Contrasting Effects of Tillage and Landscape Structure on Spiders and Springtails in Vineyards

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
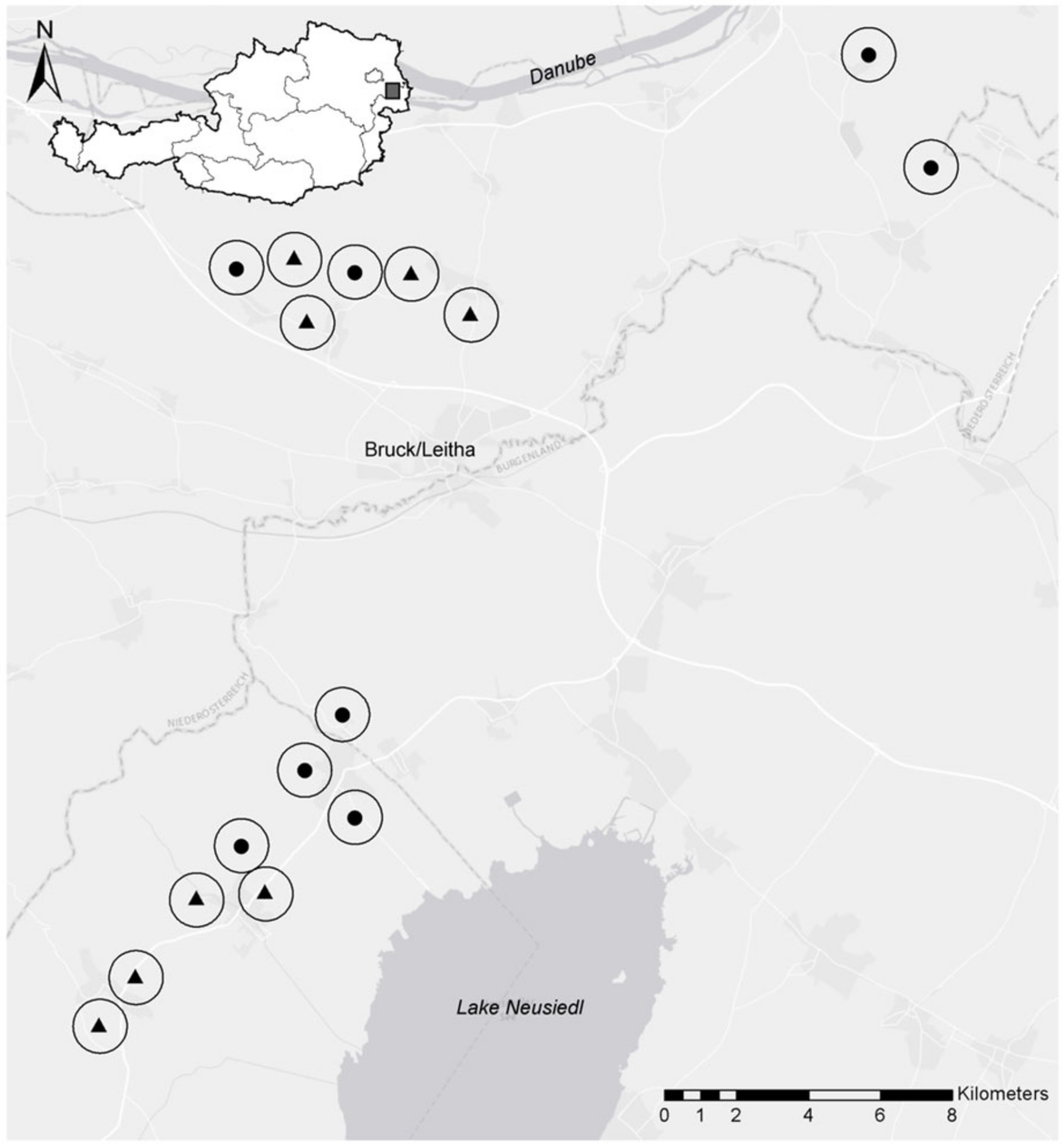
2.1. Site Description and Study Design
2.2. Sampling and Measurements
2.2.1. Spider and Springtail Sampling
2.2.2. Landscape Parameters
2.3. Statistical Analysis
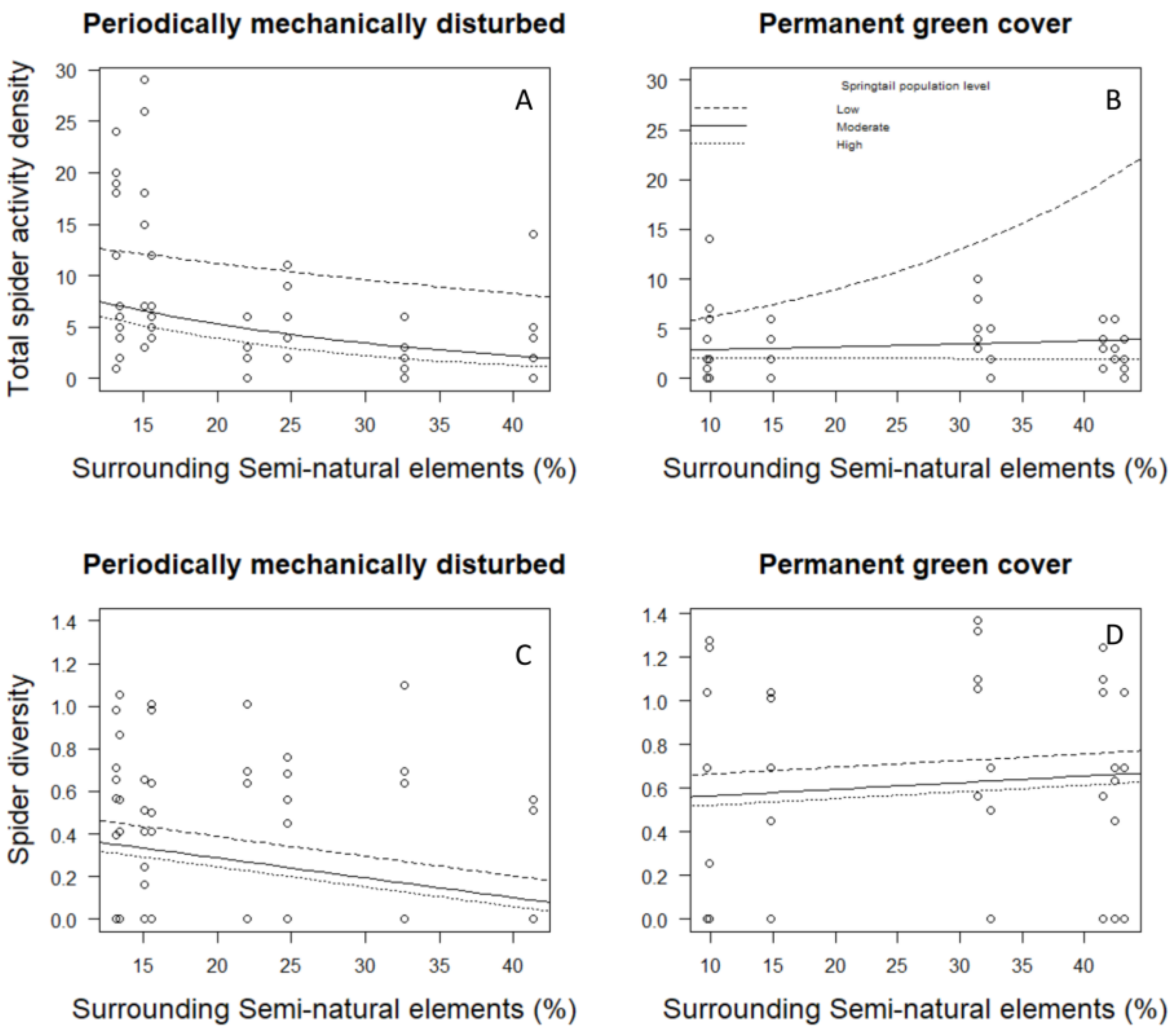
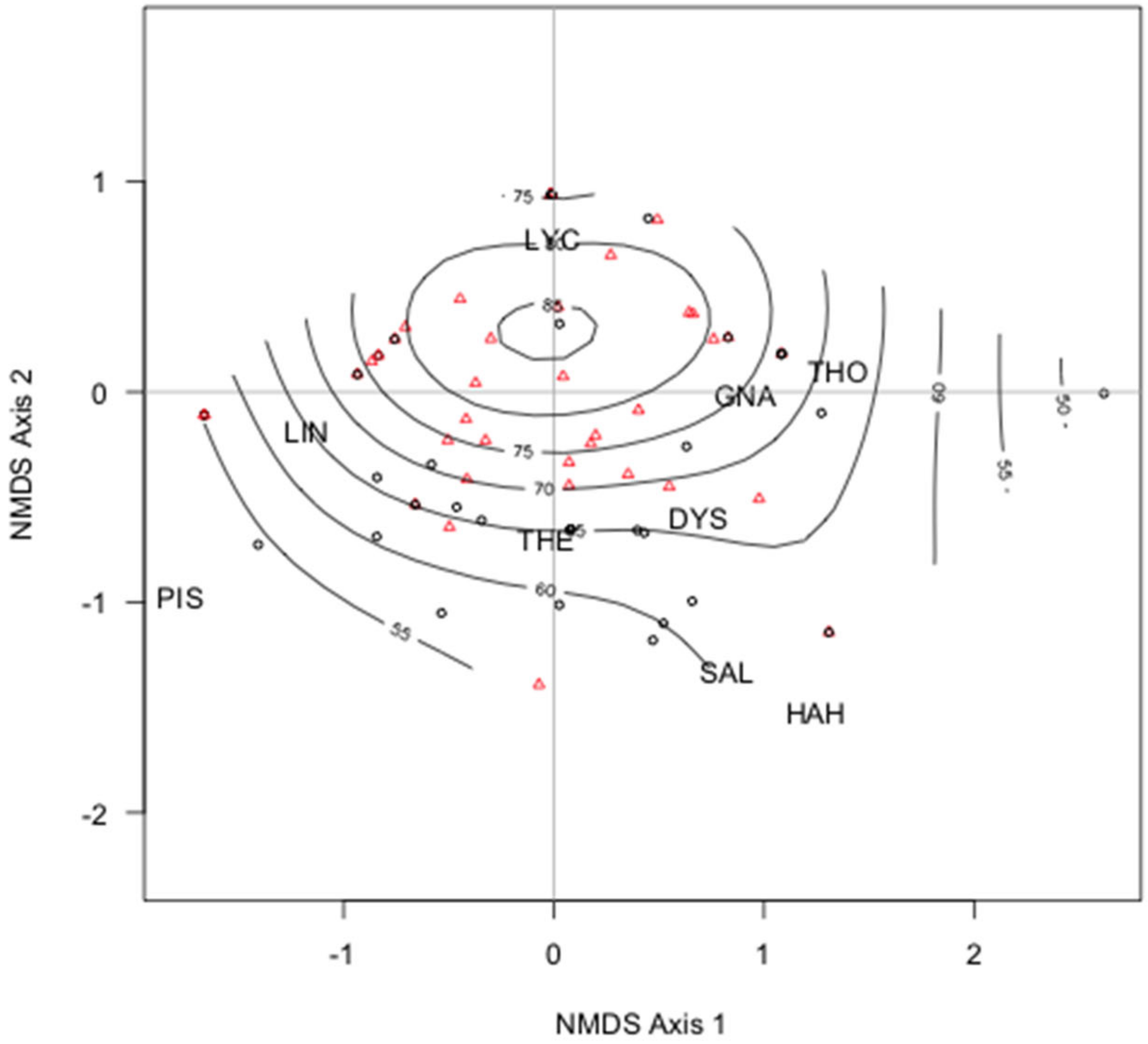
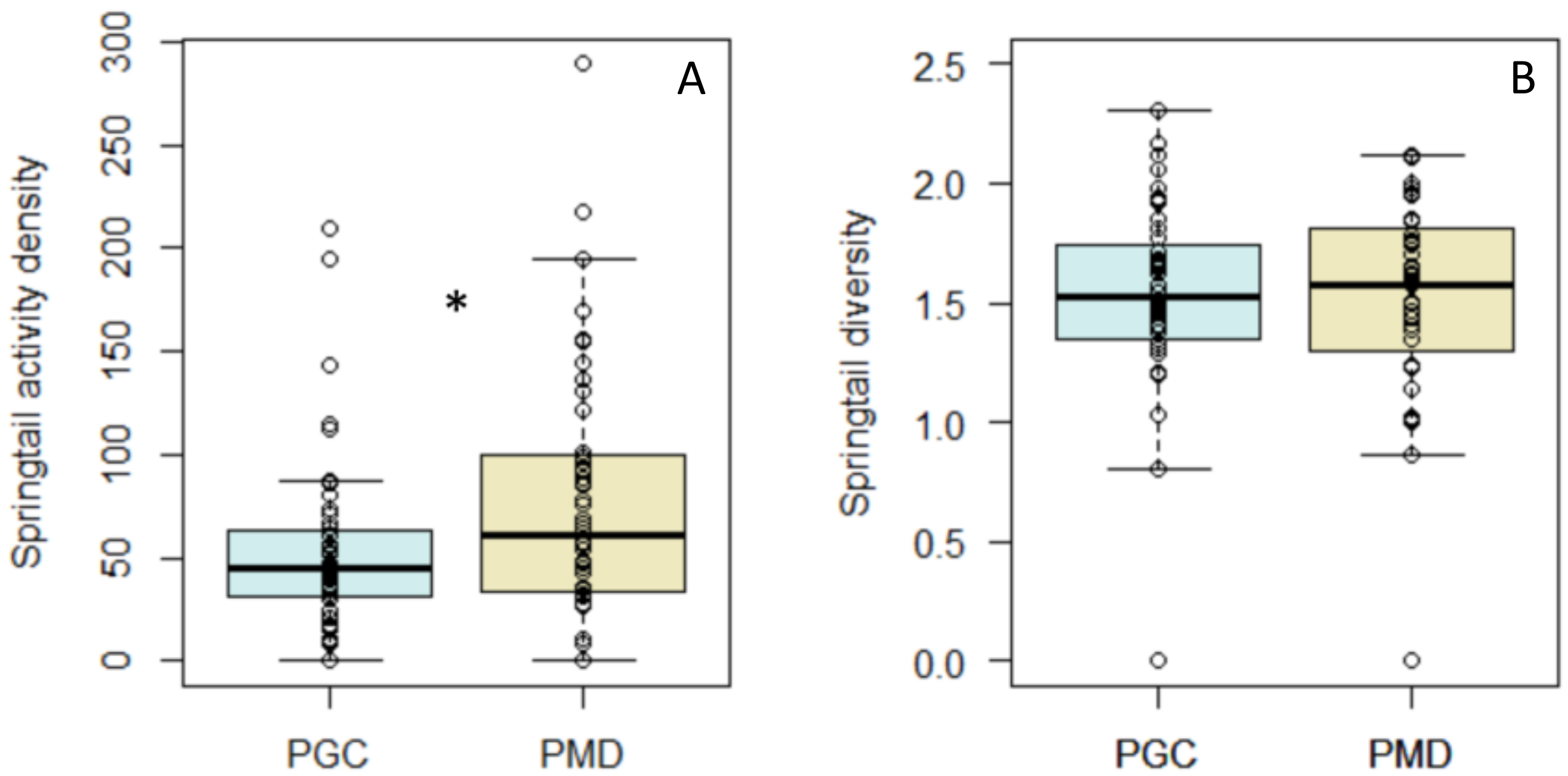
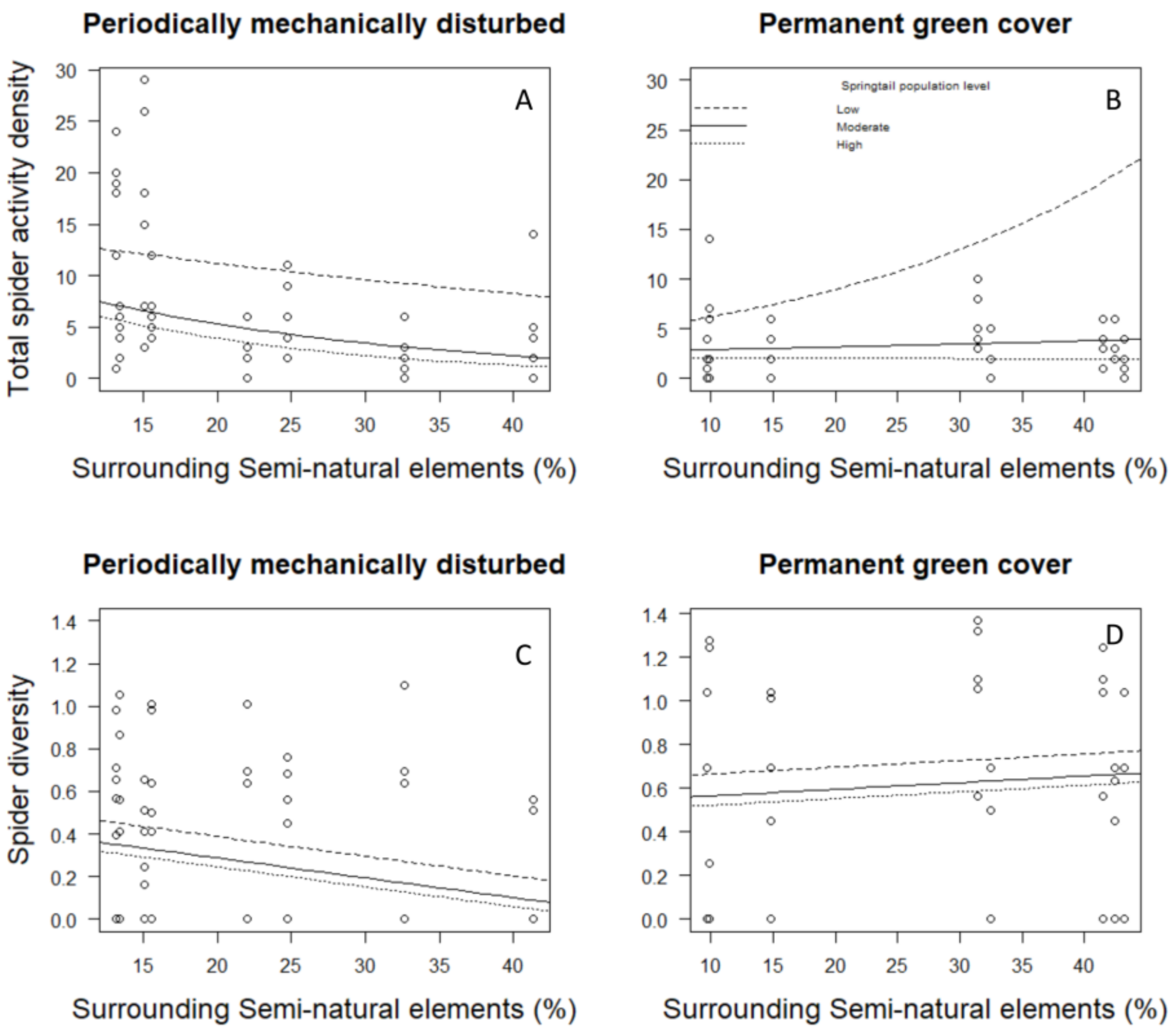
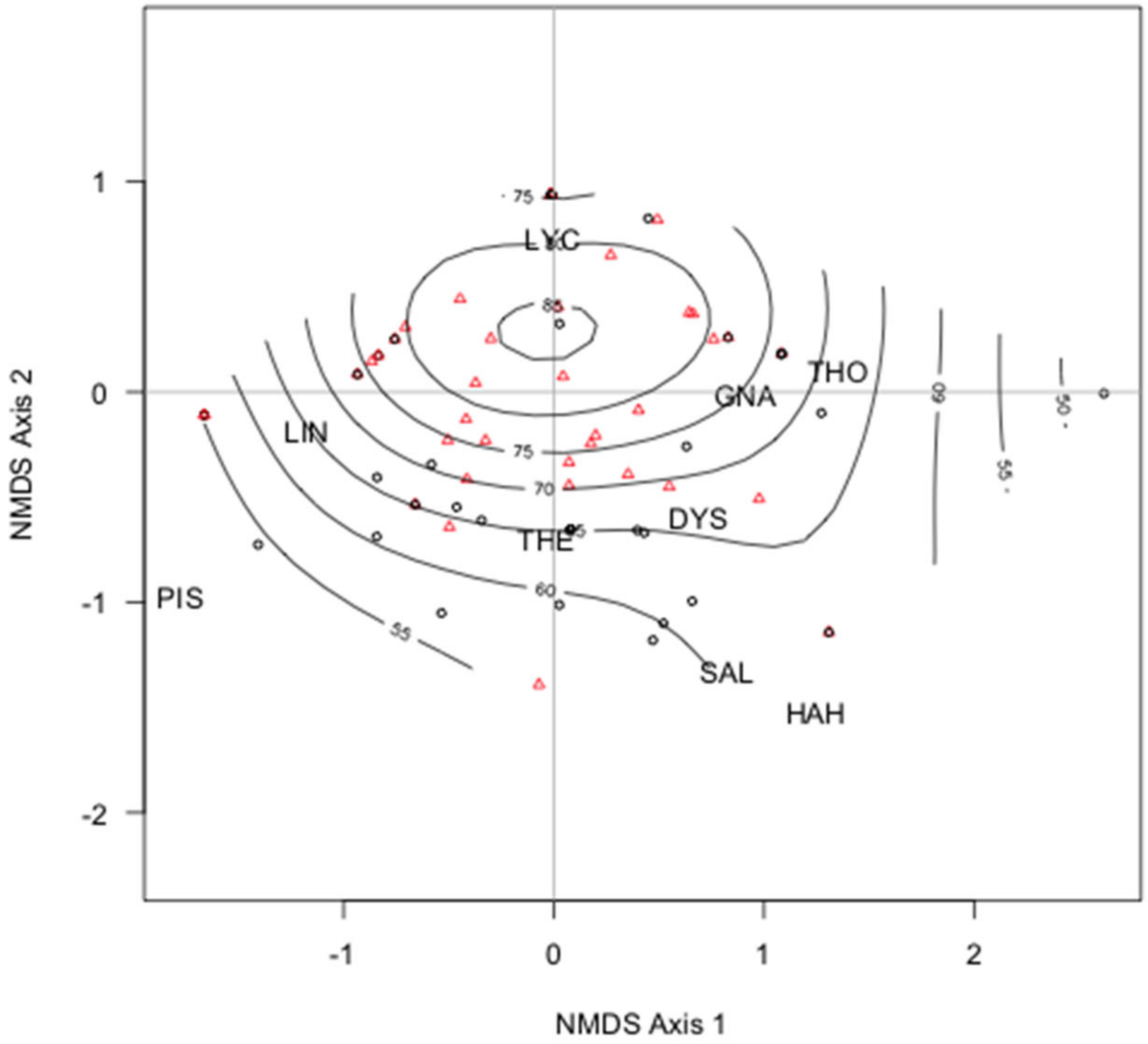
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spider Family | Inter-Row Management | |
|---|---|---|
| PMD | PGC | |
| Lycosidae | 34.5 ± 6.2 | 9.5 ± 2.2 |
| Theridiidae | 4.1 ± 1.0 | 3.3 ± 0.9 |
| Pisauridae | 0.0 ± 0.0 | 0.1 ± 0.2 |
| Gnaposidae | 1.9 ± 0.6 | 1.8 ± 0.5 |
| Linyphiidae | 2.5 ± 0.7 | 2.1 ± 0.7 |
| Hahniidae | 0.1 ± 0.1 | 0.4 ± 0.3 |
| Thomisidae | 1.1 ± 0.9 | 1.3 ± 0.6 |
| Saltiicidae | 0.0 ± 0.0 | 0.1 ± 0.2 |
