Characterization of Di-n-Butyl Phthalate Phytoremediation by Garden Lettuce (Lactuca sativa L. var. longifolia) through Kinetics and Proteome Analysis



Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cultivation of Plants
2.3. Experimental Design
2.4. Plant Leaf Chlorophyll (a + b) Concentration Determination
2.5. DBP Concentration Analyses
2.6. Statistical Analyses
2.7. Algal Biotoxicity Assays
2.8. Proteomics Analysis of the Selected Plant
2.8.1. Protein Extraction
2.8.2. Two-Dimensional Gel Electrophoresis (2-DE)
2.8.3. Digestion of In-Gel Protein
2.8.4. Proteomic Mass Spectrometry Analyses
3. Results and Discussion
3.1. Evaluation of DBP Phytoremediation Capability in Eleven Test Plants
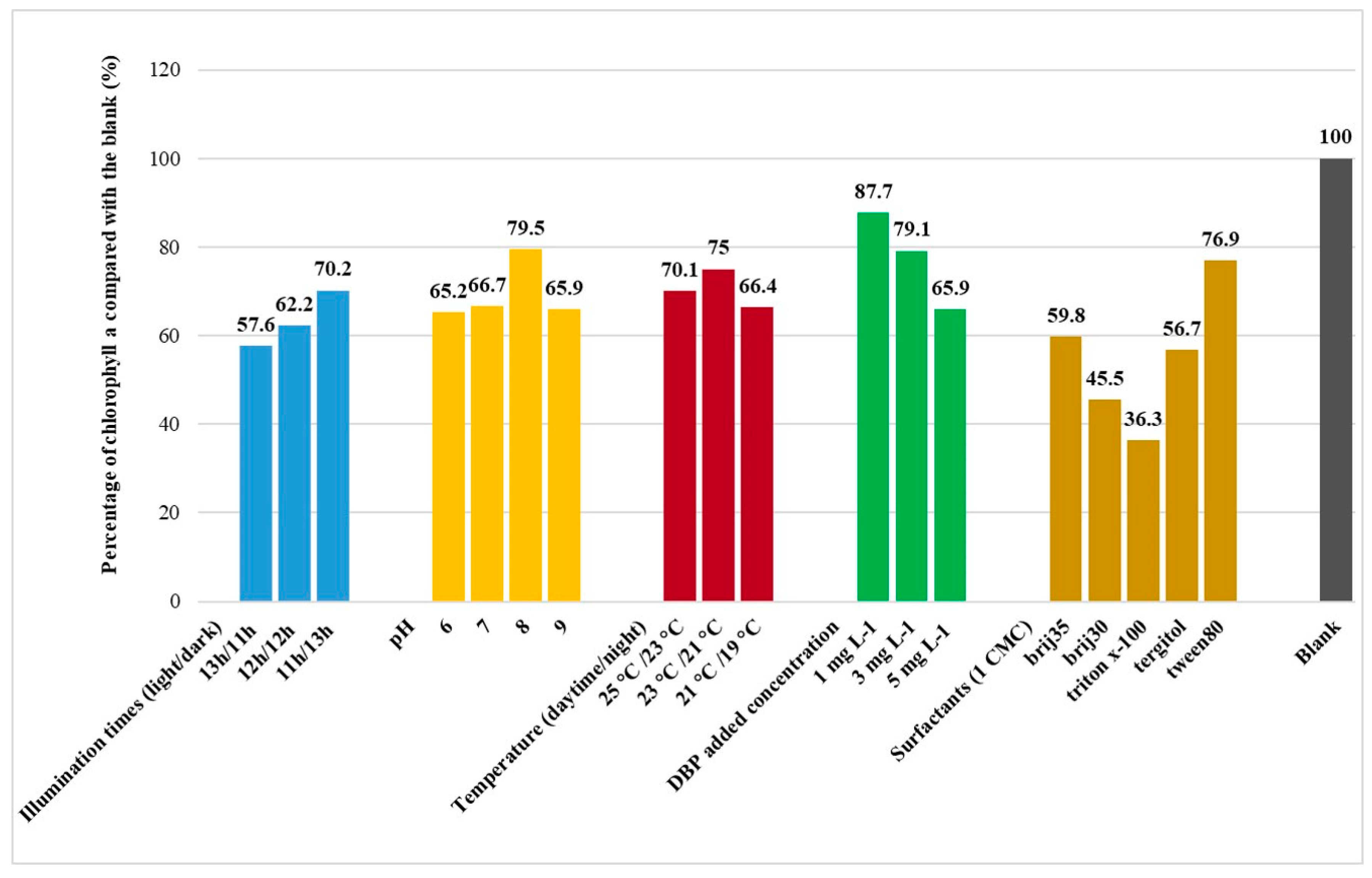
3.2. Cultural Conditions of DBP Phytoremediation
3.3. Algal Biotoxicity of DBP
3.4. Proteomic Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abdel Daiem, M.M.; Rivera-Utrilla, J.; Ocampo-Pérez, R.; Méndez-Díaz, J.D.; Sánchez-Polo, M. Environmental impact of phthalic acid esters and their removal from water and sediments by different technologies-A review. J. Environ. Manag. 2012, 109, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Feng, L.; Li, X. Pathway of diethyl phthalate photolysis in sea-water determined by gas chromatography-mass spectrometry and compound-specific isotope analysis. Chemosphere 2013, 90, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, P.; Wang, L.; Sun, G.; Zhao, J.; Zhang, H.; Du, N. The influence of facility agriculture production on phthalate esters distribution in black soils of northeast China. Sci. Total Environ. 2015, 506–507, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Kaewlaoyoong, A.; Vu, C.T.; Lin, C.; Liao, C.S.; Chen, J.R. Occurrence of phthalate esters around the major plastic industrial area in southern Taiwan. Environ. Earth Sci. 2018, 77, 457. [Google Scholar] [CrossRef]
- Ashworth, M.J.; Chappell, A.; Ashmore, E.; Fowles, J. Analysis and assessment of exposure to selected phthalates found in children’s toys in Christchurch, New Zealand. Int. J. Environ. Res. Public Health 2018, 15, 200. [Google Scholar] [CrossRef] [PubMed]
- Staples, C.A.; Peterson, D.R.; Parkerton, T.F.; Adams, W.J. The environmental fate of phthalate esters: A literature review. Chemosphere 1997, 35, 667–749. [Google Scholar] [CrossRef]
- Peijnenburg, W.J.G.M.; Struijs, J. Occurrence of phthalate esters in the environment of the Netherlands. Ecotoxicol. Environ. Saf. 2006, 63, 204–215. [Google Scholar] [CrossRef]
- Meng, X.Z.; Wang, Y.; Xiang, N.; Chen, L.; Liu, Z.; Wu, B.; Dai, X.; Zhang, Y.H.; Xie, Z.; Ebinghaus, R. Flow of sewage sludge-borne phthalate esters (PAEs) from human release to human intake: Implication for risk assessment of sludge applied to soil. Sci. Total Environ. 2014, 476–477, 242–249. [Google Scholar] [CrossRef]
