Enhanced Community Production rather than Structure Improvement under Nitrogen and Phosphorus Addition in Severely Degraded Alpine Meadows


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Field Sampling and Measurements
2.4. Data Calculation
2.5. Statistical Analysis
3. Results
3.1. Variability of Climatic Data
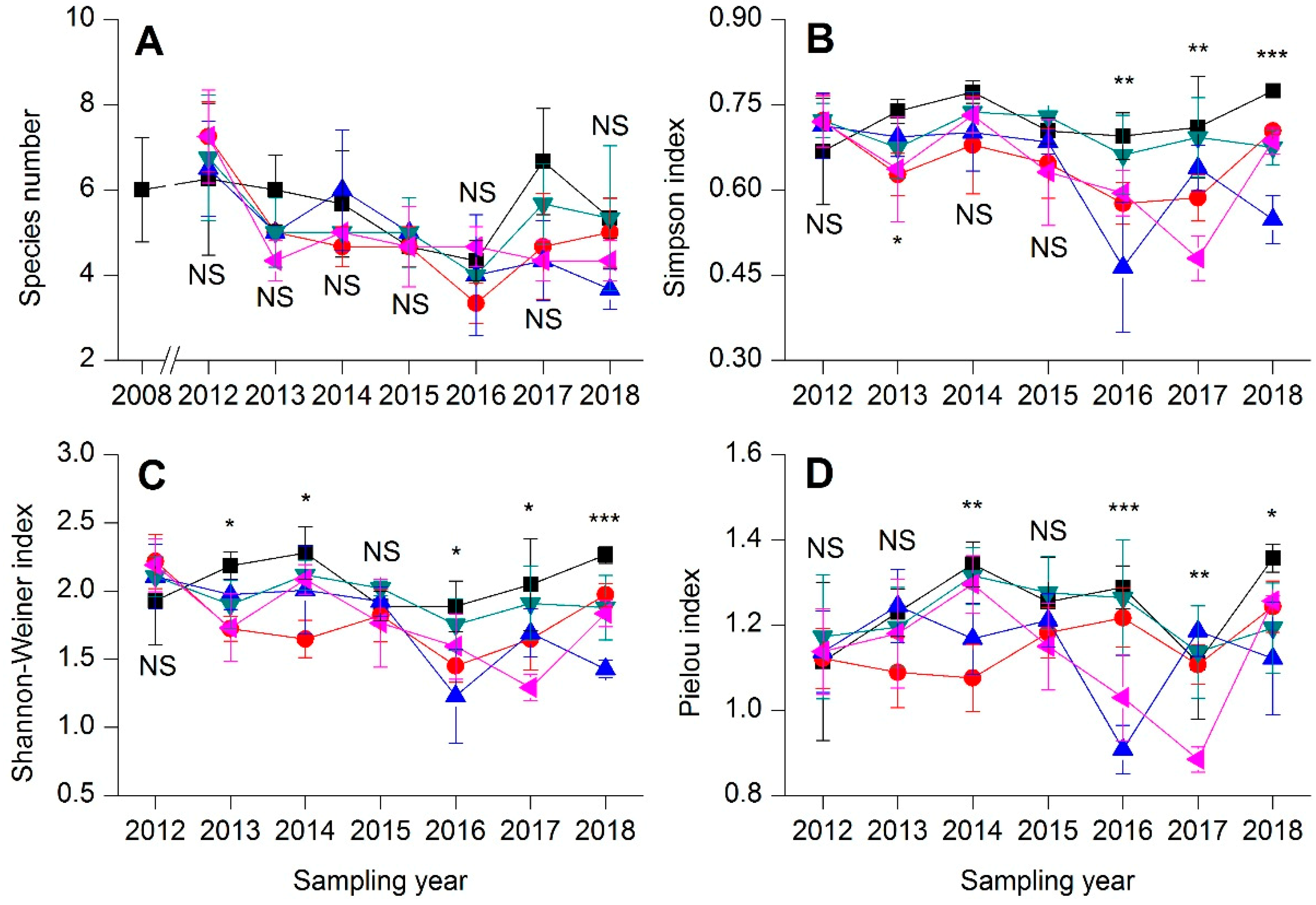
3.2. Community Composition, Species Diversity, and Temporal Stability
3.3. Community Production
3.4. Response of Different Plant Functional Groups to Fertilization Treatments
4. Discussion
4.1. Effect of Nitrogen Addition on Alpine Ecosystems
4.2. Effects of Nitrogen and Phosphorus Addition on Alpine Ecosystems
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Dong, S.; Gao, Q.; Liu, S.; Zhou, H.; Ganjurjav, H.; Wang, X. Climate change and human activities altered the diversity and composition of soil microbial community in alpine grasslands of the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 562, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Li, Y.; Wan, Y.; Qin, X.; Jiangcun, W.; Liu, Y. Dynamics of alpine grassland NPP and its response to climate change in Northern Tibet. Clim. Chang. 2009, 97, 515–528. [Google Scholar] [CrossRef]
- Gao, Q.; Li, Y.; Wan, Y.; Jiangcun, W.; Qin, X.; Wang, B. Significant achievements in protection and restoration of alpine grassland ecosystem in Northern Tibet, China. Restor. Ecol. 2009, 17, 320–323. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, S.; Gao, Q.; Liu, S.; Liang, Y.; Cao, X. Responses of alpine vegetation and soils to the disturbance of plateau pika (Ochotona curzoniae) at burrow level on the Qinghai-Tibetan Plateau of China. Ecol. Eng. 2016, 88, 232–236. [Google Scholar]
- Wang, K.; Deng, L.; Ren, Z.; Li, J.; Shangguan, Z. Grazing exclusion significantly improves grassland ecosystem C and N pools in a desert steppe of Northwest China. Catena 2016, 137, 441–448. [Google Scholar]