| Dysderidae | 0.1 ± 0.1 | 0.0 ± 0.0 |
| Springtail Species | Inter-Row Management | |
|---|---|---|
| PMD | PGC | |
| Ceratophysella succinea (Gisin 1949) | 0.6 ± 3.7 | 1.1 ± 2.1 |
| Cyphoderus albinus (Nicolet 1842) | 0.2 ± 0.8 | 0.2 ± 0.9 |
| Cyphoderus bidenticulatus (Parona 1888) | 0.1 ± 0.2 | 0 |
| Desoria fennica cf. (Reuter 1895) | 0 | 0.1 ± 0.2 |
| Desoria olivacea (Tullberg 1871) | 4.7 ± 8.5 | 1.7 ± 2.2 |
| Entomobrya handschini (Stach 1922) | 1.8 ± 1.9 | 4.8 ± 4.5 |
| Entomobrya marginata (Tullberg 1871) | 2.7 ± 6.1 | 0.5 ± 1.1 |
| Entomobrya multifasciata (Tullberg 1871) | 0 | 0.1 ± 0.2 |
| Heteromurus major (Moniez 1889) | 17.4 ± 17.3 | 13.2 ± 15.9 |
| Heteromurus nitidus (Templeton 1835) | 1.0 ± 1.2 | 0.9 ± 2.0 |
| Heterosminthurus claviger (Gisin 1958) | 0.1 ± 0.3 | 0 |
| Isotoma viridis (Bourlet 1839) | 29.0 ± 25.0 | 17.7 ± 25.3 |
| Lepidocyrtus cyaneus (Tullberg 1871) | 13.2 ± 14.6 | 2.2 ± 2.9 |
| Lepidocyrtus lanuginosus (Gmelin 1788) | 0.7 ± 2.0 | 0.1 ± 0.6 |
