Prioritizing Green Spaces for Biodiversity Conservation in Beijing Based on Habitat Network Connectivity


Abstract
:
1. Introduction
2. Materials and Methods
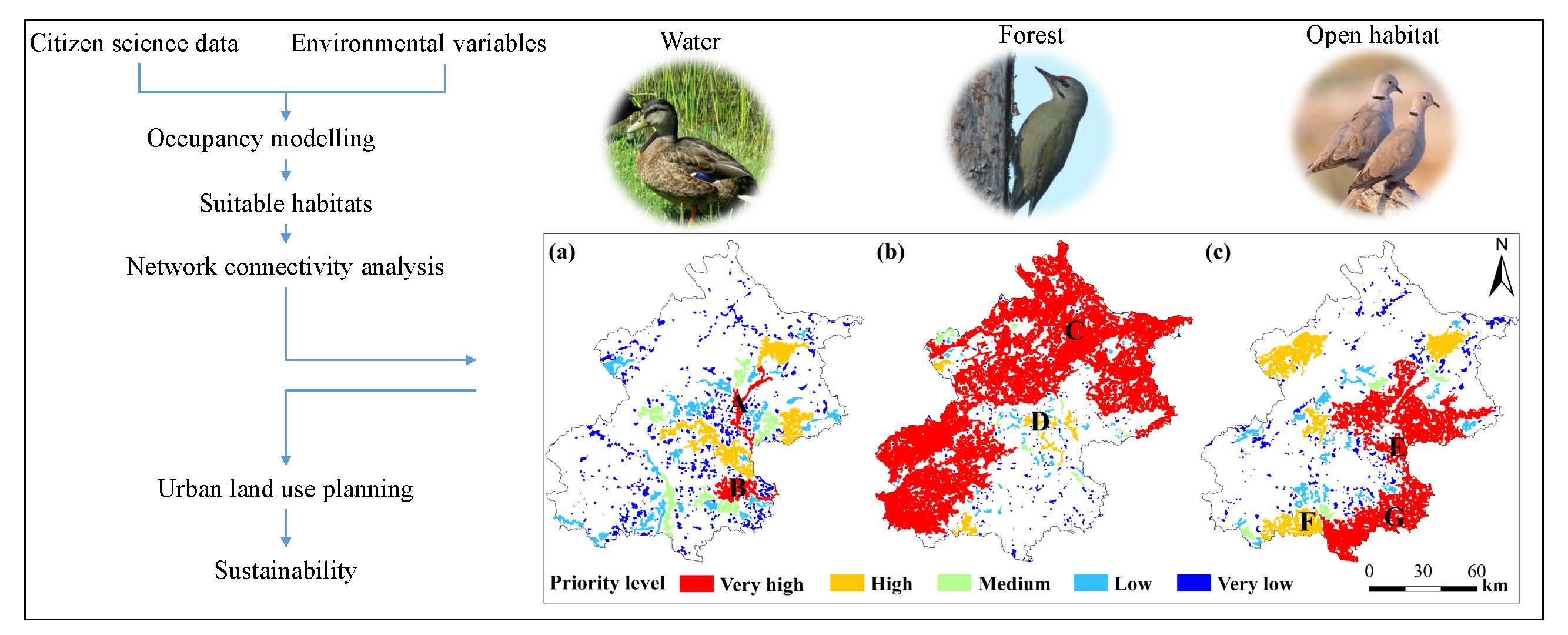
2.1. Study Area
2.2. Bird Data and Focal Species Selection
2.3. Environmental Variables
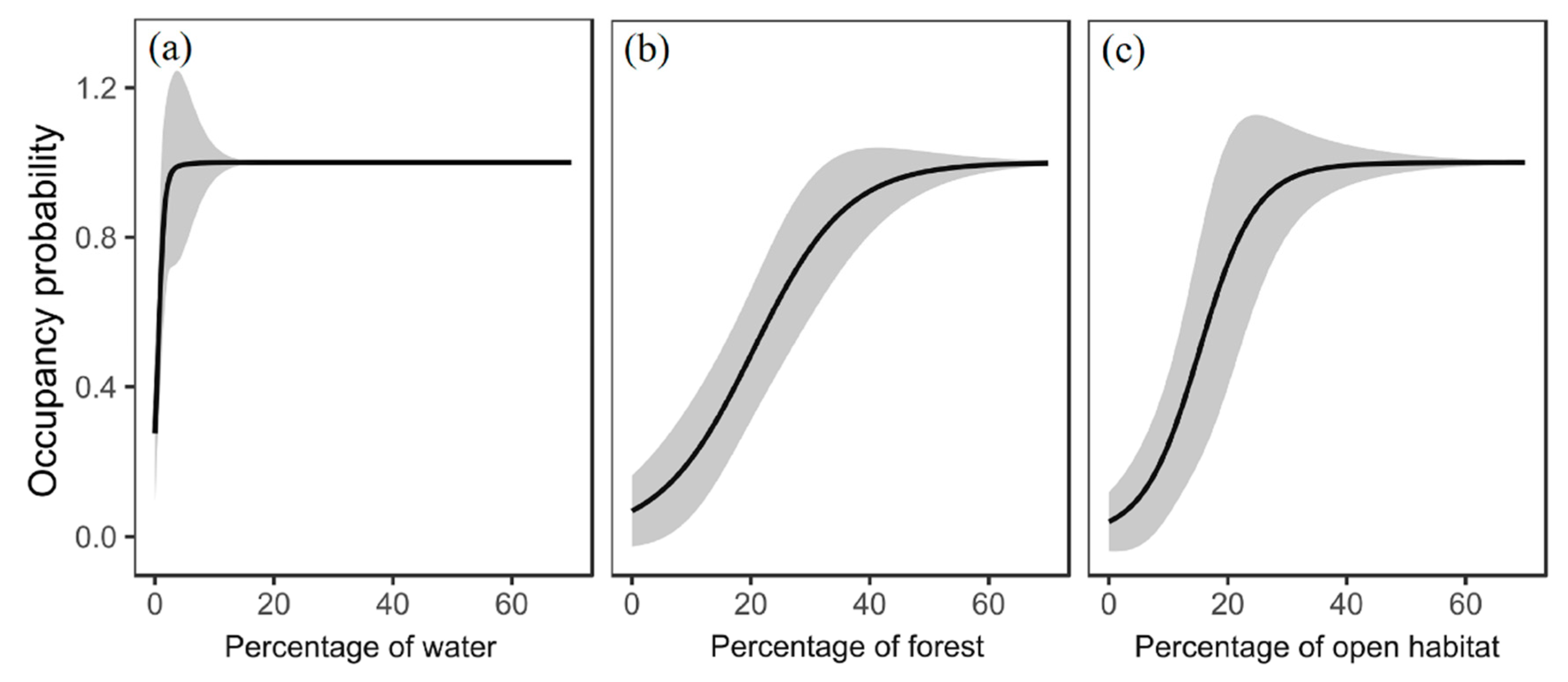
2.4. Habitat Suitability Modelling
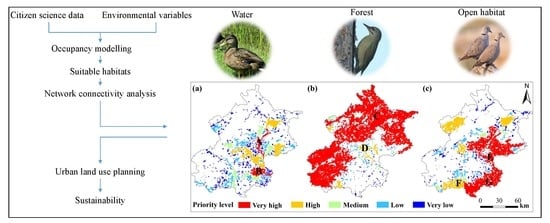
2.5. Habitat Network Connectivity Analysis
3. Results
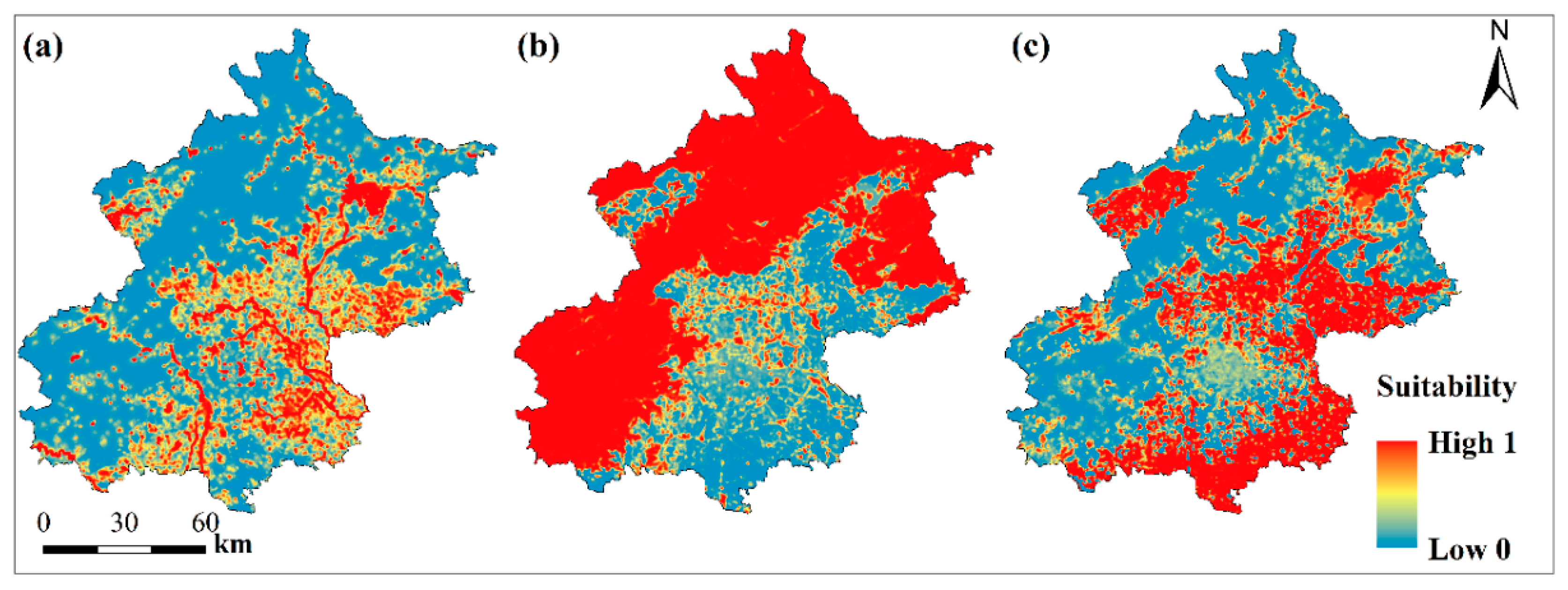
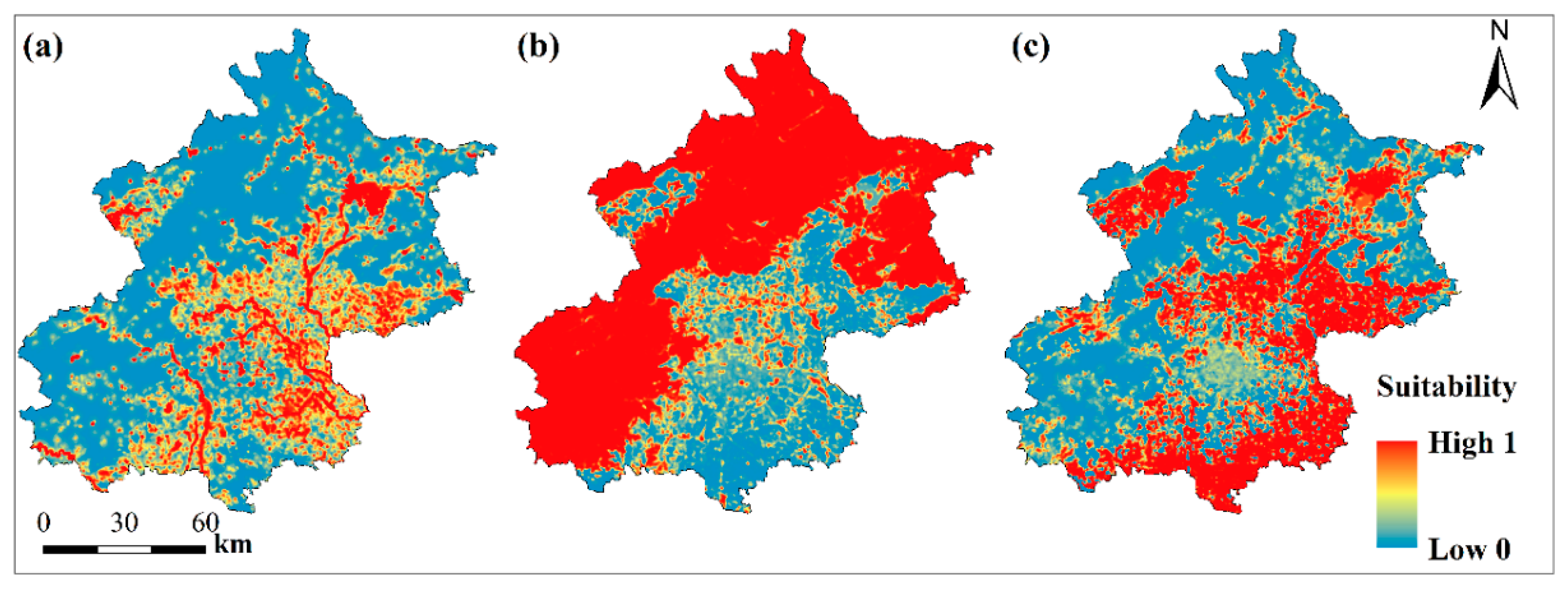
3.1. Habitat Suitability for Focal Species
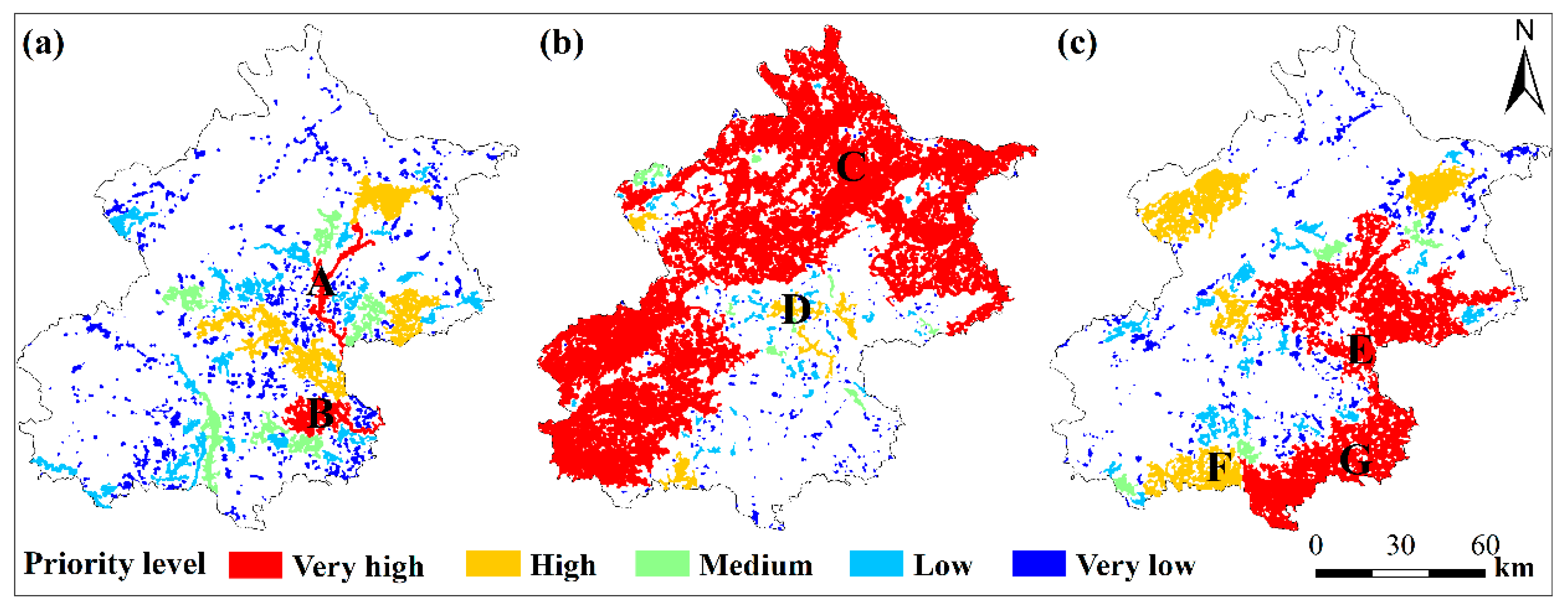
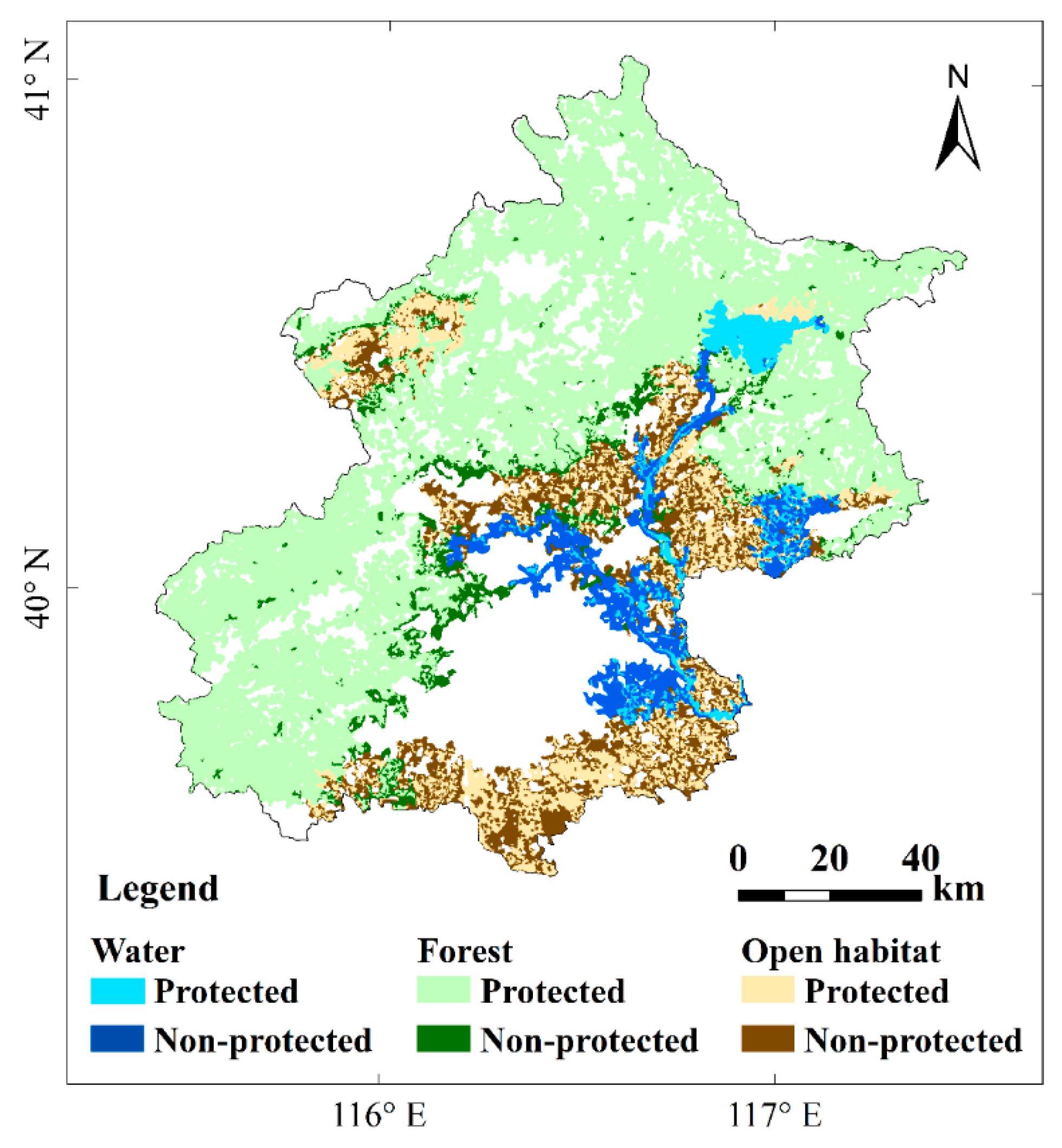
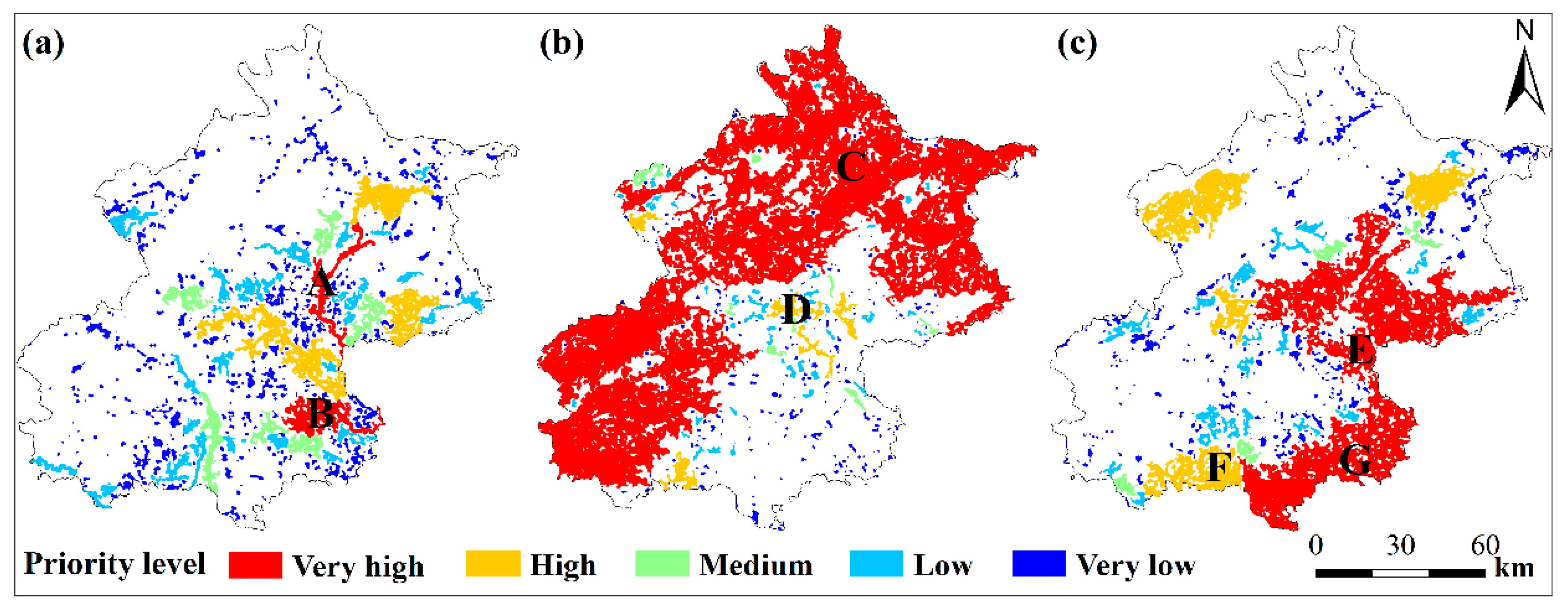
3.2. The Habitat Network Connectivity and Conservation Priority
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Id | English Name | Scientific Name | OF | LCI | UCI | VBCI | Score | |
|---|---|---|---|---|---|---|---|---|
| 1 | Mallard | Anas platyrhynchos | 0.313 | 0.453 | 0.073 | 0.183 | 0.111 | 0.586 |
| 2 | Little Grebe | Tachybaptus ruficollis | 0.317 | 0.373 | 0.082 | 0.214 | 0.132 | 0.555 |
| 3 | Eastern spot-billed Duck | Anas zonorhyncha | 0.354 | 0.213 | 0.114 | 0.326 | 0.212 | 0.485 |
| 4 | Black-crowned Night Heron | Nycticorax nycticorax | 0.278 | 0.340 | 0.055 | 0.186 | 0.132 | 0.533 |
| 5 | White wagtail | Motacilla alba | 0.214 | 0.367 | 0.054 | 0.187 | 0.133 | 0.521 |
| 6 | Eurasian teal | Anas crecca | 0.538 | 0.173 | 0.101 | 0.358 | 0.257 | 0.510 |
| 7 | Ruddy shelduck | Tadorna ferruginea | 0.517 | 0.167 | 0.106 | 0.372 | 0.265 | 0.499 |
| 8 | Common kingfisher | Alcedo atthis | 0.139 | 0.333 | 0.051 | 0.192 | 0.140 | 0.485 |
| 9 | Grey heron | Ardea cinerea | 0.240 | 0.240 | 0.066 | 0.262 | 0.196 | 0.465 |
| 10 | Mandarin duck | Aix galericulata | 0.142 | 0.213 | 0.077 | 0.288 | 0.211 | 0.422 |
| 11 | Ibisbill | Ibidorhyncha struthersii | 0.019 | 0.047 | 0.009 | 0.036 | 0.027 | 0.409 |
| 12 | Brown dipper | Cinclus pallasii | 0.020 | 0.040 | 0.007 | 0.034 | 0.026 | 0.407 |
| 13 | Crested kingfisher | Megaceryle lugubris | 0.152 | 0.173 | 0.040 | 0.268 | 0.228 | 0.406 |
| 14 | Plumbeous water redstart | Rhyacornis fuliginosa | 0.051 | 0.073 | −0.065 | 0.283 | 0.348 | 0.298 |
| Id | English Name | Scientific Name | OF | LCI | UCI | VBCI | Score | |
|---|---|---|---|---|---|---|---|---|
| 1 | Grey-headed woodpecker | Picus canus | 0.554 | 0.433 | 0.389 | 0.518 | 0.130 | 0.644 |
| 2 | Great tit | Parus major | 0.575 | 0.440 | 0.419 | 0.576 | 0.157 | 0.642 |
| 3 | Great spotted woodpecker | Dendrocopos major | 0.400 | 0.553 | 0.327 | 0.454 | 0.127 | 0.635 |
| 4 | Light-vented bulbul | Pycnonotus sinensis | 0.399 | 0.547 | 0.301 | 0.428 | 0.127 | 0.633 |
| 5 | Grey-capped greenfinch | Chloris sinica | 0.526 | 0.407 | 0.356 | 0.512 | 0.155 | 0.618 |
| 6 | Marsh tit | Poecile palustris | 0.490 | 0.420 | 0.354 | 0.508 | 0.154 | 0.611 |
| 7 | Plain laughingthrush | Garrulax davidi | 0.706 | 0.253 | 0.492 | 0.709 | 0.217 | 0.601 |