- Net, S.; Rabodonirina, S.; Sghaier, R.B.; Dumoulin, D.; Chbib, C.; Tlili, I.; Ouddane, B. Distribution of phthalates, pesticides and drug residues in the dissolved, particulate and sedimentary phases from transboundary rivers (France–Belgium). Sci. Total Environ. 2015, 521–522, 152–159. [Google Scholar] [CrossRef]
- Net, S.; Sempéré, R.; Delmont, A.; Paluselli, A.; Ouddane, B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ. Sci. Technol. 2015, 49, 4019–4035. [Google Scholar] [CrossRef]
- Li, J.; Wang, G.; Aggarwal, S.G.; Huang, Y.; Ren, Y.; Zhou, B.; Singh, K.; Gupta, P.K.; Cao, J.; Zhang, R. Comparison of abundances, compositions and sources of elements, inorganic ions and organic compounds in atmospheric aerosols from Xi’an and New Delhi, two megacities in China and India. Sci. Total Environ. 2014, 476–477, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.Y.; Liu, C.; Liao, C.S.; Chang, B.V. Occurrence and microbial degradation of phthalate esters in Taiwan river sediments. Chemosphere 2002, 49, 1295–1299. [Google Scholar] [CrossRef]
- Selvaraj, K.K.; Sundaramoorthy, G.; Ravichandran, P.K.; Girijan, G.K.; Sampath, S.; Ramaswamy, B.R. Phthalate esters in water and sediments of the Kaveri River, India: Environmental levels and ecotoxicological evaluations. Environ. Geochem. Health 2015, 37, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhang, B.T.; Teng, Y. Distribution of phthalate acid esters in lakes of Beijing and its relationship with anthropogenic activities. Sci. Total Environ. 2014, 476–477, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.F.; Ju, Y.R.; Lim, Y.; Chang, J.H.; Chen, C.W.; Dong, C.D. Spatial and temporal distribution of di-(2-ethylhexyl) phthalate in urban river sediments. Int. J. Environ. Res. Public Health 2018, 15, 2228. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Bolaños, P.; Padilla-Sánchez, J.A.; Garrido-Frenich, A.; Romero-González, R.; Martínez-Vidal, J.L. Evaluation of soil contamination in intensive agricultural areas by pesticides and organic pollutants: South-eastern Spain as a case study. J. Environ. Monit. 2012, 14, 1182. [Google Scholar] [CrossRef]
- Ma, T.T.; Christie, P.; Luo, Y.M.; Teng, Y. Phthalate esters contamination in soil and plants on agricultural land near an electronic waste recycling site. Environ. Geochem. Health 2013, 35, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Škrbić, B.D.; Ji, Y.; Đurišić-Mladenović, N.; Zhao, J. Occurence of the phthalate esters in soil and street dust samples from the Novi Sad city area, Serbia, and the influence on the children’s and adults’ exposure. J. Hazard. Mater. 2016, 312, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.E.; Ostby, J.; Furr, J.; Price, M.; Veeramachaneni, D.N.R.; Parks, L. Perinatal exposure to the phthalates DEHP, BBP, and DINP, but not DEP, DMP, or DOTP, alters sexual differentiation of the male rat. Toxicol. Sci. 2000, 58, 350–365. [Google Scholar] [CrossRef]