- Wu, J.; Shen, Z.; Shi, P.; Zhou, Y.; Zhang, X. Effects of grazing exclusion on plant functional group diversity alpine grasslands along a precipitation gradient on the Northern Tibetan Plateau. Arct. Antarct. Alp. Res. 2014, 46, 419–429. [Google Scholar] [CrossRef]
- Bai, Y.; Wu, J.; Clark, C.M.; Naeem, S.; Pan, Q.; Huang, J.; Zhang, L.; Han, X. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from inner Mongolia Grasslands. Glob. Chang. Biol. 2010, 16, 358–372. [Google Scholar]
- Jiang, J.; Zong, N.; Song, M.; Shi, P.; Ma, W.; Fu, G.; Shen, Z.; Zhang, X.; Ouyang, H. Responses of ecosystem respiration and its components to fertilization in an alpine meadow on the Tibetan Plateau. Eur. J. Soil Biol. 2013, 56, 101–106. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, F.; Chen, W.; Zhu, L.; Li, G.; Qi, D. Soil physical characteristics on different degraded alpine grasslands in Maqu County in upper Yellow River. Bull. Soil Water Conserv. 2010, 30, 16–21. [Google Scholar]
- Zhang, S.; Zhang, D.; Liu, X.; Chen, J.; Xun, C.; Duan, C. Soil nutrient characteristics of alpine meadow at different degradation degrees in Eastern Qilian Mountains. Pratacult. Sci. 2012, 29, 1028–1032. [Google Scholar]
- Bowman, W.D.; Cleveland, C.C.; Halada, L.; Hresko, J.; Baron, J.S. Negative impact of nitrogen deposition on soil buffering capacity. Nat. Geosci. 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Lieb, A.M.; Darrouzet-Nardi, A.; Bowman, W.D. Nitrogen deposition decreases acid buffering capacity of alpine soils in the southern Rocky Mountains. Geoderma 2011, 164, 220–224. [Google Scholar] [CrossRef]
- Chen, D.; Lan, Z.; Bai, X.; Grace, J.B.; Bai, Y. Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe. J. Ecol. 2013, 101, 1322–1334. [Google Scholar] [CrossRef]
- Wei, C.; Yu, Q.; Bai, E.; Lu, X.; Li, Q.; Xia, J.; Kardol, P.; Liang, W.; Wang, Z.; Han, X. Nitrogen deposition weakens plant-microbe interactions in grassland ecosystems. Glob. Chang. Biol. 2013, 19, 3688–3697. [Google Scholar] [CrossRef]