| 8 | Vinous-throated parrotbill | Sinosuthora webbiana | 0.529 | 0.360 | 0.430 | 0.603 | 0.174 | 0.597 |
| 9 | Red-billed blue magpie | Urocissa erythroryncha | 0.578 | 0.320 | 0.430 | 0.620 | 0.190 | 0.593 |
| 10 | Meadow bunting | Emberiza cioides | 0.615 | 0.260 | 0.471 | 0.674 | 0.203 | 0.581 |
| 11 | Godlewski’s bunting | Emberiza godlewskii | 0.703 | 0.227 | 0.517 | 0.763 | 0.245 | 0.581 |
| 12 | Yellow-bellied tit | Pardaliparus venustulus | 0.595 | 0.280 | 0.360 | 0.572 | 0.212 | 0.578 |
| 13 | Grey-capped pygmy woodpecker | Dendrocopos canicapillus | 0.423 | 0.373 | 0.331 | 0.485 | 0.154 | 0.577 |
| 14 | Silver-throated bushtit | Aegithalos glaucogularis | 0.628 | 0.253 | 0.427 | 0.655 | 0.228 | 0.573 |
| 15 | Chinese hill warbler | Rhopophilus pekinensis | 0.642 | 0.240 | 0.468 | 0.697 | 0.228 | 0.573 |
| 16 | Common pheasant | Phasianus colchicus | 0.518 | 0.300 | 0.458 | 0.656 | 0.198 | 0.566 |
| 17 | Koklass pheasant | Pucrasia macrolopha | 0.959 | 0.027 | 0.866 | 1.030 | 0.164 | 0.630 |
| 18 | Collared scops owl | Otus lettia | 0.794 | 0.013 | 0.738 | 0.850 | 0.112 | 0.597 |
| 19 | Tawny owl | Strix aluco | 0.808 | 0.033 | 0.699 | 0.968 | 0.269 | 0.545 |
| 20 | Eurasian wren | Troglodytes troglodytes | 0.654 | 0.167 | 0.417 | 0.675 | 0.258 | 0.543 |
| 21 | Willow tit | Poecile montanus | 0.825 | 0.093 | 0.467 | 0.803 | 0.335 | 0.541 |
| 22 | Eurasian jay | Garrulus glandarius | 0.880 | 0.053 | 0.568 | 0.918 | 0.350 | 0.540 |
| 23 | Spotted nutcracker | Nucifraga caryocatactes | 0.826 | 0.040 | 0.583 | 0.891 | 0.308 | 0.536 |
| 24 | Grey-sided thrush | Turdus feae | 0.818 | 0.060 | 0.424 | 0.831 | 0.408 | 0.500 |
| 25 | Crested myna | AcridStraggleres cristatellus | 0.292 | 0.267 | 0.274 | 0.446 | 0.172 | 0.499 |
| 26 | Coal tit | Periparus ater | 0.546 | 0.120 | 0.398 | 0.748 | 0.350 | 0.460 |
| 27 | Little owl | Athene noctua | 0.326 | 0.113 | 0.171 | 0.456 | 0.285 | 0.418 |
| 28 | Chinese thrush | Turdus mupinensis | 0.468 | 0.087 | 0.246 | 0.618 | 0.372 | 0.418 |
| 29 | Eurasian eagle-owl | Bubo bubo | 0.312 | 0.087 | 0.143 | 0.481 | 0.337 | 0.385 |
| 30 | Eurasian treecreeper | Certhia familiaris | 0.474 | 0.027 | 0.235 | 0.713 | 0.478 | 0.359 |
| 31 | Chinese beautiful rosefinch | Carpodacus davidianus | 0.552 | 0.040 | 0.402 | 0.998 | 0.595 | 0.340 |
| 32 | Japanese tit | Parus minor | 0.480 | 0.020 | 0.074 | 0.759 | 0.684 | 0.276 |
| Id | English Name | Scientific Name | OF | LCI | UCI | VBCI | Score | |
|---|---|---|---|---|---|---|---|---|
| 1 | Eurasian collared dove | Streptopelia decaocto | 0.344 | 0.207 | 0.123 | 0.279 | 0.156 | 0.503 |
| 2 | Spotted Dove | Spilopelia chinensis | 0.130 | 0.553 | 0.088 | 0.168 | 0.080 | 0.573 |
| 3 | Common Kestrel | Falco tinnunculus | 0.194 | 0.407 | 0.113 | 0.215 | 0.103 | 0.539 |
| 4 | Eurasian Hoopoe | Upupa epops | 0.203 | 0.327 | 0.113 | 0.227 | 0.114 | 0.513 |
| 5 | Oriental Turtle Dove | Streptopelia orientalis | 0.196 | 0.267 | 0.121 | 0.257 | 0.137 | 0.484 |