| Lepidocyrtus lignorum (Fabricius 1775) | 0 | 0.2 ± 0.7 |
| Lepidocyrtus paradoxus (Uzel 1891) | 1.1 ± 2.0 | 1.5 ± 2.7 |
| Metaphorura affinis (Börner 1902) | 0 | 0.1 ± 0.2 |
| Orchesella cincta (Linnaeus 1758) | 5.8 ± 7.8 | 5.4 ± 6.5 |
| Orchesella pannonica (Stach 1960) | 0.1 ± 0.2 | 0.1 ± 0.2 |
| Parisotoma notabilis (Schäffer 1896) | 1.2 ± 2.7 | 2.3 ± 6.5 |
| Pogonognathellus flavescens (Tullberg 1871) | 0.1 ± 0.3 | 0.1 ± 0.5 |
| Pseudachorutella asigillata (Börner 1901) | 0 | 0.2 ± 0.6 |
| Pseudachorutes parvulus (Börner 1901) | 0.1 ± 0.2 | 0.6 ± 1.9 |
| Pseudosinella alba (Packard 1873) | 0.8 ± 2.4 | 0.4 ± 1.2 |
| Pseudosinella petterseni (Börner 1901) | 0.1 ± 0.2 | 0 |
| Pseudosinella sexoculata (Schött 1902) | 0.3 ± 0.7 | 0.1 ± 0.3 |
| Seira spp. | 0 | 0.1 ± 0.2 |
| Sminthurinus aureus (Lubbock 1862) | 4.0 ± 4.1 | 1.5 ± 1.8 |
| Sminthurus multipunctatus (Schäffer 1896) | 0.1 ± 0.4 | 0.1 ± 0.2 |
| Sminthurus viridis (Linnaeus 1758) | 0.1 ± 0.2 | 0 |
| Sphaeridia pumilis (Krausbauer 1898) | 0.7 ± 0.9 | 0.7 ± 1.2 |
