeng Tian, Min Su, Liping Qiu, Xiaorong Wei, Yanjiang Zhang, Jian Liu, Hailong Gao, and Jimin Cheng. 2019. "Stability of Ecosystem CO2 Flux in Response to Changes in Precipitation in a Semiarid Grassland" Sustainability 11, no. 9: 2597. https://doi.org/10.3390/su11092597
APA StyleBao, K., Tian, H., Su, M., Qiu, L., Wei, X., Zhang, Y., Liu, J., Gao, H., & Cheng, J. (2019). Stability of Ecosystem CO2 Flux in Response to Changes in Precipitation in a Semiarid Grassland. Sustainability, 11(9), 2597. https://doi.org/10.3390/su11092597