| 6 | Hill Pigeon | Columba rupestris | 0.165 | 0.167 | 0.103 | 0.269 | 0.166 | 0.433 |
| 7 | Red-billed Starling | Spodiopsar sericeus | 0.057 | 0.140 | 0.021 | 0.134 | 0.113 | 0.414 |
| 8 | Golden Eagle | Aquila chrysaetos | 0.121 | 0.100 | 0.071 | 0.220 | 0.148 | 0.407 |
| 9 | Daurian Partridge | Perdix dauurica | 0.321 | 0.020 | 0.075 | 0.352 | 0.277 | 0.391 |
| 10 | Asian Short-toed Lark | Calandrella cheleensis | 0.319 | 0.060 | 0.071 | 0.390 | 0.319 | 0.386 |
| 11 | White-bellied Redstart | Hodgsonius phoenicuroides | 0.051 | 0.027 | 0.041 | 0.176 | 0.135 | 0.369 |
| 12 | Long-tailed Shrike | Lanius schach | 0.096 | 0.040 | 0.005 | 0.217 | 0.212 | 0.356 |
| 13 | Chukar Partridge | Alectoris chukar | 0.099 | 0.027 | 0.010 | 0.291 | 0.281 | 0.325 |
| 14 | Crested Lark | Galerida cristata | 0.525 | 0.020 | −0.004 | 0.780 | 0.784 | 0.250 |
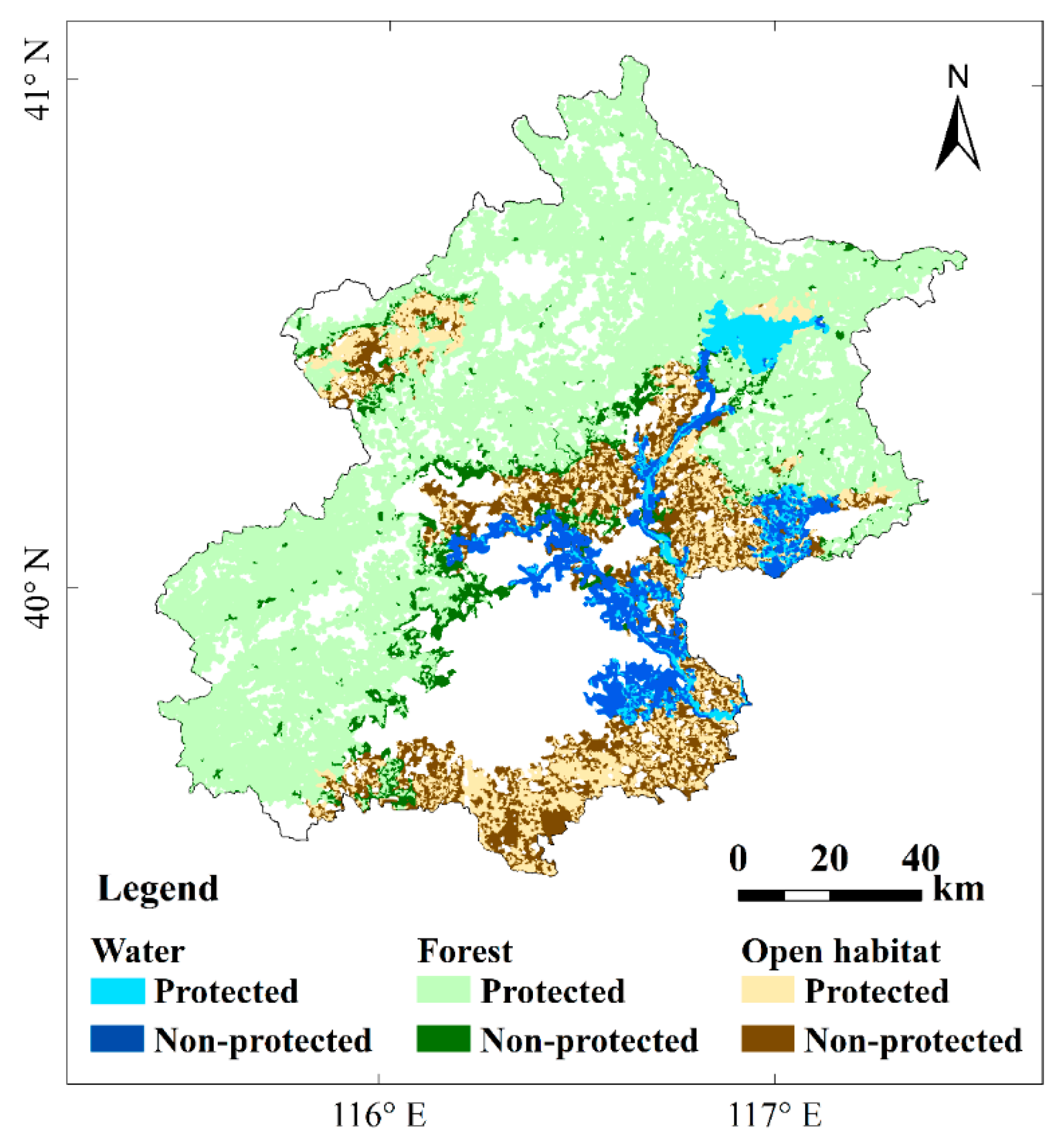
| Birds | Id | Patch Name | Area/km2 | dPC/% | Situation in Network | Location/(E°, N°) | |
|---|---|---|---|---|---|---|---|
| Water | 467 * | Chaobai River in Shunyi district | 159.9 | 34.2 | Center of the habitat network | 116.741 | 40.206 |
| 499 * | Liangshui & Xiaotaihou Rivers, wetlands and grass in Tongzhou district | 234.1 | 24.8 | Large patch | 116.729 | 39.807 | |
| 71 * | Wenyu and Ba Rivers in Chaoyang district | 122.0 | 18.6 | Large patch and close to key patches 502, 503 | 116.605 | 39.990 | |
| 446 | Miyun Reservoir | 205.8 | 18.6 | Large patch | 116.943 | 40.502 | |
| 502 * | North Canal and Yunchaojian Rivers in Tongzhou district | 56.3 | 18.3 | Close to large patch 71, 499 | 116.717 | 39.908 | |
| 464 * | Ju and Ru Rivers in Pinggu district | 190.4 | 15.8 | Large patch | 117.038 | 40.129 | |
| 504 * | Sha and Wenyu Rivers in Changping district | 120.5 | 14.7 | Center of the habitat network | 116.348 | 40.115 | |
| 503 * | Qing River and Olympic Park in Chaoyang district | 80.4 | 13.6 | Close to key patch 71, 504 and center of the habitat network | 116.447 | 40.045 | |
| 501 * | Yueya & Zhongba Rivers and Bojue golf club in Tongzhou district | 22.3 | 12.8 | Close to large patch 71 | 116.714 | 40.000 | |
| Forest | 1 | Mountains in Yanqing, Huairou and Miyun districts | 5289.2 | 86.1 | Large patch | 116.661 | 40.492 |
| 280 | Mountains in Fangshan and Shijingshan districts | 1821.0 | 39.3 | Large patch | 115.934 | 39.852 | |
| 281 | Mountains in Mentougou district | 1011.7 | 34.8 | Large patch and close to large patch 280, 1 | 115.773 | 40.045 | |
| 68 * | Forest along the Wenyu River and 6th north ring road in Shunyi district | 62.4 | 1.1 | Center of the habitat network | 116.496 | 40.158 | |
| 244 * | Forest along the Dashi River in Liulihe town | 66.0 | 0.9 | Close to large patch 279 | 116.015 | 39.628 | |
| 127 * | Forest along the Wenyu River in Chaoyang district | 41.7 | 0.7 | Center of the habitat network | 116.574 | 40.018 | |
| 34 | Forest around Guanting Reservoir and Guishui River in Yanqing district | 35.7 | 0.7 | Close to large patch 1 | 115.854 | 40.429 | |
