Response of Corals Acropora pharaonis and Porites lutea to Changes in pH and Temperature in the Gulf


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
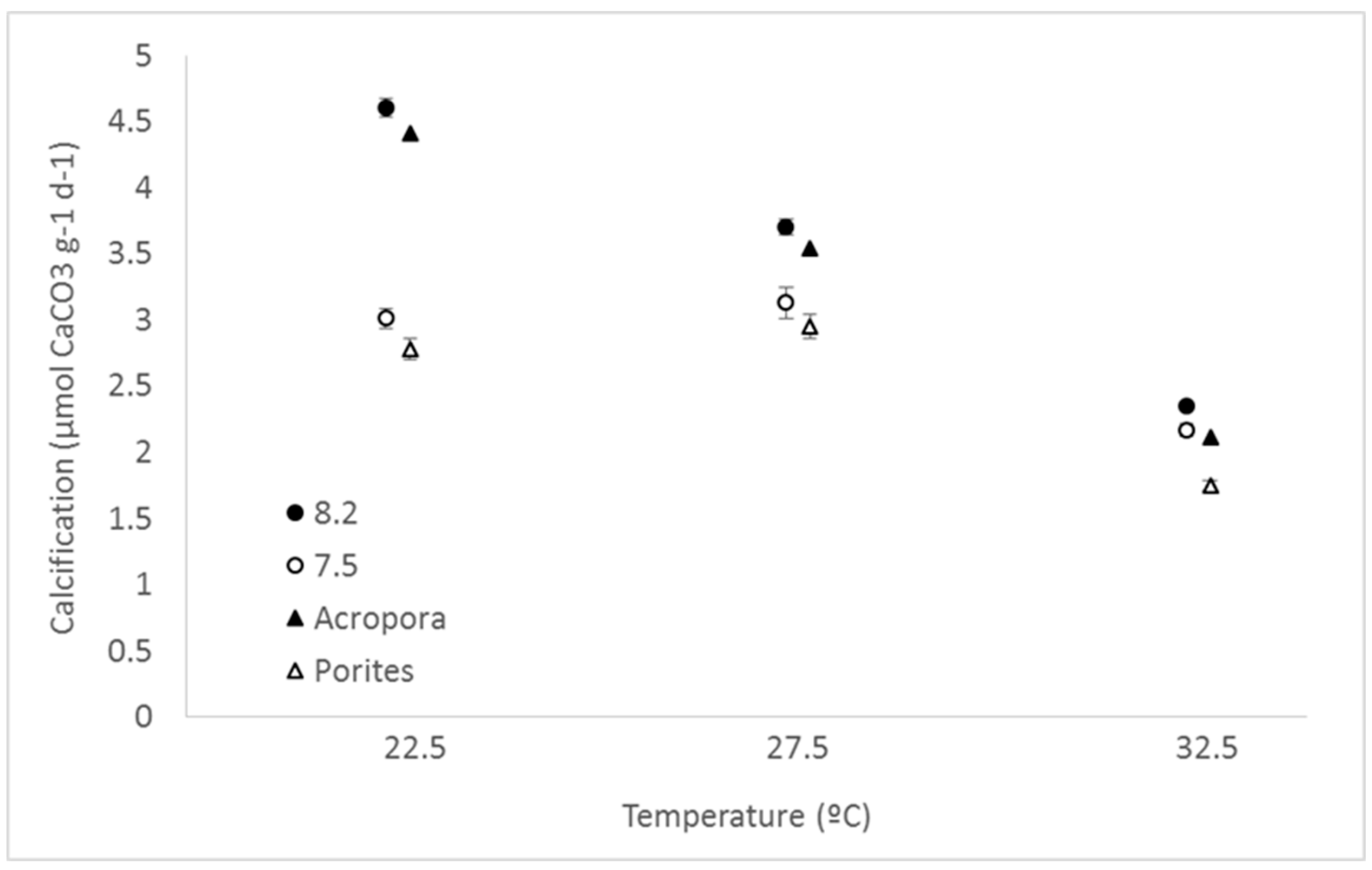
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Feary, D.A.; Burt, J.A.; Cavalcante, G.H.; Bauman, A.G. Extreme Physical Factors and the Structure of Gulf Fish and Reef Communities. In Coral Reefs of the Gulf; Riegl, B.M., Purkis, S.J., Eds.; Springer: New York, NY, USA, 2012; Volume 3, pp. 163–170. [Google Scholar]
- Bahr, K.; Jokiel, P.; Ku’ulei, S. Seasonal and annual calcification rates of the Hawaiian reef coral, Montipora capitata, under present and future climate change scenarios. ICES J. Mar. Sci. 2017, 74, 1083–1091. [Google Scholar]
- Schiedek, D.; Sundelin, B.; Readman, J.W.; Macdonald, R.W. Interactions between climate change and contaminants. Mar. Poll. Bull. 2007, 54, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- Uddin, S.; Al-Ghadban, A.N.; Al Khabbaz, A. Localized Hyper Saline Waters in Arabian Gulf from Desalination activity—An example from South Kuwait. Environ. Monit. Assess. 2010, 181, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Uddin, S.; Gevao, B.; Al-Ghadban, A.N.; Nithyanandan, M.; Al-Shamroukh, D. Acidification in Arabian Gulf—Insights from pH and temperature measurements. J. Environ. Monit. 2012, 14, 1479–1482. [Google Scholar] [CrossRef] [PubMed]
- Anlauf, H.; D’Croz, L.; O’Dea, A.A. corrosive concoction: the combined effects of ocean warming and acidification on the early growth of a stony coral are multiplicative. J. Exp. Mar. Biol. Ecol 2011, 397, 13–20. [Google Scholar] [CrossRef]
- Uddin, S.; Al Shamroukh, D.; Al-Khabaz, A.; Al-Yagoub, A. Assessment, and Monitoring of Water Quality for the Sabah Al Ahmed Sea City Project, Phase II; Technical Report KISR10980; Kuwait Institute for Scientific Research: Safat, Kuwait, 2014 2014; 30p. [Google Scholar]
