Abstract
Despite the great importance of fine roots, which are referred to as roots smaller than 2 mm in diameter, in terms of carbon and nutrient cycling in terrestrial ecosystems, how fine root biomass, production, and turnover rate change with stand development remains poorly understood. Here we assessed the variations of fine root biomass, production, and morphology of trees and understory vegetation in Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) plantations at the ages of 7 years old, 17 years old and 25 years old in southern China, representing the sapling, pole and mature stage, respectively. Fine roots of trees and understory vegetation were sampled with sequential coring method to a depth of 60 cm and sliced into 4 layers (0–15, 15–30, 30–45 and 45–60 cm). Fine root biomass and necromass were highest in the pole stages among these three different aged Chinese fir plantations, although the significant differences were only detected for fine root necromass between 25-year-old and 7-year-old plantations. Fine root biomass of Chinese fir was heterogeneous in both temporal and spatial dimensions. Seasonal variation of fine root biomass in three age groups showed a similar pattern that the standing fine root biomass reached a peak in January and fell to the lowest in July. Vertically, the fine root biomass decreased with the increase of soil depth, but this extinction rate decreased with stand development. The effects of stand age on either total fine root length and surface area, or specific root length were not significant. However, the root tissue density increased significantly with Chinese fir stand ages, which suggested that the fine roots on Chinese fir may resort more to the mycorrhizal associations for the nutrient and water acquisition in the later stage of Chinese fir plantations. In addition to the stand age effect, the fine roots exhibited highly spatial and temporal variations in Chinese plantations, indicating different root foraging strategies for soil nutrient and water acquisition. Therefore, the fine root research not only helps to understand its role in carbon sequestration in terrestrial ecosystem under global climate change, but can also improve our understanding of nutrient management in forest ecosystem. At the same time, the research on the productivity of the Chinese fir growth stage provides guiding significance for the construction and management of Chinese fir.
1. Introduction
Fine roots, which are usually defined as roots smaller than 2 mm in diameter, are the most physiologically active organs in forest soils. They are responsible for nutrient and water acquisition from soil and it is widely recognized that fine roots play a significant role in the carbon and biogeochemical cycle in forest ecosystems [1]. It has been estimated that approximately one third of annual net primary productivity was consumed in fine roots due to the rapid growth and turnover rate [2]. Consequently, fine roots are the major contributors for nutrient flux between the interface of plant and soil in forest ecosystems. Thus, the accurate estimates of fine root biomass and production are not only essential for assessing carbon budgets in forests and improving model accuracy in climate change mitigation, but also for sustainable management of forests in terms of nutrient cycling.
The activity of fine roots in forests has been regarded to be under strong genetic control and different species would adapt different strategies to meet the nutrient and water demands from soils [3]; fine roots also show variations in biomass, spatial distribution and morphology in response to stand ages and environmental changes [4]. Previous studies have demonstrated that fine root production increased with stand age. In contrast, Finer et al. [5] revealed that fine root production decreased with stand development in the southern boreal forest, whereas Yuan and Chen [6] reported that fine root production increased to a peak and then decreased at later stages in boreal forest. The dynamics of fine root biomass in different stand ages are related to above-ground stand structures, which are in association with stand development [7]. Fine root growth rate is crucial for estimating fine root dynamics and carbon cycle [8,9]. In boreal forests, fine root biomass generally increases as plants start to grow in spring and increase continually to a peak in mid-summer and then decreases in the fall [10]. So far, many studies on fine root biomass and production along chronosequence have been conducted in boreal and temperate forests, while the effects of stand age on fine root biomass and production have been relatively seldom studied in subtropical forests [11].
Generally, fine roots are capable of adapting themselves to changing environments quickly. Therefore, fine root could be used as a sensitive indicator of changes in tree physiological traits and environmental conditions [3,12]. Previous results have demonstrated that fine root biomass is dependent upon a wide range of both biotic and abiotic factors, such as climatic conditions (geographical location, precipitation, elevation and annual temperature), soil properties (texture, moisture, chemistry, nutrient availability), and stand characteristics (basal area, stand density, stand age, tree species, and intra-and interspecific competition between plants) [13,14]. In addition, fine roots are prone to modify root system architecture characteristics without changing the biomass [15]. Thus, fine root morphological plasticity has been suggested as a potential mechanism by which plants respond to changes of soil resource supply. A series of root’s morphological parameters vary with soil physical conditions and different ages [16]. For instance, specific root length (SRL) is often considered as the efficiency of roots to proliferate in the soil per unit of carbon invested, and is thus related to potential ability of nutrient uptake [17]. A higher SRL normally implies rapid nutrient uptake efficiency per unit root biomass [18]. Root tissue density is also considered to reflect absorptive potential and turnover [19]. However, there are few data on how root morphology changes with association to forest stand age.
Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) is a fast growth species and one of the popular subtropical tree species for timber production in subtropical China. It covers 1.096 × 107 ha, accounting for 6.66% of total forest coverage and 4.91% of storage volume in China [20]. Traditionally, Chinese fir plantation has been clear-cut when it reaches to the mature stage, which is around 20 years old. The log and debris removal, and prescribed burning before planting for next rotation can lead to nutrient losses. Consequently, concerns have been raised regarding the soil degradation and sustainable management of the following Chinese fir rotations. Here, the effects of stand age on fine root biomass, distribution, and morphology were investigated in the Chinese fir plantations at ages of 7 years, 17 years and 25 years. We aimed to assess the different foraging strategies of fine roots in Chinese fir plantations at different development stages and provide a potential guide for the nutrient management for Chinese fir plantations. More specifically, we aimed to evaluated (a) fine root biomass, distribution and seasonal variations in Chinese fir plantations at different stand ages; (b) fine root production and turnover rate along chronosequence; and (c) Responses of fine root morphology to stand age were coupled with fine root biomass and production.
2. Materials and Methods
2.1. Study Site and Experimental Design
The study site was located in the National Field Station for Scientific Observation and Research of Chinese fir Plantation Ecosystem in Huitong, Hunan Province, China, where the Chinese fir plantation cultivation center is located. In the 1970s, seven parallel watersheds were setup as long-term field observation sites for Chinese fir plantations (109°45′ E, 26°50′ N) (Figure 1). We conducted this research in watershed II and III with areas of ca. 2.0 ha, which were planted in 1996 and 1988 after a clear-cut of the first Chinese fir rotation to represent the pole and mature stage, respectively. As there was no young Chinese fir plantation available in the national field station, we selected the Chinese fir plantations at the sapling stage approximately 2 km apart from our main site, which was planted in 2006. Elevation is 270–400 m above sea level. The study area belongs to a typical monsoon climate with mean annual temperature of 16.8 °C and mean temperature of 4.4 °C in Jan. and 26.3 °C in July. Average annual precipitation is around 1400 mm and the average annual relative humidity is around 87%. The soils are fertile with an average depth of 1.0 m. Due to the prescribed residue burning before planting and 3-year weeding after afforestation, along with the fast growth of Chinese fir, there was almost no other tree species therein and the understory layer was mainly shrub and herbs, including Maesa japonica, Miscanthus floridulus, Ampelopsis cantoniensis, Houttuyni acordata and Parthenocissus quinquefolia.
Figure 1.
Location of Huitong National Field Research Station of Chinese fir plantation.
Three plots with size of 20 × 20 m were established in the 7-year, 17-year and 25-year-old Chinese fir plantations in 2013. All the trees were labeled and measured for diameter at breast height (DBH), height and crown area. The stand characteristics were shown in Table 1.

Table 1.
Stand structure of Chinese fir plantations at different ages.
2.2. Fine Root Sampling
Sampling was conducted by using sequential coring method with a sharp steer auger (5 cm in diameter, 30 cm height) to the depth of 60 cm and sliced into four layers (0–15, 15–30, 30–45 and 45–60 cm) in January, April, July, October of 2013, representing winter, spring, summer and autumn, respectively. Four soil cores were collected in three transects on the upper, middle and lower slope in each plot, respectively. Considering the fine root horizontal variation, the sampling locations were at least 3 m apart and two soil samples from the same slope position at the same specific soil layers were composited as one sample. Therefore, there were 24 samples from each plot and 72 samples from each stand at different ages, and in total 216 samples in each sampling occasions. Soil samples were transported into a laboratory and stored in refrigeration (4 °C) until they were processed within 1 month.