| 70 * | Forest along the Chaobai River in Shunyi district | 45.3 | 0.6 | Center of the habitat network | 116.698 | 40.129 | |
| Open habitat | 206 | Grass/cropland in Daxing and Tongzhou districts | 1090.6 | 53.8 | Large patch | 116.575 | 39.693 |
| 78 * | Grass/cropland in Changping, Shunyi and Huairou districts | 613.0 | 39.8 | Large patch | 116.558 | 40.263 | |
| 324 * | Grass/cropland in Shunyi and Pinggu districts | 795.4 | 36.1 | Large patch | 116.953 | 40.189 | |
| 307 * | Grass/cropland between Tong zhou, Chaoyang and Shunyi districts | 193.7 | 34.1 | Closed to large patches 206, 78,324 | 116.642 | 40.014 | |
| 310 * | Grass/cropland in Fangshan district | 312.5 | 16.5 | Large patch and close to large patch 206 | 116.012 | 39.634 | |
| 48 | Grass/cropland around Miyun Reservoir | 222.7 | 9.4 | Large patch | 117.025 | 40.543 | |
| 54 | Grass/cropland in Yanqing district | 455.9 | 6.9 | Large patch | 116.005 | 40.480 | |
| 319 * | Grass/cropland around Yangfang town | 117.9 | 6.9 | Close to large patch 78 and center of the habitat network | 116.168 | 40.137 | |
References
- United Nations: Department of Economic and Social Affairs. World Urbanization Prospects: The 2018 Revision; United Nations: Department of Economic and Social Affairs: New York, NY, USA, 2018. [Google Scholar]
- China Statistical Press. China Statistical Yearbook in 2018; China Statistical Press: Beijing, China, 2018. (In Chinese) [Google Scholar]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global Change and the Ecology of Cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef]
- Xun, B.; Yu, D.; Liu, Y.; Hao, R.; Sun, Y. Quantifying isolation effect of urban growth on key ecological areas. Ecol. Eng. 2014, 69, 46–54. [Google Scholar] [CrossRef]
- Ortega-Álvarez, R.; MacGregor-Fors, I. Living in the big city: Effects of urban land-use on bird community structure, diversity, and composition. Landsc. Urban Plan. 2009, 90, 189–195. [Google Scholar] [CrossRef]
- Canedoli, C.; Orioli, V.; Padoa-Schioppa, E.; Bani, L.; Dondina, O. Temporal Variation of Ecological Factors Affecting Bird Species Richness in Urban and Peri-Urban Forests in a Changing Environment: A Case Study from Milan (Northern Italy). Forests 2017, 8, 507. [Google Scholar] [CrossRef]
- Alberti, M. The Effects of Urban Patterns on Ecosystem Function. Int. Reg. Sci. Rev. 2005, 28, 168–192. [Google Scholar] [CrossRef]
- Opdam, P.; Steingröver, E.; Rooij, S.V. Ecological networks: A spatial concept for multi-actor planning of sustainable landscapes. Landsc. Urban Plan. 2006, 75, 322–332. [Google Scholar] [CrossRef]
- Defries, R.S.; Foley, J.A.; Asner, G.P. Land-use choices: Balancing human needs and ecosystem function. Front. Ecol. Environ. 2004, 2, 249–257. [Google Scholar] [CrossRef]
- Wilson, E.; MacArthur, R. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Diamond, J.M. The island dilemma: Lessons of modern biogeographic studies for the design of natural reserves. Biol. Conserv. 1975, 7, 129–146. [Google Scholar] [CrossRef]