- Saillenfait, A.M.; Sabaté, J.P.; Gallissot, F. Developmental toxic effects of diisobutyl phthalate, the methyl-branched analogue of di-n-butyl phthalate, administered by gavage to rats. Toxicol. Lett. 2006, 165, 39–46. [Google Scholar] [CrossRef]
- McKee, R.H.; Butala, J.H.; David, R.M.; Gans, G. NTP center for the evaluation of risks to human reproduction reports on phthalates: Addressing the data gaps. Reprod. Toxicol. 2004, 18, 1–22. [Google Scholar] [CrossRef]
- Hansen, J.F.; Bendtzen, K.; Boas, M.; Frederiksen, H.; Nielsen, C.H.; Rasmussen, Å.K.; Feldt-Rasmussen, U. Influence of phthalates on cytokine production in monocytes and macrophages: A systematic review of experimental trials. PLoS ONE 2015, 10, e0120083. [Google Scholar] [CrossRef]
- Bui, T.T.; Giovanoulis, G.; Cousins, A.P.; Magnér, J.; Cousins, I.T.; deWit, C.A. Human exposure, hazard and risk of alternative plasticizers to phthalate esters. Sci. Total Environ. 2016, 541, 451–467. [Google Scholar] [CrossRef]
- Chemical Manufacturers Association. Comments of the Chemical Manufacturers Association Phthalate Esters Panel in Response to Request for Public Input on Seven Phthalate Esters; Chemical Manufacturers Association: Washington DC, USA, 1999. [Google Scholar]
- Wolfe, N.L.; Steen, W.C.; Burns, L.A. Phthalate ester hydrolysis: Linear free energy relationships. Chemosphere 1980, 9, 403–408. [Google Scholar] [CrossRef]
- Gao, D.W.; Wen, Z.D. Phthalate esters in the environment: A critical review of their occurrence, biodegradation, and removal during wastewater treatment processes. Sci. Total Environ. 2016, 541, 986–1001. [Google Scholar] [CrossRef]
- Long, G.; Meek, E. Environmental Health Criteria 189: Di-n-Butyl Phthalate; World Heal. Organ: Geneva, Switzerland, 1997. [Google Scholar]
- Chan, P.K.L.; Meek, M.E. Di-n-Butyl phthalate: Evaluation of risks to health from environmental exposure in Canada. J. Environ. Sci. Heal. Part C 1994, 12, 257–268. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services. Toxicological Profile for di-n-butyl phthalate. In Public Health Service; Agency for Toxic Substances and Disease Registry: Atlanta, Georgia, USA, 1990. [Google Scholar]
- Rudel, R.A.; Camann, D.E.; Spengler, J.D.; Korn, L.R.; Brody, J.G. Phthalates, alkylphenols, pesticides, polybrominated diphenyl ethers, and other endocrine-disrupting compounds in indoor air and dust. Environ. Sci. Technol. 2003, 37, 4543–4553. [Google Scholar] [CrossRef]
- Duan, J.; Kang, J.; Deng, T.; Yang, X.; Chen, M. Exposure to DBP and high iodine aggravates autoimmune thyroid disease through increasing the levels of IL-17 and thyroid-binding globulin in Wistar rats. Toxicol. Sci. 2018, 163, 196–205. [Google Scholar] [CrossRef]