In order to separate fine roots from the soil, the samples were shortly soaked in the water and washed carefully to remove the soil. The roots suspended in the water were poured into a sieve with 0.56 mm mesh size. Then fine roots were sorted into roots of Chinese fir or understory vegetation groups visually according to the color, shape and root forks; and the Chinese fir fine roots were distinguished further into living roots and dead roots according to the color, elasticity and degree of brokenness. The Chinese fir fine root was scanned and the digital images were analyzed with software WinRhizo 2013 (Regent, Canada) to determine its morphological traits, including length, surface area (cm2), root volume (cm3) and average diameter [21]. Then, the fine root samples were oven dried to constant weight at 60 °C, and weighted to 0.0001 g with an electronic scale. Fine root biomass and necromass were calculated as live root dry weight and dead root weight per unit area square meter (m2), respectively. Understory root biomass was root biomass of shrub and herbs. The fine root parameters were calculated with formula:
Fine root biomass (g m−2) = root dry weight (g)/(π × 2.52 × 2 × 10−4)
Fine root length (km m−2) = total length (m) × 10−3/(π × 2.52 × 2 × 10−4)
Fine root surface area (m2 m−2) = total surface area × 10−3/(π × 2.52 × 2)
Specific root length (SRL) (m g−1) = root length (cm) × 10−2/dry weight (g);
Tissue density (g cm−3) = dry weight (g)/root volume (cm3)
2.3. Statistical Analyses
Data of the fine root from three stand ages, four soil layers and four sampling occasions were analyzed. All data sets on fine root mass, morphology and architectural traits were tested for normal distribution using a Shapiro-Wilk test. The effects of stand age, soil depth and sampling season on the fine root biomass, necromass, understory vegetation root mass and morphological variables were tested with a linear mixed-effect model using the slope position as random effects. As we only investigated three different aged plantations here, we treated stand age as a categorical variable. The linear mixed-effect analysis was performed using restricted maximum likelihood estimation with the lme4 package. To ensure the normality and homoscedasticity, a Box-cox transformation was applied to all dependent variables with λ = 0.15 [22,23]. Comparison of fine root biomass and morphological variables in the Chinese fir plantation in different ages were analyzed by one-way ANOVA, the method of multiple comparisons used the least significant difference (LSD). In cases when data did not meet the requirements for parametric tests, Mann-Whitney U test (Wilcoxon’s rank test) was conducted.
The comparisons of three methods (sequential soil coring, ingrowth methods, and minirhizotrons) for estimating forest fine root production and turnover revealed that the measured results were similar [1,24]. With sequential coring methods, there were also three methods to calculate production and turnover (decision matrix, the max-min, compartment) and previous results showed that the decision matrix method could represent best the actual value [25]. Therefore, here we applied the decision matrix method to estimate the fine root production [23] and the fine root turnover rate was calculated with formula [24]:
Fine root turnover rate (year−1) = production/mean biomass
All the statistical analysis and graphics were performed with the software Excel 2016 and R 3.3.3 (R Development Core Team, Vienna, Austria, 2017).
3. Results
3.1. Root Biomass and Necromass
The standing fine root biomass and necromass of Chinese fir showed a similar pattern that the 17-year-old plantations showed the highest value in these three different aged Chinese fir plantations. The significant differences were detected with linear mixed-effect model, and fine root necromass in the 17-year-old plantations was significantly higher than that in the 25-year-old plantations (Table 2 and Figure 2). The fine root biomass was 243.02, 261.96 and 233.39 g m−2 on average to soil depth of 60 cm in the 7-year-old, 17-year-old and 25-year-old plantations, respectively. In contrast, the understory root biomass, which consisted of all the herb and shrub roots, was significantly higher in the 7-year-old plantation than that in the 17-year-old and 25-year-old plantations. It reached 204.25 ± 17.65 g m−2. The live fine root length and surface area of Chinese fir was slightly higher in 7-year-old plantations than that in 17-year-old and 25-year-old plantations. The fine root length was 1.99 ± 0.12, 2.06 ± 0.08, 1.81 ± 0.13 km m−2, and the live root surface area was 4.57 ± 0.27, 4.67 ± 0.186 and 4.06 ± 0.29 m2 m−2 in the 7-year-old, 17-year-old and 25-year-old plantations, respectively (Figure 3).
Table 2.
Effects of stand age, soil depth and season on fine root biomass, necromass, understory root biomass, fine root length, fine root surface area, specific root length and root density. Bold font indicates significant differences at P < 0.05.
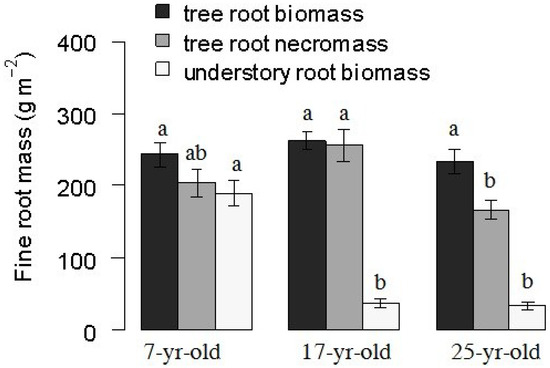
Figure 2.