- Cody, M.L.; Diamond, J.M. Ecology and Evolution of Communities. Nature 1976, 260, 204. [Google Scholar] [CrossRef]
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68, 571–573. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Xun, B.; Yu, D.; Wang, X. Prioritizing habitat conservation outside protected areas in rapidly urbanizing landscapes: A patch network approach. Landsc. Urban Plan. 2017, 157, 532–541. [Google Scholar] [CrossRef]
- Uezu, A.; Metzger, J.P.; Vielliard, J.M.E. Effects of structural and functional connectivity and patch size on the abundance of seven Atlantic Forest bird species. Biol. Conserv. 2005, 123, 507–519. [Google Scholar] [CrossRef]
- Hüse, B.; Szabó, S.; Deák, B.; Tóthmérész, B. Mapping an ecological network of green habitat patches and their role in maintaining urban biodiversity in and around Debrecen city (Eastern Hungary). Land Use Policy 2016, 57, 574–581. [Google Scholar] [CrossRef]
- Saura, S.; Rubio, L. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography 2010, 33, 523–537. [Google Scholar] [CrossRef]
- Pascual-Hortal, L.; Saura, S. Impact of spatial scale on the identification of critical habitat patches for the maintenance of landscape connectivity. Landsc. Urban Plan. 2007, 83, 176–186. [Google Scholar] [CrossRef]
- Kang, W.; Minor, E.S.; Park, C.R.; Lee, D. Effects of habitat structure, human disturbance, and habitat connectivity on urban forest bird communities. Urban Ecosyst. 2015, 18, 857–870. [Google Scholar] [CrossRef]
- Fajardo, J.; Lessmann, J.; Bonaccorso, E.; Devenish, C.; Muñoz, J. Combined use of systematic conservation planning, species distribution modelling, and connectivity analysis reveals severe conservation gaps in a megadiverse country (Peru). PLoS ONE 2014, 9, e114367. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, L.; Liu, J.; Zhuang, Z.; Huang, Q.; Li, M. Constructing Ecological Networks Based on Habitat Quality Assessment: A Case Study of Changzhou, China. Sci. Rep. 2017, 7, 46073. [Google Scholar] [CrossRef]
- Dondina, O.; Orioli, V.; Colli, L.; Luppi, M.; Bani, L. Ecological network design from occurrence data by simulating species perception of the landscape. Landsc. Ecol. 2018, 33, 275–287. [Google Scholar] [CrossRef]
- Dondina, O.; Orioli, V.; Chiatante, G.; Alberto, M.; Bani, L. Species Specialization Limits Movement Ability and Shapes Ecological Networks: The Case Study of two Forest Mammals. Curr. Zool. 2018. [Google Scholar] [CrossRef]
- Dondina, O.; Saura, S.; Bani, L.; Mateo-Sánchez, M.C. Enhancing connectivity in agroecosystems: Focus on the best existing corridors or on new pathways? Landsc. Ecol. 2018, 33, 1741–1756. [Google Scholar] [CrossRef]
- Li, F.; Wang, R.; Paulussen, J.; Liu, X. Comprehensive concept planning of urban greening based on ecological principles: A case study in Beijing, China. Landsc. Urban Plan. 2005, 72, 325–336. [Google Scholar] [CrossRef]