- Hemming, I.V.; Ann-Marie, H.; Johan, M. Effects of di-n-butyl phthalate on the carotenoid synthesis in green plants. Physiol. Plant. 1981, 53, 158–163. [Google Scholar]
- Herring, R.; Bering, C.L. Effects of phthalate esters on plant seedlings and reversal by a soil microorganism. Bull. Environ. Contam. Toxicol. 1988, 40, 626–632. [Google Scholar] [CrossRef]
- Hardwick, R.C.; Cole, R.A.; Fyfield, T.P. Injury to and death of cabbage (Brassica oleracea) seedlings caused by vapours of di butyl phthalate emitted from certain plastics. Ann. Appl. Biol. 1984, 105, 97–105. [Google Scholar] [CrossRef]
- Hannay, J.W.; Millar, D.J. Phytotoxicity of phthalate plasticisers: 1. Diagnosis and commercial implications. J. Exp. Bot. 1986, 37, 883–897. [Google Scholar] [CrossRef]
- Yin, R.; Lin, X.G.; Wang, S.G.; Zhang, H.Y. Effect of DBP/DEHP in vegetable planted soil on the quality of capsicum fruit. Chemosphere 2003, 50, 801–805. [Google Scholar] [CrossRef]
- Dueck, T.A.; VanDijk, C.J.; David, F.; Scholz, N.; Vanwalleghem, F. Chronic effects of vapour phase di-n-butyl phthalate (DBP) on six plant species. Chemosphere 2003, 53, 911–920. [Google Scholar] [CrossRef]
- Li, C.; Chen, J.; Wang, J.; Han, P.; Luan, Y.; Ma, X.; Lu, A. Phthalate esters in soil, plastic film, and vegetable from greenhouse vegetable production bases in Beijing, China: Concentrations, sources, and risk assessment. Sci. Total Environ. 2016, 568, 1037–1043. [Google Scholar] [CrossRef]
- Ma, T.; Zhou, W.; Chen, L.; Wu, L.; Christie, P.; Liu, W. Toxicity of phthalate esters to lettuce (Lactuca sativa) and the soil microbial community under different soil conditions. PLoS ONE 2018, 13, e0208111. [Google Scholar] [CrossRef]
- Parker, R.; Flowers, T.J.; Moore, A.L.; Harpham, N.V.J. An accurate and reproducible method for proteome profiling of the effects of salt stress in the rice leaf lamina. J. Exp. Bot. 2006, 57, 1109–1118. [Google Scholar] [CrossRef]
- Aghaei, K.; Ehsanpour, A.A.; Komatsu, S. Proteome Analysis of Potato under Salt Stress. J. Proteome Res. 2008, 7, 4858–4868. [Google Scholar] [CrossRef]
- Liao, C.S.; Yen, J.H.; Wang, Y.S. Effects of endocrine disruptor di-n-butyl phthalate on the growth of Bok choy (Brassica rapa subsp. chinensis). Chemosphere 2006, 65, 1715–1722. [Google Scholar] [CrossRef]
- Liao, C.S.; Yen, J.H.; Wang, Y.S. Growth inhibition in Chinese cabbage (Brassica rapa var. chinensis) growth exposed to di-n-butyl phthalate. J. Hazard. Mater. 2009, 163, 625–631. [Google Scholar] [CrossRef]
- He, L.; Gielen, G.; Bolan, N.S.; Zhang, X.; Qin, H.; Huang, H.; Wang, H. Contamination and remediation of phthalic acid esters in agricultural soils in China: A review. Agron. Sustain. Dev. 2015, 35, 519–534. [Google Scholar] [CrossRef]