- Andersson, A.J.; Mackenzie, F.T.; Gattuso, J.-P. Effects of ocean acidification on benthic processes, organisms, and ecosystems. In Ocean acidification, Gattuso J.-P.; Hanson, L., Ed.; Oxford University Press: New York, NY, USA, 2011; pp. 122–153. [Google Scholar]
- Eyre, B.; Andersson, A.; Cyronak, T. Benthic coral reef calcium carbonate dissolution in an acidifying ocean. Nat. Clim. Chang. 2014, 4, 969–976. [Google Scholar] [CrossRef]
- Allemand, D.; Tambutte, E.; Zaccola, D.; Tambutte, S. Coral calcification, cells to reefs. In Coral reefs: An ecosystem in transition; Dubinsky, Z., Stambler, N., Eds.; Springer: New York, NY, USA, 2011; pp. 119–150. [Google Scholar]
- Anderson, L.; Dyrssen, D. Alkalinity and total carbonate in the Arabian Sea. Carbonate depletion in the Red Sea and Persian Gulf. Mar. Chem. 1994, 47, 195–202. [Google Scholar] [CrossRef]
- Riegl, B. Corals in a non-reef setting in the southern Persian Gulf (Dubai, UAE): fauna and community structure in response to recurring mass mortality. Coral Reefs 1999, 18, 63–73. [Google Scholar] [CrossRef]
- Purkis, S.J.; Riegl, B. Spatial and temporal dynamics of Arabian Gulf coral assemblages quantified from remote-sensing and in situ monitoring data (Jebel Ali, Dubai, U.A.E.). Mar Ecol Prog Ser 2005, 287, 99–113. [Google Scholar] [CrossRef]
- Tumbutte, E.; Allemand, D.; Bourge, I.; Gattuso, J.-P. An improved 45Ca protocol for investigating physiological mechanisms in coral calcification. Mar. Biol. 1995, 122, 453–459. [Google Scholar] [CrossRef]
- Lewis, E.; Wallace, D.; Allison, L.J. Program Developed for CO2 System Calculations; ORNL/CDIAC-105; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, US Department of Energy: Oak Ridge, TN, USA, 1998.