Tree fine root biomass, necromass and understory root biomass to the soil depth of 60 cm in 7-year, 17-year, 25-year-old Chinese fir plantations. Data are means of four sampling occasions from April 2014 to January 2015. Error bars indicate standard error. Different letters indicate significant differences among different stand ages (P < 0.05).
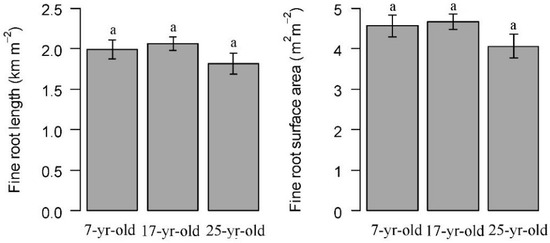
Figure 3.
Total live fine root length and surface area in Chinese fir plantations at different ages. Error bars indicate standard error. Different letters indicate significant differences among different stand ages (P < 0.05).
The slope position substantially affected the fine root biomass of Chinese fir in these different aged Chinese fir plantations in this study. In 7-year-old stands, the fine root biomass decreased with increasing elevation, although no significant differences were detected. In contrast, the fine root biomass increased significantly along the increasing slope consistently in the 17-year-old and 25-year-old plantations (Figure 4).
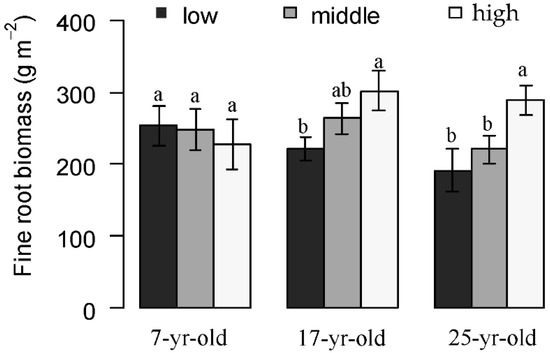
Figure 4.
Fine root biomass at different slope positions in Chinese fir plantations at different stand ages. Error bars indicate standard error. Different letters indicate significant differences among different slope locations at the same aged plantations (P < 0.05).
In contrast to the stand age, soil depth and season exhibited a significant effect on all the fine root variables (P < 0.05) (Table 2). In these three different aged Chinese fir plantations the fine root biomass decreased with increasing soil depth. Fine root biomass in the top soil layer (0–15 cm) was significantly higher than that in the deeper layer (15–60 cm). However, this vertical decreasing gratitude was different. In 7-year-old plantations, fine root biomass declined dramatically from the top layer (0–15 cm) to the 30–45 cm layer and remained steady to the 45–60 cm layer, while in 25-year-old stands the fine root distributed gradually from the top layer (0–15 cm) to the deepest layer (45–60 cm). Besides, the fine root biomass was almost the same in the deepest layer (45–60 cm) in Chinese fir plantations at different ages (Figure 5). The standing fine root biomass varied seasonally and showed a similar pattern temporally as the fine root biomass was the highest in the winter (January) and the lowest in the summer (July) for all the Chinese fir plantations at different ages (Figure 6).
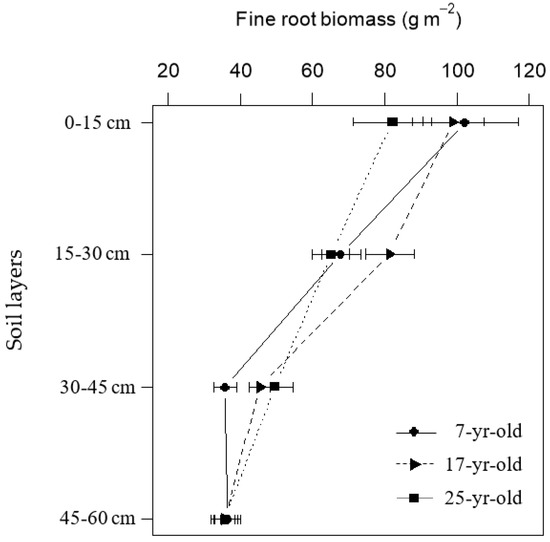
Figure 5.
Vertical distribution of fine root biomass with increasing soil depth in Chinese fir plantations at different stand ages.