- Bowman, W.D.; Gartner, J.R.; Holland, K.; Wiedermann, M. Nitrogen critical loads for alpine vegetation and terrestrial ecosystem response: Are we there yet? Ecol. Appl. 2006, 16, 1183–1193. [Google Scholar] [CrossRef]
- Bowman, W.D.; Theodose, T.A.; Schardt, J.C.; Conant, R.T. Constraints of nutrient availability on primary production in two alpine tundra communities. Ecology 1993, 74, 2085–2097. [Google Scholar]
- Bowman, W.D. Accumulation and use of nitrogen and phosphorus following fertilization in two alpine tundra communities. Oikos 1994, 70, 261–270. [Google Scholar]
- Yang, X.; Ren, F.; Zhou, H.; He, J. Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang Plateau. Chin. J. Plant Ecol. 2014, 38, 159–166. [Google Scholar]
- Zong, N.; Shi, P.; Niu, B.; Jiang, J.; Song, M.; Zhang, X.; He, Y. Effects of nitrogen and phosphorous fertilization on community structure and productivity of degraded alpine meadows in northern Tibet, China. Chin. J. Appl. Ecol. 2014, 25, 3458–3468. [Google Scholar]
- Cao, G.M.; Zhang, J.X. Soil nutrition and substance cycle of Kobresia meadow. In Chinese Kobresia Meadows; Zhou, X.M., Ed.; China Science Press: Beijing, China, 2001; pp. 58–147. [Google Scholar]
- Zhou, X. Chinese Kobresia Meadows; Science Press: Beijing, China, 2001. [Google Scholar]
- Lal, R. Soil erosion impact on agronomic productivity and environment quality. Crit. Rev. Plant Sci. 1998, 17, 319–464. [Google Scholar] [CrossRef]
- Zhang, N.; Guo, R.; Song, P.; Guo, J.; Gao, Y. Effects of warming and nitrogen deposition on the coupling mechanism between soil nitrogen and phosphorus in Songnen Meadow Steppe, Northeastern China. Soil Biol. Biochem. 2013, 65, 96–104. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Y.; Zhang, C.; Niu, S.; Yang, H.; Yu, G.; Wang, H.; Blagodatskaya, E.; Kuzyakov, Y.; Tian, D.; et al. Contrasting responses of phosphatase kinetic parameters to nitrogen and phosphorus additions in forest soils. Funct. Ecol. 2018, 32, 106–116. [Google Scholar] [CrossRef]