- Cai, Q.Y.; Mo, C.H.; Wu, T.; Zeng, Q.Y. Accumulation of phthalic acid esters in water spinach (Ipomoea aquatica) and in Paddy soil. Bull. Environ. Contam. Toxicol. 2006, 77, 411–418. [Google Scholar] [CrossRef]
- Cai, Q.Y.; Mo, C.H.; Zeng, Q.Y.; Wu, Q.T.; Férard, J.F.; Antizar-Ladislao, B. Potential of Ipomoea aquatica cultivars in phytoremediation of soils contaminated with di-n-butyl phthalate. Environ. Exp. Bot. 2008, 62, 205–211. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Chai, W.; Wei, J.; Wang, Q.; Li, B.; Li, H. The use of proteomic analysis for exploring the phytoremediation mechanism of Scirpus triqueter to pyrene. J. Hazard. Mater. 2013, 260, 1001–1007. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 22. [Google Scholar]
- Liao, C.S.; Chen, L.C.; Chen, B.S.; Lin, S.H. Bioremediation of endocrine disruptor di-n-butyl phthalate ester by Deinococcus radiodurans and Pseudomonas stutzeri. Chemosphere 2010, 78, 342–346. [Google Scholar] [CrossRef]
- Chang, R.; Jien, S.H.; Weng, C.H.; Lee, T.W.; Liao, C.S. Fast removal of polybrominated diphenyl ethers from aqueous solutions by using low-cost adsorbents. Sustainability 2017, 9, 102. [Google Scholar] [CrossRef]
- Hochstrasser, D.F.; Harrington, M.G.; Hochstrasser, A.C.; Miller, M.J.; Merril, C.R. Methods for increasing the resolution of two-dimensional protein electrophoresis. Anal. Biochem. 1988, 173, 424–435. [Google Scholar] [CrossRef]
- Hellman, U.; Wernstedt, C.; Gonez, J.; Heldin, C.-H. Improvement of an “In-Gel” digestion procedure for the micropreparation of internal protein fragments for amino acid sequencing. Anal. Biochem. 1995, 224, 451–455. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.C.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Chang, B.V.; Liao, C.S.; Yuan, S.Y. Anaerobic degradation of diethyl phthalate, di-n-butyl phthalate, and di-(2-ethylhexyl) phthalate from river sediment in Taiwan. Chemosphere 2005, 58, 1601–1607. [Google Scholar] [CrossRef]
- Gu, S.; Zheng, H.; Xu, Q.; Sun, C.; Shi, M.; Wang, Z.; Li, F. Comparative toxicity of the plasticizer dibutyl phthalate to two freshwater algae. Aquat. Toxicol. 2017, 191, 122–130. [Google Scholar] [CrossRef]
- Heazlewood, J.L.; Jorrín-Novo, J.V.; Agrawal, G.K.; Mazzuca, S.; Lüthje, S. Editorial: International Plant Proteomics Organization (INPPO) world congress 2014. Front. Plant Sci. 2016, 7, 1190. [Google Scholar] [CrossRef]
- Pasqualato, S.; Renault, L.; Cherfils, J. Arf, Arl, Arp and Sar proteins: A family of GTP-binding proteins with a structural device for ‘front–back’ communication. EMBO Rep. 2002, 3, 1035–1041. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Biomass (g, dry wt) a | Chlorophyll Concentration (mg g−1, fresh wt) a | Residual DBP in Hydroponic Solution (mg L−1) a | |||
|---|---|---|---|---|---|---|
| 0 day | 21 days | 0 day | 21 days | 0 day | 21 days | |
| Edible rape | 2.64 ± 0.68 | 4.32 ± 0.52 | 0.227 ± 0.032 | 0.259 ± 0.048 | 5.02 ± 0.15 | 3.89 ± 0.18 |