| Tomocerus vulgaris (Tullberg 1871) | 0 | 0.1 ± 0.2 |
| Willowsia nigromaculata (Lubbock 1873) | 0.1 ± 0.2 | 0 |
| Xenylla maritima (Tullberg 1869) | 0 | 0.1 ± 0.2 |
References
- McGovern, P.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R. Wine Science: Principles and Applications, 4th ed.; Academic Press: London, UK, 2014; 978p. [Google Scholar]
- Zaller, J.G.; Cantelmo, C.; Santos, G.D.; Muther, S.; Gruber, E.; Pallua, P.; Mandl, K.; Friedrich, B.; Hofstetter, I.; Schmuckenschlager, B.; et al. Herbicides in vineyards reduce grapevine root mycorrhization and alter soil microorganisms and the nutrient composition in grapevine roots, leaves, xylem sap and grape juice. Environ. Sci. Pollut. Res. 2018, 25, 23215–23226. [Google Scholar] [CrossRef] [PubMed]
- Kratschmer, S.; Pachinger, B.; Schwantzer, M.; Paredes, D.; Guernion, M.; Burel, F.; Nicolai, A.; Strauss, P.; Bauer, T.; Kriechbaum, M.; et al. Tillage intensity or landscape features: What matters most for wild bee diversity in vineyards? Agric. Ecosyst. Environ. 2018, 266, 142–152. [Google Scholar] [CrossRef]
- Buchholz, J.; Querner, P.; Paredes, D.; Bauer, T.; Strauss, P.; Guernion, M.; Scimia, J.; Cluzeau, D.; Burel, F.; Kratschmer, S.; et al. Soil biota in vineyards are more influenced by plants and soil quality than by tillage intensity or the surrounding landscape. Sci. Rep. 2017, 7, 17445. [Google Scholar] [CrossRef] [PubMed]
- Faber, F.; Wachter, E.; Zaller, J.G. Earthworms are little affected by reduced soil tillage methods in vineyards. Plant Soil Environ. 2017, 63, 257–263. [Google Scholar]
- Winter, S.; Bauer, T.; Strauss, P.; Kratschmer, S.; Paredes, D.; Popescu, D.; Landa, B.; Guzmán, G.; Gómez, J.A.; Guernion, M.; et al. Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. J. Appl. Ecol. 2018, 55, 2484–2495. [Google Scholar] [CrossRef]
- World Spider Catalog. World Spider Catalog Version 19.5. Natural History Museum Bern. Available online: https://wsc.nmbe.ch (accessed on 3 July 2017).