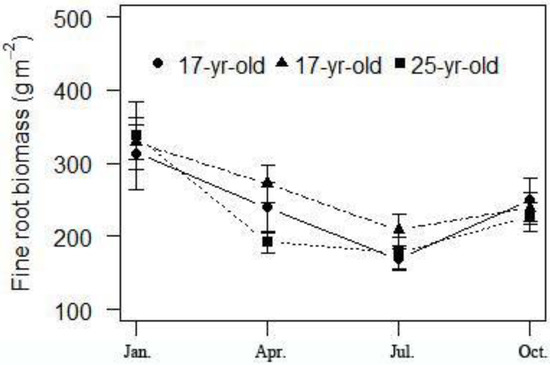
Figure 6.
Seasonal variations of fine root biomass in Chinese fir plantations at different ages.
3.2. Fine Root Production and Turnover
Fine root production and turnover varied dramatically with stand age. Fine root production and turnover were consistent with the trend of age, and the value increased first and then decreased with Chinese fir stand development. The fine root production was 352.54, 410.85 and 217.61 g m−2 year−1, and turnover rate was 1.48, 1.57 and 0.92 year−1 in 7-year-old, 17-year-old and 25-year-old plantations, respectively (Figure 7).
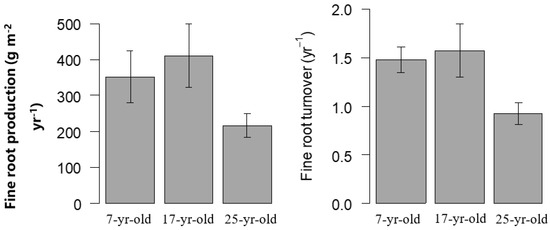
Figure 7.
Fine root production and turnover in Chinese fir plantations at different stand ages.
3.3. Fine Root Morphology
The fine root specific root length of Chinese fir was 10.02, 9.65 and 10.08 m g−1 on average in 7-year-old, 17-year-old and 25-year-old plantations, respectively; no significant differences were detected among them. However, the root tissue density increased consistently with stand development and the root tissue density was significantly higher in 17-year-old plantations (0.30 g cm−3) and 25-year-old plantation (0.31 g cm−3) than that in 7-year-old plantations (0.27 g cm−3) (Figure 8).
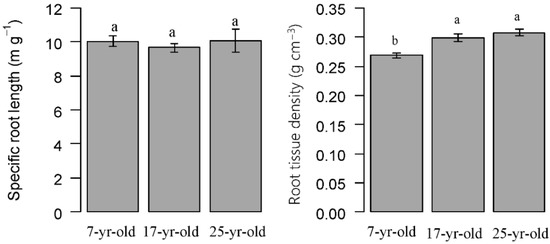
Figure 8.
Comparison of specific root length and root tissue density in Chinese fir plantations at different ages. Error bars indicate standard error. Different letters indicate significant differences among different stand ages (P < 0.05).
4. Discussion
4.1. Fine Root Biomass and Necromass
In this study, we focused on the variation of tree fine root biomass on sapling, and pole and mature stages of Chinese fir plantations. Taking into account the three stands studied, the mean fine root biomass was similar to the value of 230 g m−2 reported in boreal forests, but relatively lower than the value of 500 g m−2 reported in the temperate coniferous forest [26]. The value was in the range of the fine root biomass (1.1~5.8 t hm−2) of the subtropical forest [7] and similar to the value reported in Chinese fir plantations by Yang et al. [27], but higher than the value reported by Li et al. [28]. Compared with broad-leaved forests or natural forests, fine root biomass of the Chinese fir plantation was relatively lower [27,29]. Our result also supported the findings that fine root biomass increased first and then decreased with increasing stand age [30]. The variations in fine root biomass at different stand ages in forests may be related to many factors, including canopy closure, stand tree density, aboveground standing biomass, local site conditions, soil depth and previous management practices [31]. In terms of an individual tree, an old tree may produce more fine root biomass than the younger ones. On the stand level, however, this was not always the case. Helmisaari et al. [32] demonstrated that the fine root biomass reached the peak when the stand density was maximum. Furthermore, the soil conditions might also affect the amount of carbon allocated to belowground fine roots. Cao [33] investigated the soil organic carbon content of Chinese fir at different stand ages in the same region, which first decreased and then increased from the 7-, 17- to 25-year-old plantation, and may imply that the soil organic carbon is closely related to the fine root biomass. In the previous study, it also revealed that the underground growing space was suppressed with the closure of the canopy layer [14]. The study also confirmed this statement as the understory vegetation root biomass declined dramatically from 7-year-old plantations to 17-year-old and 25-year-old plantations (Figure 2).