- Kong, F.; Yin, H.; Nakagoshi, N.; Zong, Y. Urban green space network development for biodiversity conservation: Identification based on graph theory and gravity modeling. Landsc. Urban Plan. 2010, 95, 16–27. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen Science as an Ecol. Res. Tool: Challenges and Benefits. Annu. Rev. Ecol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef]
- Strien, A.J.; Swaay, C.A.M.; Termaat, T.; Devictor, V. Opportunistic citizen science data of animal species produce reliable estimates of distribution trends if analysed with occupancy models. J. Appl. Ecol. 2013, 50, 1450–1458. [Google Scholar] [CrossRef]
- Mackenzie, D.I. Occupancy Estimation and Modeling; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- De Wan, A.A.; Sullivan, P.J.; Lembo, A.J.; Smith, C.R.; Maerz, J.C.; Lassoie, J.P.; Richmond, M.E. Using occupancy models of forest breeding birds to prioritize conservation planning. Biol. Conserv. 2009, 142, 982–991. [Google Scholar] [CrossRef]
- Padoa-Schioppa, E.; Baietto, M.; Massa, R.; Bottoni, L. Bird communities as bioindicators: The focal species concept in agricultural landscapes. Ecol. Indic. 2006, 6, 83–93. [Google Scholar] [CrossRef]
- Ogden, J.C.; Baldwin, J.D.; Bass, O.L.; Browder, J.A.; Cook, M.I.; Frederick, P.C.; Frezza, P.E.; Galvez, R.A.; Hodgson, A.B.; Meyer, K.D.; et al. Waterbirds as indicators of ecosystem health in the coastal marine habitats of southern Florida: 1. Selection and justification for a suite of indicator species. Ecol. Indic. 2014, 44, 148–163. [Google Scholar] [CrossRef]
- Rahman, F.; Ismail, A. Waterbirds: An important bio-indicators of ecosystem. Pertanika J. Schol. Res. Rev. 2018, 4, 81–90. [Google Scholar]
- Li, X.; Gong, P.; Liang, L. A 30-year (1984–2013) record of annual urban dynamics of Beijing City derived from Landsat data. Remote Sens. Environ. 2015, 166, 78–90. [Google Scholar] [CrossRef]
- Strohbach, M.W.; Lerman, S.B.; Warren, P.S. Are small greening areas enhancing bird diversity? Insights from community-driven greening projects in Boston. Landsc. Urban Plan. 2013, 114, 69–79. [Google Scholar] [CrossRef]
- Silva de Araújo, M.L.V.; Bernard, E. Green remnants are hotspots for bat activity in a large Brazilian urban area. Urban Ecosyst. 2016, 19, 287–296. [Google Scholar] [CrossRef]
- Fuyuki, A.; Yamaura, Y.; Nakajima, Y.; Ishiyama, N.; Akasaka, T.; Nakamura, F. Pond area and distance from continuous forests affect amphibian egg distributions in urban green spaces: A case study in Sapporo, Japan. Urban For. Urban Green. 2014, 13, 397–402. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Walker, K.; Williams, N.S.G.; Hahs, A.K.; Mata, L.; Stork, N.; Livesley, S.J. The conservation value of urban green space habitats for Australian native bee communities. Biol. Conserv. 2015, 187, 240–248. [Google Scholar] [CrossRef]