- Wise, D.H. Spiders in Ecological Webs; University Press: Cambridge, UK, 1993. [Google Scholar]
- Foelix, R.F. Biology of Spiders, 3rd ed.; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Menta, C.; Leoni, A.; Gardi, C.; Conti, F.D. Are grasslands important habitats for soil microarthropod conservation? Biodivers. Conserv. 2011, 20, 1073–1087. [Google Scholar] [CrossRef]
- Nyffeler, M.; Benz, G. Spiders in natural pest control: A review. J. Appl. Entomol. 1987, 103, 321–339. [Google Scholar] [CrossRef]
- Michalko, R.; Pekár, S.; Entling, M.H. An updated perspective on spiders as generalist predators in biological control. Oecologia 2018. [CrossRef] [PubMed]
- Nyffeler, M.; Birkhofer, K. An estimated 400–800 million tons of prey are annually killed by the global spider community. Sci. Nat. Naturwiss. 2017, 104, 30. [Google Scholar] [CrossRef]
- Wood, J.R.; Holdaway, R.J.; Orwin, K.H.; Morse, C.; Bonner, K.I.; Davis, C.; Bolstridge, N.; Dickie, I.A. No single driver of biodiversity: Divergent responses of multiple taxa across land use types. Ecosphere 2017, 8, 17. [Google Scholar] [CrossRef]
- Kuusk, A.K.; Ekbom, B. Lycosid spiders and alternative food: Feeding behavior and implications for biological control. Biol. Control 2010, 55, 20–26. [Google Scholar] [CrossRef]
- Drapela, T.; Moser, D.; Zaller, J.G.; Frank, T. Spider assemblages in winter oilseed rape affected by landscape and site factors. Ecography 2008, 31, 254–262. [Google Scholar] [CrossRef]
- Costello, M.J.; Daane, K.M. Spider and leafhopper (Erythroneura spp.) response to vineyard ground cover. Environ. Entomol. 2003, 32, 1085–1098. [Google Scholar]
- Nyffeler, M.; Benz, G. Prey and predator importance of microphantid spiders in winter wheat fields and hay meadows. J. Appl. Entomol. 1988, 105, 190–197. [Google Scholar] [CrossRef]
- Bruggisser, O.T.; Schmidt-Entling, M.H.; Bacher, S. Effects of vineyard management on biodiversity at three trophic levels. Biol. Conserv. 2010, 143, 1521–1528. [Google Scholar] [CrossRef]
- Hanna, R.; Zalom, F.G.; Roltsch, W.J. Relative impact of spider predation and cover crop on population dynamics of Erythroneura variabilis in a raisin grape vineyard. Entomol. Exp. Appl. 2003, 107, 177–191. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 3 July 2017).
- Salamon, J.-A.; Wissuwa, J.; Moder, K.; Frank, T. Effects of Medicago sativa, Taraxacum officinale and Bromus sterilis on the density and diversity of Collembola in grassy arable fallows of different ages. Pedobiologia 2011, 54, 63–70. [Google Scholar] [CrossRef]
- Hopkin, S.P. Biology of the Springtails (Insecta: Collembola); Oxford University Press: Oxford, NY, USA, 1997. [Google Scholar]
- Rusek, J. Biodiversity of Collembola and their functional role in the ecosystem. Biodivers. Conserv. 1998, 7, 1207–1219. [Google Scholar] [CrossRef]
- Roy, S.; Bano, R.; Saxena, P.; Bhatt, R.K. Land uses and its impact on community structure of soil collembola. Range Manag. Agrofor. 2014, 35, 27–31. [Google Scholar]
- Coulibaly, S.F.M.; Coudrain, V.; Hedde, M.; Brunet, N.; Mary, B.; Recous, S.; Chauvat, M. Effect of different crop management practices on soil Collembola assemblages: A 4-year follow-up. Appl. Soil Ecol. 2017, 119, 354–366. [Google Scholar] [CrossRef]
- Song, L.H.; Liu, J.; Yan, X.M.; Chang, L.; Wu, D.H. Euedaphic and hemiedaphic Collembola suffer larger damages than epedaphic species to nitrogen input. Environ. Pollut. 2016, 208, 413–415. [Google Scholar] [CrossRef]
- Zaller, J.G.; König, N.; Tiefenbacher, A.; Muraoka, Y.; Querner, P.; Ratzenböck, A.; Bonkowski, M.; Koller, R. Pesticide seed dressings can affect the activity of various soil organisms and reduce decomposition of plant material. BMC Ecol. 2016, 16, 37. [Google Scholar] [CrossRef]
- Agusti, N.; Shayler, S.P.; Harwood, J.D.; Vaughan, I.P.; Sunderland, K.D.; Symondson, W.O.C. Collembola as alternative prey sustaining spiders in arable ecosystems: Prey detection within predators using molecular markers. Mol. Ecol. 2003, 12, 3467–3475. [Google Scholar] [CrossRef]
- Fiera, C. Application of stable isotopes and lipid analysis to understand trophic interactions in springtails. North West J. Zool. 2014, 10, 227–235. [Google Scholar]
- Sunderland, K.D.; Fraser, A.M.; Dixon, A.F.G. Distribution of linyphiid spiders in relation to capture of prey in cereal fields. Pedobiologia 1986, 29, 367–375. [Google Scholar]
- Greenstone, M.H. Spider feeding behaviour optimizes dietary essential amino acid composition. Nature 1979, 282, 501–503. [Google Scholar] [CrossRef]
- Bilde, T.; Axelsen, J.A.; Toft, S. The value of Collembola from agricultural soils as food for a generalist predator. J. Appl. Ecol. 2000, 37, 672–683. [Google Scholar] [CrossRef]
- Costello, M.J.; Daane, K.M. Influence of ground cover on spider populations in a table grape vineyard. Ecol. Entomol. 1998, 23, 33–40. [Google Scholar] [CrossRef]
- Sharley, D.J.; Hoffman, A.A.; Thomson, L.J. The effect of soil tillage on beneficial invertebrates within the vineyard. Agric. For. Entomol. 2008, 10, 233–243. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Roschewitz, I.; Thies, C.; Tscharntke, T. Differential effects of landscape and management on diversity and density of ground-dwelling farmland spiders. J. Appl. Ecol. 2005, 42, 281–287. [Google Scholar] [CrossRef]
- Isaia, M.; Bona, F.; Badino, G. Influence of landscape diversity and agricultural practices on spider assemblage in Italian vineyards of Langa Astigiana (Northwest Italy). Environ. Entomol. 2006, 35, 297–307. [Google Scholar] [CrossRef]
- Hogg, B.N.; Daane, K.M. Diversity and invasion within a predator community: Impacts on herbivore suppression. J. Appl. Ecol. 2011, 48, 453–461. [Google Scholar] [CrossRef]
- Wilson, H.; Wong, J.S.; Thorp, R.W.; Miles, A.F.; Daane, K.M.; Altieri, M.A. Summer Flowering Cover Crops Support Wild Bees in Vineyards. Environ. Entomol. 2017, 47, 63–69. [Google Scholar] [CrossRef]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape perspectives on agricultural intensification and biodiversity—Ecosystem service management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Öberg, S.; Ekbom, B.; Bommarco, R. Influence of habitat type and surrounding landscape on spider diversity in Swedish agroecosystems. Agric. Ecosyst. Environ. 2007, 122, 211–219. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Thies, C.; Nentwig, W.; Tscharntke, T. Contrasting responses of arable spiders to the landscape matrix at different spatial scales. J. Biogeogr. 2008, 35, 157–166. [Google Scholar] [CrossRef]
- Gardiner, M.M.; Landis, D.A.; Gratton, C.; Schmidt, N.; O’Neal, M.; Mueller, E.; Chacon, J.; Heimpel, G.E. Landscape composition influences the activity density of Carabidae and Arachnida in soybean fields. Biol. Control 2010, 55, 11–19. [Google Scholar] [CrossRef]
- Ponge, J.F.; Dubs, F.; Gillet, S.; Sousa, J.P.; Lavelle, P. Decreased biodiversity in soil springtail communities: The importance of dispersal and landuse history in heterogeneous landscapes. Soil Biol. Biochem. 2006, 38, 1158–1161. [Google Scholar] [CrossRef]
- Querner, P.; Bruckner, A.; Drapela, T.; Moser, D.; Zaller, J.G.; Frank, T. Landscape and site effects on Collembola diversity and abundance in winter oilseed rape fields in eastern Austria. Agric. Ecosyst. Environ. 2013, 164, 145–154. [Google Scholar] [CrossRef]
- Zentralanstalt für Meteorologie und Geodynamik. Klimamittel Bruckneudorf. Available online: https://www.zamg.ac.at/ (accessed on 22 June 2017).