Slope positions, indicating the micro-variation of soil conditions here, lead to differences in soil nutrients and moisture, resulting in differences in fine root biomass. In general, the uphill biomass was smaller than the downhill because the downhill site was normally better than the uphill as the downhill was better able to meet the needs of the tree’s moisture and nutrients. Li et al. [28] found that the biomass of all fine root types was the highest in the low slope position, and the uphill pit was the lowest, which was different from this study. Here in the 7-year-old plantations, the fine root biomass was ranked as downhill > mid-slope > uphill, and 17-year-old and 25-year-old plantations showed the opposite. This is in agreement with the result of Fu et al. [34] that C. lanceolata seedlings in different stand types, that were studied together for 2-year-old, were dominant in Huitong. It is likely that the root grew first in the resource hotspot in the young stands, and the plant could maximize the nutrient return to supply the plant growth; when experiencing stress in space, nutrients and moisture, the plants were forced to invest more carbon to roots for nutrient and water absorption in 17-year-old and 25-year-old plantations [35].
The spatial structure and distribution of roots in the soil were determined by the amount of resources acquired by plants and the soil competitiveness of individuals or groups of plants [24]. Tree fine root distribution could be affected by soil nutrient content, soil texture and physicochemical properties as well. The vertical stratification was in agreement with many other studies in forest ecosystems [36]. In all stands, the study showed fine root biomass decreased with soil depth, and there was an obvious vertical gradient in fine root mass distribution, with the highest biomass in the upper soil layers [37]. With the increasing year, the litter accumulated on the surface, and fine root tended to surface to absorb more nutrients [38]. The study was different from this conclusion. In this study, the fine roots biomass in the deepest layer (45–60 cm) were similar in these three different aged Chinese fir plantation, suggesting the belowground root exploitation and penetration were sufficient even in the young sapling stages. However, the fine roots were concentrated on the top soil layer in this sapling stage (7-year-old plantations), probably due to the enriched nutritional content derived by the prescribed burning before the establishment, or the Chinese fir adapted to the competition from the understory vegetation, which was reflected by the high understory root biomass here. Due to high annual rainfall, and fragile soils and steep slopes, the subtropical forests in China were susceptible to nutrient loss, especially in artificial forests. This may reflect the nutrient distribution pattern that returned to the soil from aboveground litter and belowground fine root input [25]. The fine root biomass in the subsoil layer (15–30 and 30–45 cm) increased with stand age.
Fine root biomass would fluctuate within a certain year, which may be due to changes in soil temperature and moisture [39]. The studies on seasonal variations in root biomass had reported that stand ages were consistent with the seasonal variation pattern [40]. Fu et al. [34] believed that low temperature is the influencing factor of fine root dynamics. Some scholars believed that the rising soil temperature and rainfall will lead to a fine root vigorous growth period in the early spring season with sufficient carbohydrates stored in the previous season to store [41]. Yang and Li [42] had given an overview describing fine root distribution research and pointed out that the peak of the community fine root growth occurred in July and October in the broad-leaved and conifer mixed forest. Makkonen and Helmisaari [41] inferred that fine root biomass arose in about March or April. It is surprising that our results showed different patterns, that fine root biomass was highest in January and lowest in July in all the Chinese fir plantations. Although similar results also had been reported that the minimum value of fine root biomass generally appeared from May to July [40,43]. The fine root biomass dynamics were not only related to the fine root growth, but also the longevity and turnover rate, which merits further investigation.
4.2. Fine Root Production and Turnover
There were many factors that determine the fine root production and turnover rate, such as calculation methods, sampling methods, changing in biomass, stand age and so on. In the present study, fine root production and turnover increased within the age from 7-year-old, 17-year-old stands, and then decreased in the 25-year-old stands. These results were consistent with some of the reported surveys. For instance, Yuan and Chen [44] revealed that in boreal forest fine root production increased initially to a peak, and declined thereafter, although other studies showed positive [32], negative stand age effects [45] or no effects on fine root production and turnover rate [46].