- Luppi, M.; Dondina, O.; Orioli, V.; Bani, L. Local and landscape drivers of butterfly richness and abundance in a human-dominated area. Agric. Ecosyst. Environ. 2018, 254, 138–148. [Google Scholar] [CrossRef]
- Hu, W.; Wang, S.; Li, D. Biological conservation security patterns plan in Beijing based on the focal species approach (in Chinese). Acta Ecol. Sin. 2010, 30, 4266–4276. [Google Scholar]
- China Bird Report Center. Available online: http://www.birdreport.cn (accessed on 21 December 2017).
- Li, X.; Liang, L.; Gong, P.; Liu, Y.; Liang, F. Bird watching in China reveals bird distribution changes. Chin. Sci. Bull. 2013, 58, 649–656. [Google Scholar] [CrossRef]
- Hamish, W.; Jonathan, B.; Jennifer, S.; Carolina, D.L.R.; Rivadeneira, M.; Walter, J. EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 2014, 95, 2027. [Google Scholar] [CrossRef]
- Brazil, M. Birds of East Asia; Christopher Helm: London, UK, 2009. [Google Scholar]
- Lambeck, R.J. Focal Species: A multi-species umbrella for nature conservation. Conserv. Biol. 1997, 11, 849–856. [Google Scholar] [CrossRef]
- Bani, L.; Baietto, M.; Bottoni, L.; Massa, R. The use of focal species in designing a habitat network for a lowland area of Lombardy, Italy. Conserv. Biol. 2002, 16, 826–831. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 2.8-19. Available online: https://cran.r-project.org/web/packages/raster/index.html (accessed on 25 March 2017).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Huang, C.; Yang, J.; Lu, H.; Huang, H.; Yu, L. Green Spaces as an Indicator of Urban Health: Evaluating Its Changes in 28 Mega-Cities. Remote Sens. 2017, 9, 1266. [Google Scholar] [CrossRef]
- Advanced Spaceborne Thermal Emission and Reflection Radiometer Global Digital Elevation Model. Available online: http://reverb.echo.nasa.gov/reverb/ (accessed on 27 May 2018).
- Vector Data of Major Roads. Available online: http://www.bjdata.gov.cn/zyml/ajg/sgtj/5173.htm (accessed on 15 August 2018).
- Operational Linescan System (OLS) by the U.S. Air Force Defense Meteorological Satellite Program (DMSP)/Version 4 DMSP-OLS Nighttime Lights Time Series. Available online: https://ngdc.noaa.gov/eog/dmsp/downloadV4composites.html (accessed on 18 August 2018).
- Land Surface Temperature/Emissivity Monthly L3 Global 0.05Deg CMG. Available online: https://e4ftl01.cr.usgs.gov/MOLT/ (accessed on 5 August 2018).
- European Centre for Medium-Range Weather Forecasts (ECMWF) ERA-Interim, Monthly means of Daily Forecast Accumulations. Available online: http://apps.ecmwf.int/datasets/data/interim-mdfa/levtype=sfc/ (accessed on 6 August 2018).
- Huang, Y.; Zhao, Y.; Li, S.; von Gadow, K. The Effects of habitat area, vegetation structure and insect richness on breeding bird populations in Beijing urban parks. Urban For. Urban Green. 2015, 14, 1027–1039. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical Maps; Computer software program produced by the authors at the University of Massachusetts: Amherst, MA, USA, 2012; Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 13 April 2017).