- Gao, Q.; Guo, Y.; Xu, H.; Ganjurjav, H.; Li, Y.; Wan, Y.; Qin, X.; Ma, X.; Liu, S. Climate change and its impacts on vegetation distribution and net primary productivity of the alpine ecosystem in the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 554, 34–41. [Google Scholar] [CrossRef]
- Gao, Q.; Li, Y.; Wan, Y. Grassland degradation in Northern Tibet based on remote sensing data. J. Geogr. Sci. 2006, 16, 165–173. [Google Scholar] [CrossRef]
- Shi, P.; Sun, X.; Xu, L.; Zhang, X.; He, Y.; Zhang, D.; Yu, G. Net ecosystem CO2 exchange and controlling factors in a steppe-Kobresia meadow on the Tibetan Plateau. Sci. China Ser. D Earth Sci. 2006, 49, 207–218. [Google Scholar] [CrossRef]
- Zong, N.; Shi, P.; Song, M.; Zhang, X.; Jiang, J.; Chai, X. Nitrogen critical loads for an alpine meadow ecosystem on the Tibetan Plateau. Environ. Manag. 2016, 57, 531–542. [Google Scholar] [CrossRef]
- Zong, N.; Song, M.; Shi, P.; Jiang, J.; Zhang, X.; Shen, Z. Timing patterns of nitrogen application alter plant production and CO2 efflux in an alpine meadow on the Tibetan Plateau, China. Pedobiologia 2014, 57, 263–269. [Google Scholar] [CrossRef]
- Guo, W.; Qiangba, D. The eco-environment conditions and restoration measures of alpine grasslands in Damxuang County, Tibet. Tibet Sci. Technol. 2000, 21, 70–71. [Google Scholar]
- Jiang, J.; Shi, P.; Zong, N.; Fu, G.; Shen, Z.; Zhang, X.; Song, M. Climatic patterns modulate ecosystem and soil respiration responses to fertilization in an alpine meadow on the Tibetan Plateau, China. Ecol. Res. 2015, 30, 3–13. [Google Scholar] [CrossRef]
- Zhang, B. The Changes of Community Characteristics, Biomass Allocation and Soil Nutrient during Degradation and Recovery Process of Alpine Meadow in North Tibet. Master’s Thesis, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing, China, 2009. [Google Scholar]