4.3. Fine Root Morphology
The fine root architecture was composed of several parameter for soil resource acquisition efficiency, which reflected the fine root plasticity in response to environmental change [47]. The study indicated that there is a certain relationship to the fine root biomass and morphology. Trees either invest more carbon underground to increase fine root biomass and production, or improve their nutrient and water uptake efficiency from the soil by changing root morphology. A number of studies have demonstrated that fine root biomass and morphology are closely related to tree species and stand ages [14,35]. Thus, estimating changes in underground biomass, production, and morphology is crucial to understand the fine root dynamics and processes in the development of forest ecosystems. The total fine root length and surface area did not show significant differences among Chinese fir plantations at different stand ages (Table 2 and Figure 3), neither did the SRL (Figure 8). SRL was used as an indicator of environmental change and was linked to the ability of nutrient, water uptake efficiency [15,18]. In the Chinese fir plantation, the SRL remained relatively stable. In contrast, root tissue density, which was considered as an indicator for root functional status [48], showed high variations. The root tissue density increased consistently with stand age. The result was in agreement with the previous study by Borja et al. [2]. It is likely that that the lignification of fine root may also increase as the age of the tree increases, and the nutrient uptake level of the fine root may resort more to the mycorrhizal associations, as suggested by Bauhus et al. [49]. In addition, other several studies demonstrated that the traits of relatively high root tissue density and low SRL indicated that fine roots adapt to the poor resource environment by reducing absorptive capacities and prolong the fine root longevity [18].
5. Conclusions
This study investigated the spatial and temporal variations of fine root biomass and morphology in different ages, as well as fine root annual productivity and turnover rate. The results showed that the age effect on fine root biomass production and specific root length was not significant, although the fine root biomass, production and turnover rate were higher in the pole stage than that in sapling and mature stages. However, the fine roots of Chinese fir showed high variations spatially and temporally. Fine root biomass decreased with stand ages in all Chinese fir plantations, but it was more evenly distributed with increasing stand ages. The fine root biomass peaked in January and reached the lowest value in July, which merits further investigation. Furthermore, the root tissue density increased significantly with stand ages, suggesting the Chinese fir plantations might resort to the mycorrhizal association for this conifer species. The fine roots were the main organs responsible for water and nutrient uptake for tree species; the research on it not only helps to understand the relationship between plant structure and function, but also helps to explore the carbon pattern and process of the terrestrial ecosystem under global change. Correct understanding of the production and turnover of fine roots is of great significance to the scientific guidance of production practice in order to improve the productivity of forest ecosystems. In order to obtain a more comprehensive understanding of Chinese fir fine roots, it is necessary to explore the aspect of mycorrhizal associations and root longevity as well.
Author Contributions
P.L. and W.X. initiate the conceptual idea and designed the experiments; Y.P., Y.X. and S.O. conducted the experiments; Y.P. and P.L. analyzed the data; W.X. and S.O. contributed technical advice. Y.P. and P.L. develop the draft and revision.
Funding
This study was sponsored by the National Key Research and Development Program of China (2016YFD0600202), Young Talent Projects from Education Department of Hunan Province (14B195).
Acknowledgments
We are grateful to the ecological station of Huitong to provide experimental base. We would like to thank Dongjiao Kuang and Xiaoxiao Shen for the assistance in the field and laboratory work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lukac, M. Fine Root Turnover. Meas. Roots 2012, 363–373. [Google Scholar] [CrossRef]
- Borja, D.W. Steffenrem and Majdi. Effect of stand age on fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway. Tree Physiol. 2008, 28, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, M.; Inagaki, Y.; Kamo, K.; Titin, J. Fine-root production in response to nutrient application at three forest plantations in Sabah, Malaysia: Higher nitrogen and phosphorus demand by Acacia mangium. J. For. Res. 2009, 14, 178–182. [Google Scholar] [CrossRef]