- Blume, H.-P.; Brümmer, G.W.; Horn, R.; Kandeler, E.; Kögler-Knabner, I.; Kretzschmar, R.; Stahr, K.; Wilke, B.-M. Lehrbuch der Bodenkunde, 16th ed.; Spektrum Akademischer Verlag: Heidelberg, Germany, 2010. [Google Scholar]
- Drapela, T.; Frank, T.; Heer, X.; Moser, D.; Zaller, J.G. Landscape structure affects activity density, body size and fecundity of Pardosa wolf spiders (Araneae, Lycosidae) in winter oilseed rape. Eur. J. Entomol. 2011, 108, 609–614. [Google Scholar] [CrossRef]
- Bährmann, R.; Müller, H.J. Bestimmung wirbelloser Tiere—Bildtafeln für Zoologische Bestimmungsübungen und Exkursionen; Spektrum Akademischer Verlag: Heidelberg, Germany, 2008. [Google Scholar]
- Bellmann, H. Der Kosmos Spinnenführer; Franckh-Kosmos Verlag: Heidelberg, Germany, 2016. [Google Scholar]
- Podgaiski, L.R.; Joner, F.; Lavorel, S.; Moretti, M.; Ibanez, S.; Mendoca, M.D.S.; Pillar, V.D. Spider trait assembly patterns and resilience under fire-induced vegetation change in South Brazilian grasslands. PLoS ONE 2013, 8, e60207. [Google Scholar] [CrossRef]
- Gisin, H. Collembolenfauna Europas; Museum d’Histoire Naturelle: Genéve, Switzerland, 1960. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of this Group of Insects. Tribe: Orchesellini; Polska Akademia Nauk: Kraków, Poland, 1960. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of this Group of Insects. Tribe: Entomobryini; Polska Akademia Nauk: Kraków, Poland, 1963. [Google Scholar]
- Bretfeld, G. Synopses on Palaearctic Collembola, Vol. 2 Symphypleona; Staatliches Museum für Naturkunde: Görliz, Germany, 1999. [Google Scholar]
- Potapov, M. Synopses on Palaearctic Collembola, Volume 3: Isotomidae; State Museum of the Natural History Museum of Gorlitz: Görliz, Germany, 2001. [Google Scholar]
- Thibaud, J.M.; Schulz, H.-J.; da Gama Assalino, M.M. Synopses on Palaearctic Collembola, Vol. 4. Hypogastruridae; Staatliches Museum für Naturkunde: Görliz, Germany, 2004. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark, Part II: Entomobryomorpha and Symphypleona; Brill: Leiden, The Netherlands, 2007. [Google Scholar]
- Hopkin, S.P. A Key to the Collembola (Springtails) of Britain and Ireland; Field Studies Council: Shrewsbury, UK, 2007. [Google Scholar]
- Dunger, W.; Schlitt, B. Synopses on Palaearctic Collembola, Volume 6, Part 1: Onychiuroidea: Tullbergiidae; State Museum of the Natural History Museum of Gorlitz: Görliz, Germany, 2011. [Google Scholar]
- European Commission. CORINE Land Cover—Contents; European Environment Agency: Copenhagen, Denmark, 1994; pp. 1–94. [Google Scholar]
- Davies, C.E.; Moss, D.; Hill, M.O. EUNIS Habitat Classification Revised; European Topic Centre on Nature Protection and Biodiversity: Paris, France, 2004. [Google Scholar]
- ESRI. ArcGIS Desktop: Release 10.2; Environmental Systems Research Institute: Redlands, CA, USA, 2013. [Google Scholar]
- Paredes, D.; Cayuela, L.; Campos, M. Synergistic effects of ground cover and adjacent vegetation on natural enemies of olive insect pests. Agric. Ecosyst. Environ. 2013, 173, 72–80. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 2, 133–142. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.J.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0-3. 2012. Available online: http://CRAN.R-project.org/package=vegan (accessed on 7 July 2017).
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.9.13. 2013. Available online: http://CRAN.R-project.org/package=MuMIn (accessed on 7 July 2017).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R—A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 7 July 2017).