- Wilsey, B.J.; Chalcraft, D.R.; Bowles, C.M.; Willig, M.R. Relationships among indices suggest that richness is an incomplete surrogate for grassland biodiversity. Ecology 2005, 86, 1178–1184. [Google Scholar] [CrossRef]
- Li, Y.; Dong, S.; Liu, S.; Su, X.; Wang, X.; Zhang, Y.; Zhao, Z.; Gao, X.; Li, S.; Tang, L. Relationships between plant diversity and biomass production of alpine grasslands are dependent on the spatial scale and the dimension of biodiversity. Ecol. Eng. 2019, 127, 375–382. [Google Scholar] [CrossRef]
- Wei, Z.; Yang, J.; Yang, S. Studies on community stability under different grazing systems on desert steppe. J. Soil Water Conserv. 2003, 17, 121–124. [Google Scholar]
- Lehman, C.L.; Tilman, D. Biodiversity, stability, and productivity in competitive communities. Am. Nat. 2000, 156, 534–552. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, F.; Ma, Y.; Niu, C. Study on status quo of grassland degradation and reason in source region of the Three Rivers. Sci. Technol. Qinghai Agric. For. 2007, 4, 29–32. [Google Scholar]
- Lü, X.; Reed, S.; Yu, Q.; He, N.; Wang, Z.; Han, X. Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland. Glob. Chang. Biol. 2013, 19, 2775–2784. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M.; Jackson, R.D. Plant nitrogen and phosphorus limitation in 98 North American grassland soils. Plant Soil 2010, 334, 73–84. [Google Scholar] [CrossRef]
- Djigal, D.; Brauman, A.; Diop, T.A.; Chotte, J.L.; Villenave, C. Influence of bacterial-feeding nematodes (Cephalobidae) on soil microbial communities during maize growth. Soil Biol. Biochem. 2004, 36, 323–331. [Google Scholar] [CrossRef]
- Li, C.; Li, Q.; Zhao, L.; Zhao, X. Responses of plant community biomass to nitrogen and phosphorus additions in natural and restored grasslands around Qinghai Lake Basin. Acta Phytoecol. Sin. 2016, 40, 1015–1027. [Google Scholar]
- Gao, Y.; Cooper, D.J.; Ma, X. Phosphorus additions have no impact on plant biomass or soil nitrogen in an alpine meadow on the Qinghai-Tibetan Plateau, China. Appl. Soil Ecol. 2016, 106, 18–23. [Google Scholar] [CrossRef]