- Pierrot, D.; Lewis, E.; Wallace, D. MS Excel Program Developed for CO2 System Calculations; ORNL/CDIAC-105a; Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, US Department of Energy: Oak Ridge, TN, USA, 2006.
- Castillo, K.; Ries, J.; Bruno, J.; Westfield, I. The reef-building coral Siderastrea siderea exhibits parabolic responses to ocean acidification and warming. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141856. [Google Scholar] [CrossRef] [PubMed]
- Hoeke, R.; Jokiel, P.; Buddemeier, R.; Brainard, R. Projected changes to growth and mortality of Hawaiian corals over the next 100 years. PLoS ONE 2011, 6, e18038. [Google Scholar] [CrossRef] [PubMed]
- Jokiel, P. Predicting the impact of ocean acidification on coral reefs: evaluating the assumptions involved. ICES J. Mar. Sci. 2016, 73, 550–557. [Google Scholar] [CrossRef]
- Coles, S.L. Coral species diversity and environmental factors in the Arabian Gulf and the Gulf of Oman: a comparison to the Indo-Pacific region. Atoll. Res. Bull. 2003, 507, 1–19. [Google Scholar] [CrossRef]
- Sheppard, C.; Al-Husiani, M.; Al-Jamali, F.; Al-Yamani, F.; Baldwin, R.; Bishop, J.; Benzoni, F.; Dutrieux, E.; Dulvy, N.K.; Durvasula, S.R.V.; et al. Environmental Concerns for the Future of Gulf Coral Reefs. In Coral Reefs of the Gulf: Adaptation to Climatic Extremes; Riegl, B.M., Purkis, S.J., Eds.; Springer Science and Business Media B.V.: New York, NY, USA, 2012. [Google Scholar]
- Bauman, A.G.; Baird, A.H.; Cavalcante, G.H. Coral reproduction in the world’s warmest reefs: southern Persian Gulf (Dubai, United Arab Emirates). Coral Reefs 2011, 30, 405–413. [Google Scholar] [CrossRef]
- Schoepf, V.; Stat, M.; Falter, J.L.; McCulloch, M.T. Limits to the thermal tolerance of corals adapted to a highly fluctuating, naturally extreme temperature environment. Scientific Reports 2015, 5, 1–14. [Google Scholar] [CrossRef]
- Purkis, S.; Renegar, D.; Riegl, B. The most temperature-adapted corals have an Achilles’ Heel. Mar. Poll. Bull. 2011, 62, 246–250. [Google Scholar] [CrossRef]
- Riegl, B.; Purkis, S.J. Coral Reefs of the Gulf Adaptation to Climatic Extremes; Springer: New York NY, USA, 2012. [Google Scholar]
- Burt, J.; Bartholomew, A.; Usseglio, P. Recovery of corals a decade after a bleaching event in Dubai, United Arab Emirates. Mar. Biol. 2008, 154, 27–36. [Google Scholar] [CrossRef]
- Wilkinson, C.; Hodgson, G. Coral reefs and the 1997–1998 mass bleaching and mortality. Nat. Resour. 1999, 35, 16–25. [Google Scholar]
- Wilkinson, C.R.; Linden, O.; Cesar, H.; Hodgson, G.; Rubens, J.; Strong, A.E. Ecological and socio-economic impacts of 1998 coral mortality in the Indian Ocean: an ENSO (El Nino-Southern Oscillation) impact and a warning of future change? Ambio 1999, 28, 188–196. [Google Scholar]