| Control c | 2.83 ± 0.58 | 5.63 ± 0.82 | 0.264 ± 0.034 | 0.306 ± 0.066 | ||
| Chinese cabbage | 2.82 ± 0.95 | 6.58 ± 1.52 | 0.132 ± 0.028 | 0.194 ± 0.042 | 5.02 ± 0.16 | 3.56 ± 0.13 |
| Control c | 2.74 ± 0.84 | 7.08 ± 1.38 | 0.144 ± 0.032 | 0.226 ± 0.064 | ||
| Spinach | 2.35 ± 0.76 | 3.28 ± 0.85 | 0.253 ± 0.044 | 0.268 ± 0.048 | 4.98 ± 0.14 | 4.72 ± 0.15 |
| Control c | 2.39 ± 0.54 | 2.98 ± 0.78 | 0.244 ± 0.028 | 0.318 ± 0.054 | ||
| Chinese mustard | 1.48 ± 0.36 | 4.63 ± 1.46 | 0.154 ± 0.035 | 0.204 ± 0.047 | 4.99 ± 0.16 | 4.79 ± 0.16 |
| Control c | 1.52 ± 0.26 | 5.79 ± 1.24 | 0.156 ± 0.028 | 0.417 ± 0.068 | ||
| Water cabbage | 2.68 ± 0.28 | 7.18 ± 1.44 | 0.252 ± 0.046 | 0.309 ± 0.063 | 5.06 ± 0.21 | 2.24 ± 0.08 |
| Control c | 2.57 ± 0.23 | 6.92 ± 0.88 | 0.264 ± 0.058 | 0.364 ± 0.088 | ||
| Ceylon spinach | 1.12 ± 0.25 | 3.76 ± 0.88 | 0.279 ± 0.055 | 0.319 ± 0.058 | 4.95 ± 0.12 | 3.52 ± 0.14 |
| Control c | 1.15 ± 0.22 | 4.21 ± 0.82 | 0.282 ± 0.048 | 0.355 ± 0.098 | ||
| Garden lettuce | 1.45 ± 0.26 | 4.68 ± 0.52 | 0.165 ± 0.037 | 0.269 ± 0.042 | 4.99 ± 0.14 | n.d. b |
| Control c | 1.49 ± 0.31 | 4.72 ± 0.52 | 0.169 ± 0.037 | 0.272 ± 0.042 | ||
| Chinese celery | 0.92 ± 0.32 | 4.06 ± 0.85 | 0.141 ± 0.032 | 0.191 ± 0.038 | 5.03 ± 0.18 | 4.72 ± 0.18 |
| Control c | 1.02 ± 0.29 | 4.54 ± 0.92 | 0.144 ± 0.038 | 0.255 ± 0.049 | ||
| Edible amaranth | 1.54 ± 0.56 | 6.13 ± 1.22 | 0.153 ± 0.025 | 0.192 ± 0.035 | 5.05 ± 0.24 | 3.33 ± 0.16 |
| Control c | 1.47 ± 0.48 | 5.74 ± 0.89 | 0.149 ± 0.023 | 0.214 ± 0.042 | ||
| Cauliflower | 3.14 ± 0.85 | 9.42 ± 1.98 | 0.082 ± 0.016 | 0.129 ± 0.023 | 5.02 ± 0.05 | 2.16 ± 0.05 |
| Control c | 3.06 ± 0.62 | 9.68 ± 1.42 | 0.083 ± 0.016 | 0.133 ± 0.031 | ||
| Chinese chive | 0.65 ± 0.22 | 3.43 ± 0.34 | 0.203 ± 0.038 | 0.249 ± 0.046 | 4.99 ± 0.05 | 4.92 ± 0.06 |
| Control c | 0.66 ± 0.23 | 3.42 ± 0.29 | 0.205 ± 0.033 | 0.251 ± 0.044 | ||
| Blank d | 5.04 ± 0.12 | 4.96 ± 0.08 | ||||
| Common Name | DBP Accumulated Concentration (mg Kg−1) a | |||||
|---|---|---|---|---|---|---|
| Root | Stem | Leaf | ||||
| 0 day | 21 days | 0 day | 21 days | 0 day | 21 days | |
| Edible rape | n.d. b | 0.05 ± 0.01 | n.d. b | 0.16 ± 0.04 | n.d. b | 0.68 ± 0.06 |
| Chinese cabbage | n.d. b | 0.10 ± 0.02 | n.d. b | 0.22 ± 0.06 | n.d. b | 0.74 ± 0.12 |
| Spinach | n.d. b | 0.03 ± 0.01 | n.d. b | n.d. b | n.d. b | n.d. b |
| Chinese mustard | n.d. b | 0.06 ± 0.02 | n.d. b | n.d. b | n.d. b | n.d. b |
| Water cabbage | n.d. b | 0.43 ± 0.12 | n.d. b | 0.88 ± 0.14 | n.d. b | 0.82 ± 0.18 |
| Ceylon spinach | n.d. b | 0.08 ± 0.02 | n.d. b | 0.26 ± 0.04 | n.d. b | 0.46 ± 0.12 |
| Garden lettuce | n.d. b | 3.35 ± 0.42 | n.d. b | 2.74 ± 0.38 | n.d. b | 4.35 ± 0.42 |
| Chinese celery | n.d. b | n.d. b | n.d. b | n.d. b | n.d. b | n.d. b |