- Xi, Y. Effects of Nitrogen, Phosphorus and Potassium Fertilizer on Ecosystem Productivity of an Alpine Meadow in Northern Tibet. Ph.D. Thesis, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing, China, 2015. [Google Scholar]
- Tang, Y.; Zhang, X.; Li, D.; Wang, H.; Chen, F.; Fu, X.; Fang, X.; Sun, X.; Yu, G. Impacts of nitrogen and phosphorus additions on the abundance and community structure of ammonia oxidizers and denitrifying bacteria in Chinese fir plantations. Soil Biol. Biochem. 2016, 103, 284–293. [Google Scholar]
- Rowe, E.C.; Smart, S.M.; Kennedy, V.H.; Emmett, B.A.; Evans, C.D. Nitrogen deposition increases the acquisition of phosphorus and potassium by heather Calluna vulgaris. Environ. Pollut. 2008, 155, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liu, X.; Xiao, M.; Li, Z. Effects of reseeding rate and rhizobia application on early seedling establishment of reseeding Medicago sativa. J. Jilin Agric. Sci. 2014, 39, 58–61. [Google Scholar]
- Liu, G.; He, F.; Wan, L.; Li, X. Management regimen and seeding rate modify seedling establishment of Leymus chinensis. Rangel. Ecol. Manag. 2015, 68, 204–210. [Google Scholar] [CrossRef]
- Sun, L.; Wang, X.; Wei, X.; Yang, X.; Wu, G. Effects of restoration measures on vegetation features of alpine degraded grassland of Amdo County in Tibet. Acta Agrestia Sin. 2012, 20, 616–620. [Google Scholar]
- Shang, Z.; Deng, B.; Ding, L.; Ren, G.; Xin, G.; Liu, Z.; Wang, Y.; Long, R. The effects of three years of fencing enclosure on soil seed banks and the relationship with above-ground vegetation of degraded alpine grasslands of the Tibetan plateau. Plant Soil 2013, 364, 229–244. [Google Scholar]
- Wu, G.; Li, W.; Zhao, L.; Shi, Z. Artificial management improves soil moisture, C, N and P in an alpine sandy meadow of Western China. Pedosphere 2011, 21, 407–412. [Google Scholar] [CrossRef]
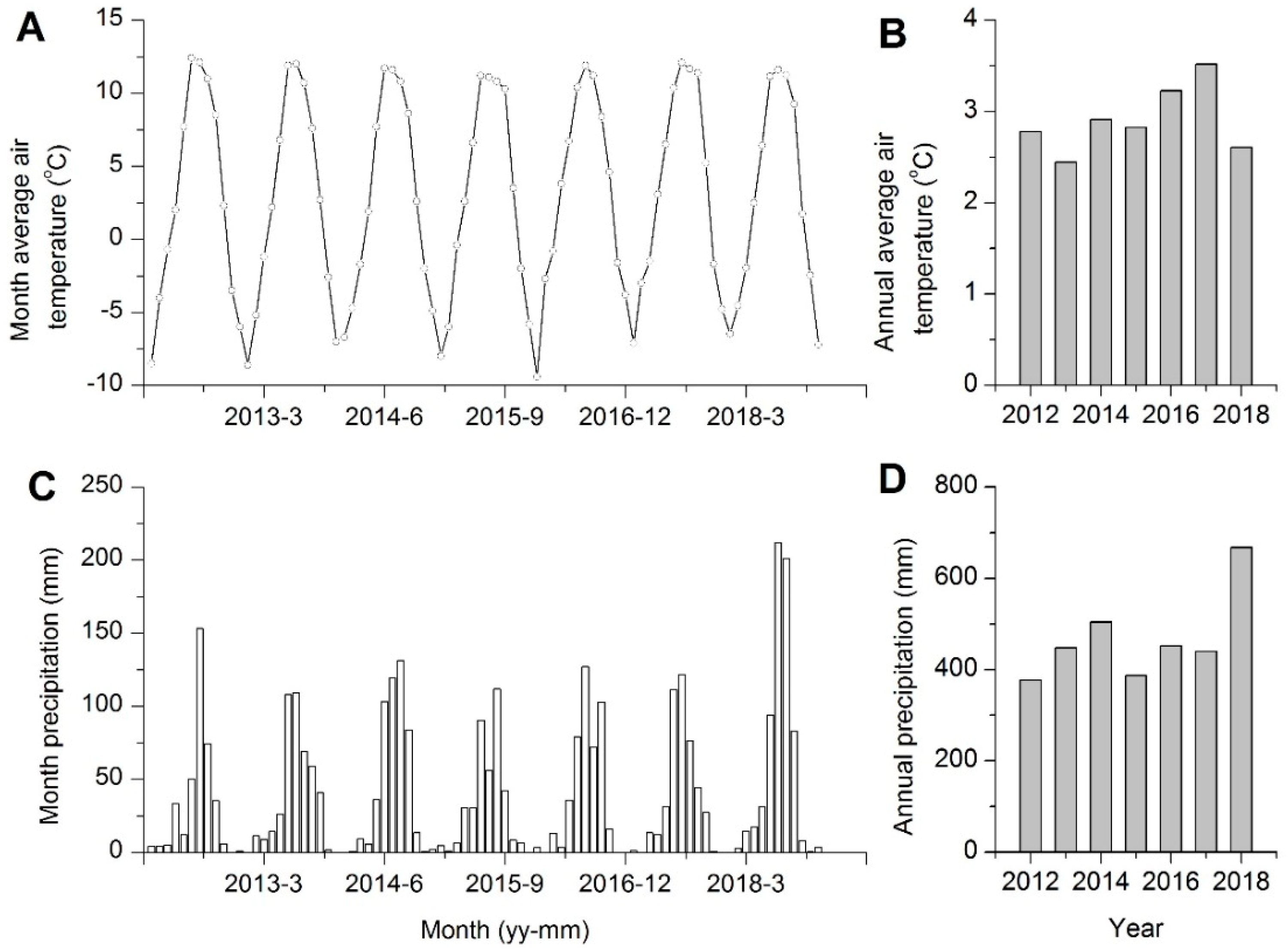
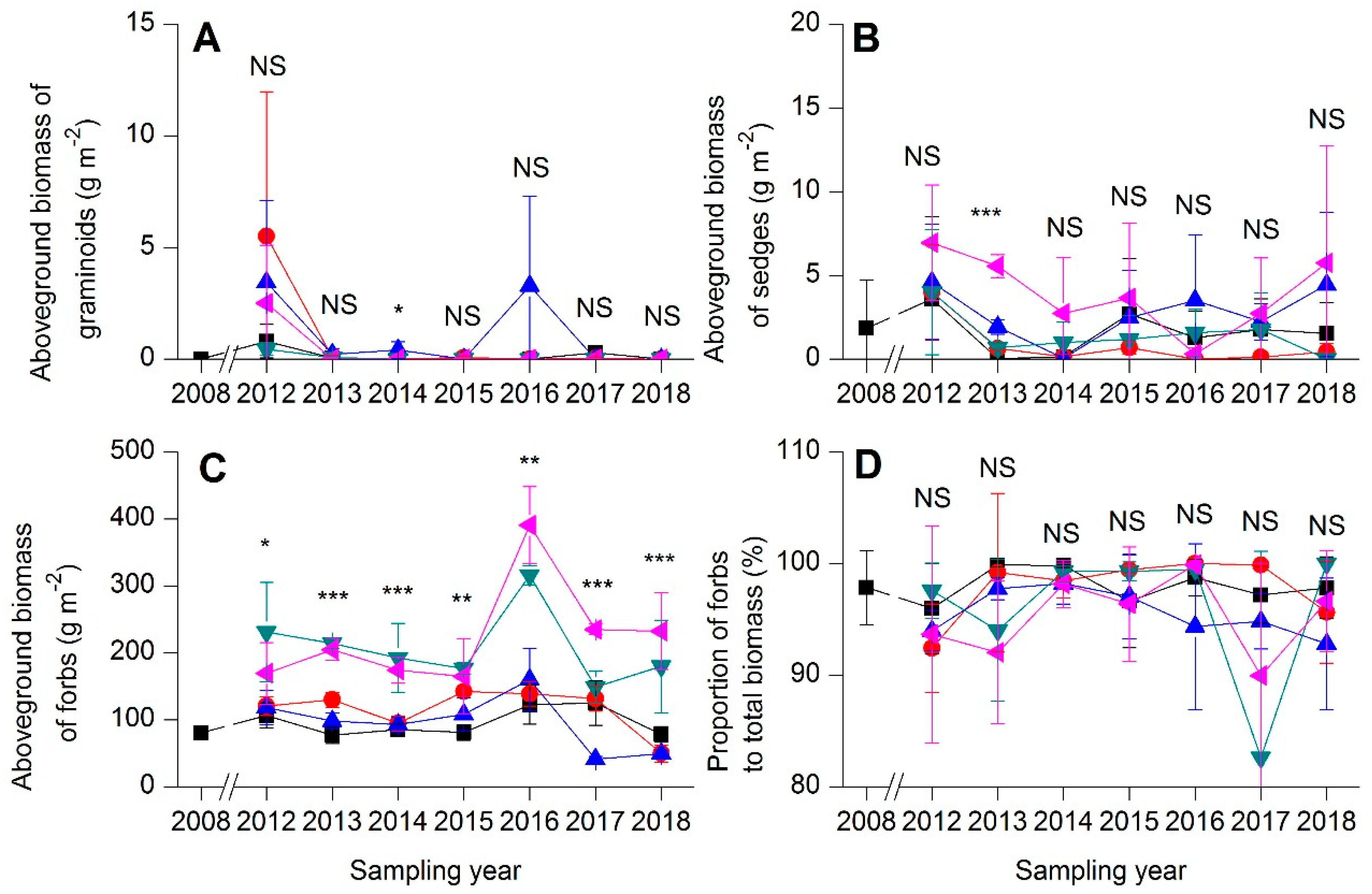
 ).
).
).
).