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Ann. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Roger-Estrade, J.; Anger, C.; Bertrand, M.; Richard, G. Tillage and soil ecology: Partners for sustainable agriculture. Soil Till. Res. 2010, 111, 33–40. [Google Scholar] [CrossRef]
- Nentwig, W. Non-webbuilding spiders: Prey specialists or generalists? Oecologia 1986, 69, 571–576. [Google Scholar] [CrossRef]
- Zulka, K.P.; Milasowszky, N.; Lethmayer, C. Spider biodiversity potential of an ungrazed and grazed inland salt meadow in the National Park ‘Neusiedler See- Seewinkel’ (Austria): Implications for management (Arachnida: Araneae). Biodivers. Conserv. 1997, 6, 75–88. [Google Scholar] [CrossRef]
- Seung-Tae, K.; Uhm, K.-B.; Lee, J.-H.; Yun, J.-C.; Park, C.-G.; Jung, M.-P. Spiders in a grape vineyard in Korea. J. Asia Pac. Entomol. 2002, 5, 181–184. [Google Scholar] [CrossRef]
- Bolduc, E.; Buddle, C.M.; Bostanian, N.J.; Vincent, C. Ground-dwelling spider fauna (Araneae) of two vineyards in Southern Quebec. Environ. Entomol. 2005, 34, 635–645. [Google Scholar] [CrossRef]
- Green, J.A. Sampling method and time determines composition of spider collections. J. Arachnol. 1999, 27, 176–182. [Google Scholar]
- Thomas, L.J.; Hoffmann, A.A. Vegetation increases the abundance of natural enemies in vineyards. Biol. Control 2009, 49, 259–269. [Google Scholar]
- Bell, J.R.; Bohan, D.A.; Shaw, E.M.; Weyman, G.S. Ballooning dispersal using silk: World fauna, phylogenies, genetics and models. Bull. Entomol. Res. 2005, 95, 69–114. [Google Scholar] [CrossRef]
- D’Alberto, C.F.; Hoffmann, A.A.; Thomson, L.J. Limited benefits of non-crop vegetation on spiders in Australian vineyards: Regional or crop differences. Biol. Control 2012, 57, 541–552. [Google Scholar] [CrossRef]
- Toft, S.; Wise, D.H. Growth, development, and survival of a generalist predator fed single- and mixed-species diets of different quality. Oecologia 1999, 119, 191–197. [Google Scholar] [CrossRef]
- Greenstone, M. The numerical response to prey availability of Pardosa ramulosa (McCook) (Araneae: Lycosidae) and its relation- ship to the role of spiders in the balance of nature. Symp. Zool. Soc. Lond. 1978, 42, 183–193. [Google Scholar]
- Harwood, J.D.; Sunderland, K.D.; Symondson, W.O.C. Living where the food is: Web location by linyphiid spiders in relation to prey availability in winter wheat. J. Appl. Ecol. 2001, 38, 88–99. [Google Scholar] [CrossRef]
- Sereda, E.; Blick, T.; Dorow, W.H.O.; Wolters, V.; Birkhofer, K. Spatial distribution of spiders and epedaphic Collembola in an environmentally heterogeneous forest floor habitat. Pedobiologia 2012, 55, 241–245. [Google Scholar] [CrossRef]
| Landscape Effect | Prey Effect | Nomenclature | Total Spider Density | Total Spider Diversity | |
|---|---|---|---|---|---|
| in | Null | Null | No | 553.75 | 109.98 |
| Null | Coll effect | Col | 532.48 | 112.07 | |
| SNE | Null | SNE | 553.44 | 112.16 | |
| SNE | Coll effect | SNE + Coll | 532.41 | 113.84 | |
| Interaction (SNE & Coll effect) | SNE + Coll + (SNE × Coll) | 530.50 | 115.99 | ||
| Management models | Null | Null | Mgmt | 550.63 | 110.22 |
| SNE | Null | Mgmt + SNE | 551.29 | 112.37 | |
| Null | Coll effect | Mgmt + Coll | 531.09 | 111.58 | |
| SNE | Coll effect | Mgmt + SNE + Coll | 531.89 | 113.77 | |
| Interaction (Mgmt & SNE) | Mgmt + SNE + (Mgmt × SNE) | 551.53 | 111.61 | ||
| Interaction (Mgmt & Collembola effect) | Mgmt + Coll + (Mgmt × Coll) | 532.80 | 113.04 | ||
| SNE | Coll effect + Interaction (Mgmt & Coll effect) | Mgmt + SNE + Coll + (Mgmt × Coll) | 533.84 | 115.13 | |
| SNE + Interaction (Mgmt & SNE) | Coll effect | Mgmt + SNE + Coll + (Mgmt × SNE) | 530.69 | 112.77 | |
| Interaction (SNE & Coll effect) | Mgmt + SNE + Coll + (SNE × Coll) | 531.26 | 115.70 | ||
| Interaction (Mgmt & Collembola effect) + Interaction (Coll effect & SNE) | Mgmt + SNE + Coll + (Mgmt × Col) + (Coll × SNE) | 529.61 | 114.49 | ||
| Interaction (Mgmt & SNE) + Interaction (Mgmt & Coll effect) | Mgmt + SNE + Coll + (Mgmt × SNE) + (M × Coll) | 532.67 | 113.90 | ||
| Interaction (Mgmt & SNE) + Interaction (Coll effect & SNE) | Mgmt + SNE + Coll + (Mgmt × SNE) + (Coll × SNE) | 532.68 | 117.30 | ||
| Interaction (Mgmt & SNE) + Interaction (Mgmt & Coll effect) + Interaction (Coll effect & SNE) | Mgmt + SNE + Coll + (Mgmt × SNE) + (Mgmt × Coll) + (Coll × SNE) | 531.09 | 115.89 | ||
| R2m | 0.30 | ||||
| R2c | 0.76 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfingstmann, A.; Paredes, D.; Buchholz, J.; Querner, P.; Bauer, T.; Strauss, P.; Kratschmer, S.; Winter, S.; Zaller, J. Contrasting Effects of Tillage and Landscape Structure on Spiders and Springtails in Vineyards. Sustainability 2019, 11, 2095. https://doi.org/10.3390/su11072095
Pfingstmann A, Paredes D, Buchholz J, Querner P, Bauer T, Strauss P, Kratschmer S, Winter S, Zaller J. Contrasting Effects of Tillage and Landscape Structure on Spiders and Springtails in Vineyards. Sustainability. 2019; 11(7):2095. https://doi.org/10.3390/su11072095
Chicago/Turabian StylePfingstmann, Alexandra, Daniel Paredes, Jacob Buchholz, Pascal Querner, Thomas Bauer, Peter Strauss, Sophie Kratschmer, Silvia Winter, and Johann Zaller. 2019. "Contrasting Effects of Tillage and Landscape Structure on Spiders and Springtails in Vineyards" Sustainability 11, no. 7: 2095. https://doi.org/10.3390/su11072095
APA StylePfingstmann, A., Paredes, D., Buchholz, J., Querner, P., Bauer, T., Strauss, P., Kratschmer, S., Winter, S., & Zaller, J. (2019). Contrasting Effects of Tillage and Landscape Structure on Spiders and Springtails in Vineyards. Sustainability, 11(7), 2095. https://doi.org/10.3390/su11072095