| Edible amaranth | n.d. b | 0.22 ± 0.04 | n.d. b | 0.12 ± 0.02 | n.d. b | 0.25 ± 0.06 |
| Cauliflower | n.d. b | 0.37 ± 0.02 | n.d. b | 0.45 ± 0.06 | n.d. b | 0.96 ± 0.08 |
| Chinese chive | n.d. b | n.d. b | n.d. b | n.d. b | n.d. b | n.d. b |
| Treatment | Garden Lettuce (Lactuca sativa L. var. longifolia.) | ||
|---|---|---|---|
| k1 (day−1) | t1/2 (days) | r2 | |
| Illumination times (light/dark) | |||
| 13h/11h | 1.295 | 10.362 | 0.811 |
| 12h/12h | 0.986 | 10.237 | 0.896 |
| 11h/13h | 2.283 | 5.299 | 0.896 |
| pH | |||
| 6 | 1.575 | 6.155 | 0.784 |
| 7 | 3.506 | 3.597 | 0.921 |
| 8 | 4.605 | 2.887 | 0.899 |
| 9 | 3.912 | 3.173 | 0.921 |
| Temperature (daytime/night) | |||
| 25 °C /23 °C | 3.616 | 3.265 | 0.942 |
| 23 °C /21 °C | 4.609 | 2.991 | 0.820 |
| 21 °C /19 °C | 2.477 | 4.834 | 0.926 |
| DBP added concentration | |||
| 1 mg L−1 | 4.741 | 2.504 | 0.866 |
| 3 mg L−1 | 4.273 | 2.840 | 0.905 |
| 5 mg L−1 | 3.783 | 3.193 | 0.931 |
| Surfactants (1 CMC) | |||
| Brij35 | 2.442 | 4.719 | 0.963 |
| Brij30 | 1.677 | 7.706 | 0.894 |
| Triton x-100 | 1.085 | 10.730 | 0.698 |
| Tergitol | 2.354 | 5.797 | 0.831 |
| Tween 80 | 4.605 | 2.686 | 0.937 |
| Spots | Protein Name | Accession No. | Estimated MW (kD) | Estimated pI | Amounts a |
|---|---|---|---|---|---|
| 1 | Photosystem II reaction center protein H | Q332U8 | 7.7 | 6.23 | - |
| 2 | Putative ADP-ribosylation factor | A8QVJ0 | 15.6 | 4.89 | - |
| 3 | Chloroplastic 30S ribosomal protein S7 | Q332R9 | 17.2 | 5.35 | - |
| 4 | Protection of telomeres 1 protein | B7T1J4 | 50.1 | 6.38 | - |
| 5 | NBS-LRR resistance-like protein RGC1F | Q56P11 | 51.8 | 7.95 | + |
| 6 | DNA-directed RNA polymerase subunit beta | Q91Fl1 | 80.4 | 7.51 | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, C.-S.; Nishikawa, Y.; Shih, Y.-T. Characterization of Di-n-Butyl Phthalate Phytoremediation by Garden Lettuce (Lactuca sativa L. var. longifolia) through Kinetics and Proteome Analysis. Sustainability 2019, 11, 1625. https://doi.org/10.3390/su11061625
Liao C-S, Nishikawa Y, Shih Y-T. Characterization of Di-n-Butyl Phthalate Phytoremediation by Garden Lettuce (Lactuca sativa L. var. longifolia) through Kinetics and Proteome Analysis. Sustainability. 2019; 11(6):1625. https://doi.org/10.3390/su11061625
Chicago/Turabian StyleLiao, Chien-Sen, Yoshikazu Nishikawa, and Yu-Ting Shih. 2019. "Characterization of Di-n-Butyl Phthalate Phytoremediation by Garden Lettuce (Lactuca sativa L. var. longifolia) through Kinetics and Proteome Analysis" Sustainability 11, no. 6: 1625. https://doi.org/10.3390/su11061625
APA StyleLiao, C.-S., Nishikawa, Y., & Shih, Y.-T. (2019). Characterization of Di-n-Butyl Phthalate Phytoremediation by Garden Lettuce (Lactuca sativa L. var. longifolia) through Kinetics and Proteome Analysis. Sustainability, 11(6), 1625. https://doi.org/10.3390/su11061625