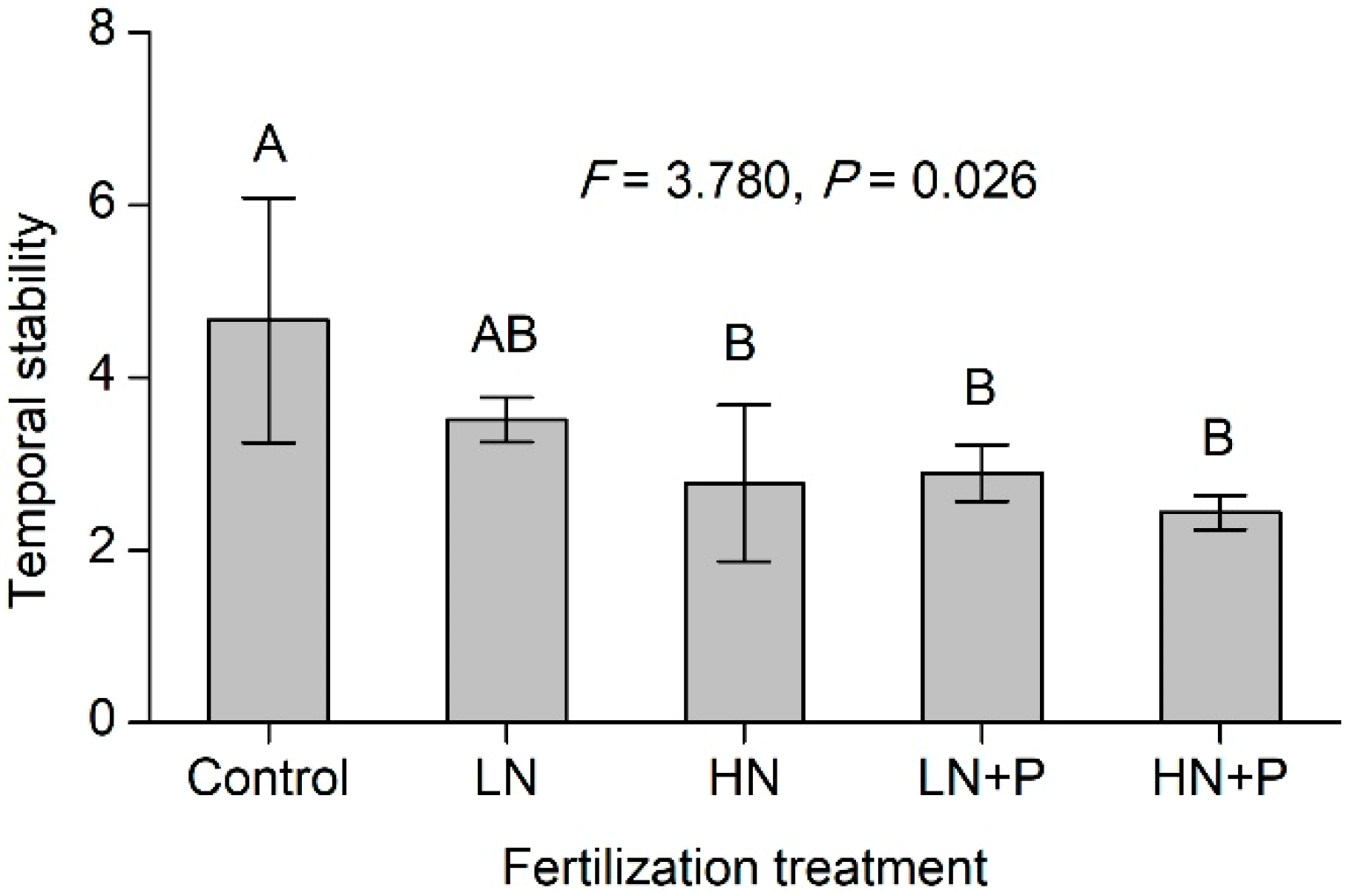 ).
).
).
).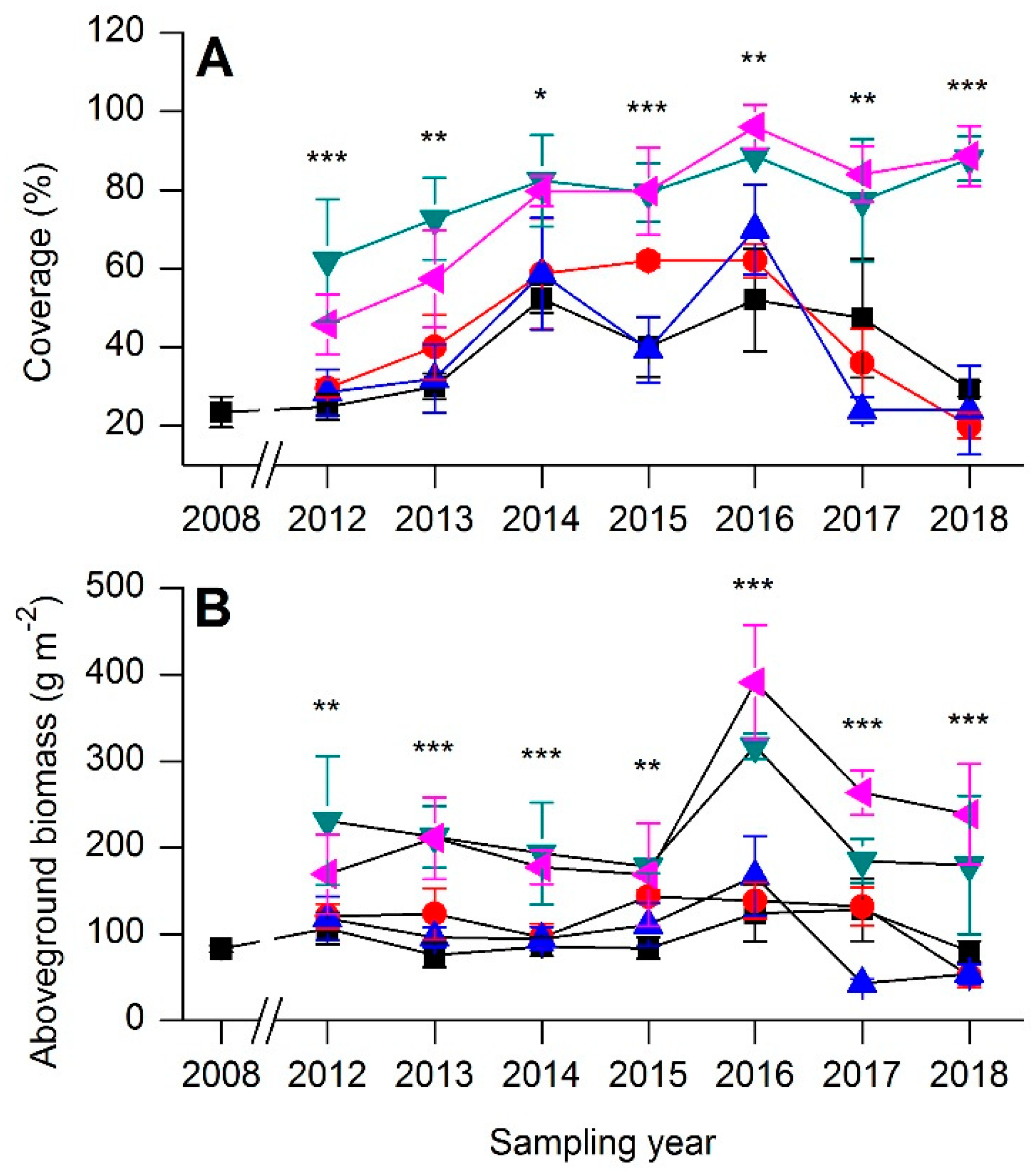 ).
).
).
).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year (Y) | Fertilization (F) | Y × F | ||||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| Species number | 14.828 | <0.001 | 2.199 | 0.118 | 1.511 | 0.145 |
| Simpson index | 11.216 | <0.001 | 7.210 | 0.002 | 3.305 | 0.001 |
| Shannon-Weiner index | 12.471 | <0.001 | 8.796 | 0.001 | 2.979 | 0.002 |
| Pielou index | 6.185 | <0.001 | 5.437 | 0.007 | 3.093 | 0.001 |
| Community coverage | 35.382 | <0.001 | 54.260 | <0.001 | 5.455 | <0.001 |
| Community aboveground biomass | 25.563 | <0.001 | 45.254 | <0.001 | 5.844 | <0.001 |
| Aboveground biomass of grasses | 5.676 | 0.016 | 1.809 | 0.180 | 1.134 | 0.375 |
| Aboveground biomass of sedges | 2.689 | 0.066 | 1.528 | 0.244 | 0.490 | 0.841 |
| Aboveground biomass of forbs | 24.900 | <0.001 | 36.998 | <0.001 | 5.839 | <0.001 |
| Proportion of forbs to total biomass | 2.742 | 0.017 | 0.907 | 0.485 | 1.333 | 0.167 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zong, N.; Shi, P. Enhanced Community Production rather than Structure Improvement under Nitrogen and Phosphorus Addition in Severely Degraded Alpine Meadows. Sustainability 2019, 11, 2023. https://doi.org/10.3390/su11072023
Zong N, Shi P. Enhanced Community Production rather than Structure Improvement under Nitrogen and Phosphorus Addition in Severely Degraded Alpine Meadows. Sustainability. 2019; 11(7):2023. https://doi.org/10.3390/su11072023
Chicago/Turabian StyleZong, Ning, and Peili Shi. 2019. "Enhanced Community Production rather than Structure Improvement under Nitrogen and Phosphorus Addition in Severely Degraded Alpine Meadows" Sustainability 11, no. 7: 2023. https://doi.org/10.3390/su11072023
APA StyleZong, N., & Shi, P. (2019). Enhanced Community Production rather than Structure Improvement under Nitrogen and Phosphorus Addition in Severely Degraded Alpine Meadows. Sustainability, 11(7), 2023. https://doi.org/10.3390/su11072023