- Edmunds, P.J.; Cumbo, V.; Fan, T.Y. Effect of temperature on the respiration of brooded larvae from tropical reef corals. J. Exp. Biol. 2011, 138, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Baria, M.; Kurihara, H.; Harii, S. Tolerance to elevated temperature and ocean acidification of the larvae of the solitary corals Fungia fungites (Linnaues, 1758) and Lithophyllon repanda (Dana, 1846). Zool Sci. 2015, 32, 447–454. [Google Scholar]


{kind=link}
| Nominal pH | Temperature [°C] | pHnbs | pCO2 [atm] | Ωcalc | Ωar |
|---|---|---|---|---|---|
| 8.2 | 22.5 | 8.18 ± 0.003 | 451 ± 6 | 6.48 ± 0.05 | 4.24 ± 0.04 |
| 27.5 | 437 ± 5 | 7.21 ± 0.04 | 4.80 ± 0.03 | ||
| 32.5 | 447 ± 6 | 7.81 ± 0.06 | 5.29 ± 0.04 | ||
| 7.5 | 22.5 | 7.50 ± 0.002 | 2453 ± 16 | 1.54 ± 0.01 | 1.01 ± 0.01 |
| 27.5 | 2525 ± 28 | 1.75 ± 0.02 | 1.17 ± 0.01 | ||
| 32.5 | 2688 ± 18 | 2.00 ± 0.01 | 1.36 ± 0.01 | ||
| ANOVA 3 | |||||
| Model (F11,83, p) | 4015.9 <0.0001 | 2422.5 <0.0001 | 2751.1 <0.0001 | 2763.4 <0.0001 | |
| pH (F1, p) | 44157.0 <0.0001 | 26529.3 <0.0001 | 29578.1 <0.0001 | 29454.0 <0.0001 | |
| Temperature (F2, p) | 1.39 0.26 | 28.38 <0.0001 | 272.2 <0.0001 | 372.2 <0.0001 | |
| pH x temperature (F2, p) | 0.63 0.54 | 28.95 <0.0001 | 65.52 <0.0001 | 95.0 <0.0001 | |
| Replicate (F6, p) | 0.38 0.89 | 0.47 0.83 | 1.47 0.20 | 1.45 0.21 | |
| F | P | |
|---|---|---|
| Acropora pharaonis | ||
| Model | F11,35 = 74.3 | <0.0001 |
| pH | F1 = 190.0 | <0.0001 |
| Temperature | F2 = 258.4 | <0.0001 |
| pH × Temperature | F2 = 51.0 | <0.0001 |
| Replicate | F6 = 1.41 | 0.25 |
| Porites lutea | ||
| Model | F11,35 = 114.71 | <0.0001 |
| pH | F1 = 298.4 | <0.0001 |
| Temperature | F2 = 415.6 | <0.0001 |
| pH × Temperature | F2 = 61.83 | <0.0001 |
| Replicate | F6 = 1.44 | 0.24 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behbehani, M.; Uddin, S.; Dupont, S.; Sajid, S.; Al-Musalam, L.; Al-Ghadban, A. Response of Corals Acropora pharaonis and Porites lutea to Changes in pH and Temperature in the Gulf. Sustainability 2019, 11, 3156. https://doi.org/10.3390/su11113156
Behbehani M, Uddin S, Dupont S, Sajid S, Al-Musalam L, Al-Ghadban A. Response of Corals Acropora pharaonis and Porites lutea to Changes in pH and Temperature in the Gulf. Sustainability. 2019; 11(11):3156. https://doi.org/10.3390/su11113156
Chicago/Turabian StyleBehbehani, Montaha, Saif Uddin, Sam Dupont, Sufiya Sajid, Lamya Al-Musalam, and Abdulnabi Al-Ghadban. 2019. "Response of Corals Acropora pharaonis and Porites lutea to Changes in pH and Temperature in the Gulf" Sustainability 11, no. 11: 3156. https://doi.org/10.3390/su11113156
APA StyleBehbehani, M., Uddin, S., Dupont, S., Sajid, S., Al-Musalam, L., & Al-Ghadban, A. (2019). Response of Corals Acropora pharaonis and Porites lutea to Changes in pH and Temperature in the Gulf. Sustainability, 11(11), 3156. https://doi.org/10.3390/su11113156