- Li, X.; Si, Y.; Ji, L.; Gong, P. Dynamic response of East Asian Greater White-fronted Geese to changes of environment during migration: Use of multi-temporal species distribution model. Ecol. Model. 2017, 360, 70–79. [Google Scholar] [CrossRef]
- Rosin, Z.M.; Piotr, S.; Paweł, S.; Marcin, T.; Andrzej, L.; Piotr, T. Constant and seasonal drivers of bird communities in a wind farm: Implications for conservation. PeerJ 2016, 4, e2105. [Google Scholar] [CrossRef]
- Summers, P.D.; Cunnington, G.M.; Fahrig, L. Are the negative effects of roads on breeding birds caused by traffic noise? J. Appl. Ecol. 2011, 48, 1527–1534. [Google Scholar] [CrossRef]
- Orłowski, G. Roadside hedgerows and trees as factors increasing road mortality of birds: Implications for management of roadside vegetation in rural landscapes. Landsc. Urban Plan. 2008, 86, 153–161. [Google Scholar] [CrossRef]
- Ghosh, T.; Anderson, S.J.; Elvidge, C.D.; Sutton, P.C. Using nighttime satellite imagery as a proxy measure of human well-being. Sustainability 2013, 5, 4988–5019. [Google Scholar] [CrossRef]
- Strittholt, J.R.; Dellasala, D.A. Importance of Roadless Areas in Biodiversity Conservation in Forested Ecosystems: Case Study of the Klamath-Siskiyou Ecoregion of the United States. Conserv. Biol. 2001, 15, 1742–1754. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10.3.1; Environmental Systems Research Institute: Redlands, CA, USA, 2015. [Google Scholar]
- Fiske, I.J.; Chandler, R.B. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference, R Package Version 1.42.1; Available online: https://cran.rstudio.com/web/packages/MuMIn/ (accessed on 23 November 2017).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- MacKenzie, D.I.; Bailey, L.L. Assessing the fit of site-occupancy models. J. Agric. Biol. Environ. Stat. 2004, 9, 300–318. [Google Scholar] [CrossRef]
- Symonds, M.R.E.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Richard, G. Pearson Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Saura, S.; Torné, J. Conefor Sensinode 2.2: A software package for quantifying the importance of habitat patches for landscape connectivity. Environ. Model. Softw. 2009, 24, 135–139. [Google Scholar] [CrossRef]
- Jenks, G.F. The Data Model Concept in Statistical Mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Shimazaki, H.; Tamura, M.; Darman, Y.; Andronov, V.; Parilov, M.P.; Nagendran, M.; Higuchi, H. Network analysis of potential migration routes for Oriental White Storks (Ciconia boyciana). Ecol. Res. 2004, 19, 683–698. [Google Scholar] [CrossRef]
- Saura, S.; Bodin, Ö.; Fortin, M.-J. Editor’s choice: Stepping stones are crucial for species’ long-distance dispersal and range expansion through habitat networks. J. Appl. Ecol. 2014, 51, 171–182. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y. Applying various algorithms for species distribution modelling. Integr. Zool. 2013, 8, 124–135. [Google Scholar] [CrossRef]
- Tiwary, N.K.; Urfi, A.J. Spatial variations of bird occupancy in Delhi: The significance of woodland habitat patches in urban centres. Urban For. Urban Green. 2016, 20, 338–347. [Google Scholar] [CrossRef]
- Merken, R.; Deboelpaep, E.; Teunen, J.; Saura, S.; Koedam, N. Wetland Suitability and Connectivity for Trans-Saharan Migratory Waterbirds. PLoS ONE 2015, 10, e0135445. [Google Scholar] [CrossRef]


| Variables | Data Source | Data Resolution | |
|---|---|---|---|
| Site-level | Percentage of land cover types & distance to the nearest water body (Dis2water) | 2015 land cover map [51] | 30 m |
| Elevation (Elev) | Advanced Spaceborne Thermal Emission and Reflection Radiometer Global Digital Elevation Model (ASTER GDEM) data [52] | 30 m | |
| Distance to the nearest major road (Dis2road) | Vector data of major roads [53] | 30 m | |
| Nightlight index (Light) | Operational Linescan System (OLS) by the U.S. Air Force Defense Meteorological Satellite Program (DMSP) [54] | 1 km | |
| Observation-level | Date | 2008–2017 Bird observation data from China Bird Report Center [43] | daily |
| Land surface temperature | Land Surface Temperature/Emissivity Monthly L3 Global 0.05Deg CMG (the NASA Land Processes Distributed Active Archive Center [55]) | 0.05 degree | |
| Precipitation | European Centre for Medium-Range Weather Forecasts (ECMWF) ERA-Interim, Monthly means of Daily Forecast Accumulations [56] | 0.75 degree |
| Birds | Models |
|---|---|
| Water | ψ(water + bareland + grass + dis2water + dis2road)p(date + temperature + precipitation) |
| Forest | ψ(water + bareland + forest + grass + cropland)p(date + temperature + precipitation) |
| Open-habitat | ψ(water + bareland + forest + grass/cropland + dis2water)p(date + temperature + precipitation) |
| Birds | Models | Intercept | Water | Bareland | Forest | Grass | Cropland | Dis2water | Dis2road |
|---|---|---|---|---|---|---|---|---|---|
| Water | Coefficient | 11.63 | 31.39 | 0.81 | - | 0.81 | - | −0.82 | 0.25 |
| SE | 5.19 | 13.56 | 0.57 | - | 0.36 | - | 0.65 | 0.34 | |
| p-value | 0.03 | 0.02 | 0.15 | - | 0.02 | - | 0.21 | 0.46 | |
| Forest | Coefficient | 2.68 | 0.07 | −0.40 | 3.89 | 0.05 | −0.21 | - | - |
| SE | 0.88 | 0.29 | 0.61 | 1.09 | 0.32 | 0.42 | - | - | |
| p-value | 0.00 | 0.81 | 0.52 | 0.00 | 0.89 | 0.62 | - | - | |
| Open habitat | Coefficient | −0.09 | 0.22 | −2.52 | −0.36 | 4.13 * | −1.37 | - | |
| SE | 0.59 | 0.34 | 1.42 | 0.48 | 1.55 * | 0.84 | - | ||
| p-value | 0.88 | 0.53 | 0.08 | 0.45 | 0.01 * | 0.10 | - | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Z.; Yang, J.; Wielstra, B.; Wei, J.; Xu, F.; Si, Y. Prioritizing Green Spaces for Biodiversity Conservation in Beijing Based on Habitat Network Connectivity. Sustainability 2019, 11, 2042. https://doi.org/10.3390/su11072042
Lv Z, Yang J, Wielstra B, Wei J, Xu F, Si Y. Prioritizing Green Spaces for Biodiversity Conservation in Beijing Based on Habitat Network Connectivity. Sustainability. 2019; 11(7):2042. https://doi.org/10.3390/su11072042
Chicago/Turabian StyleLv, Zhiyuan, Jun Yang, Ben Wielstra, Jie Wei, Fei Xu, and Yali Si. 2019. "Prioritizing Green Spaces for Biodiversity Conservation in Beijing Based on Habitat Network Connectivity" Sustainability 11, no. 7: 2042. https://doi.org/10.3390/su11072042
APA StyleLv, Z., Yang, J., Wielstra, B., Wei, J., Xu, F., & Si, Y. (2019). Prioritizing Green Spaces for Biodiversity Conservation in Beijing Based on Habitat Network Connectivity. Sustainability, 11(7), 2042. https://doi.org/10.3390/su11072042