- Jia, Q.; Liu, Q.; Li, J. Individual-based fine root biomass and its functional relationship with leaf for Pinus tabuliformis in northern China. Eur. J. For. Res. 2015, 134, 705–714. [Google Scholar] [CrossRef]
- Finer, S.E.; Finer, S.E. The History of Government from the Earliest Times: Ancient Monarchies and Empires; Oxford University Press: Oxford, UK, 1997; Volume 1. [Google Scholar]
- Yuan, Z.Y.; Chen, H. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Vogt, K.A.; Grier, C.C.; Vogt, D.J. Production, turnover, and nutrient dynamics of above- and belowground detritus of world forests. Adv. Ecol. Res. 1986, 15, 303–377. [Google Scholar]
- Fukuzawa, K.; Dannoura, M.; Kanemitsu, S.; Kosugi, Y. Seasonal patterns of root production of Japanese oak seedlings and dwarf bamboo grown in rhizoboxes. Plant Biosyst. 2010, 144, 434–439. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2010, 147, 13–31. [Google Scholar] [CrossRef]
- Paudel, T.; Gao, Y.; Lan, Y.; Macmohan, G.; Kempa, K.; Naughton, M.; Ren, Z. Influence of environmental variability on root dynamics in northern forests. Crit. Rev. Plant Sci. 2009, 28, 179–197. [Google Scholar]
- Kizha, A.R.; Han, H.S. Predicting aboveground biomass in second growth coast redwood: Comparing localized with generic allometric models. Forests 2016, 7, 96. [Google Scholar] [CrossRef]
- Helmisaari, H.S.; Derome, J.; Nöjd, P.; Kukkola, M. Fine root biomass in relation to site and stand characteristics in Norway spruce and Scots pine stands. Tree Physiol. 2007, 27, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Farfał, D. Root vitality in the upper soil of pine stands ten years after thinning. For. Res. Pap. 2010, 71, 225–230. [Google Scholar] [CrossRef]
- Jagodzin´ski, A.M.; Kałucka, I. Fine root biomass and morphology in an age-sequence of post-agricultural Pinus sylvestris L. stands. Dendrobiology. 2011, 66, 71–84. [Google Scholar]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Terzaghi, M.; Montagnoli, A.; Iorio, A.D.; Scippa, G.S.; Chiatante, D. Fine-root carbon and nitrogen concentration of European beech (Fagus sylvatica L.) in Italy Prealps: Possible implications of coppice conversion to high forest. Front. Plant Sci. 2013, 4, 192. [Google Scholar] [CrossRef] [PubMed]
- Persson, H.; Ahlström, K.; Persson, H.; Ahlström, K. Fine-root response to nitrogen supply in nitrogen manipulated Norway spruce catchment areas. For. Ecol. Manag. 2002, 168, 29–41. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Duncan, L.W. Root growth and carbohydrate responses in bearing citrus trees following partial canopy removal. Tree Physiol. 1992, 10, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Craine, J.M. Competition for nutrients and optimal root allocation. Plant Soil 2006, 285, 171–185. [Google Scholar] [CrossRef]
- State Forestry Administration. Eighth Forest Resources Inventory in China; China Forestry Publishing House: Beijing, China, 2014; pp. 1–2.
- Leppälammi-Kujansuu, J.; Salemaa, M.; Dan, B.K.; Linder, S.; Helmisaari, H.S. Fine root turnover and litter production of Norway spruce in a long-term temperature and nutrient manipulation experiment. Plant Soil 2014, 374, 73–88. [Google Scholar] [CrossRef]
- Danise, T.; Fioretto, A.; Innangi, M. Spectrophotometric methods for lignin and cellulose in forest soils as predictors for humic substances. Eur. J. Soil Sci. 2018, 1–12. [Google Scholar] [CrossRef]
- Feng, C.; Wang, Z.; Zhu, Q.; Fu, S.L.; Chen, H.Y.H. Rapid increases in fine root biomass and production following cessation of anthropogenic disturbances in degraded forests. Land Degrad. Dev. 2018, 29, 461–470. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y. Indirect methods produce higher estimates of fine root production and turnover rates than direct methods. PLoS ONE 2012, 7, e48989. [Google Scholar] [CrossRef] [PubMed]
- Vogt, K.A.; Publicover, D.A.; Bloomfield, J.; Perez, J.M.; Vogt, D.J.; Silver, W.L. Belowground responses as indicators of environmental change. Environ. Exp. Bot. 1993, 33, 189–205. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Chen, G.S.; He, Z.M.; Chen, Y.X.; Guo, J.F. Production, distribution and nutrient return of fine roots in a mixed and a pure forest in subtropical China. Chin. J. Appl. Environ. Biol. 2002, 8, 223–233. [Google Scholar]
- Li, S.Z.; Tian, D.L.; Wang, G.J.; Yan, W.D.; Duan, W. Fine root biomass and space-time dynamics of 4 types main plantation community, in Hunan Provine. J. Cent. South Univ. For. Technol. 2011, 31, 51–63. [Google Scholar]
- Liao, L.; Chen, C.; Sinica, A.; Shen, Y. Turnover of fine roots in pure and mixed Cunninghamia lanceolata and Michelia macclurei forests. Chin. J. Appl. Ecol. 1995, 6, 5–10. [Google Scholar]
- Borja, I.; Nilsen, P. Long term effect of liming and fertilization on ectomycorrhizal colonization and tree growth in old Scots pine (Pinus sylvestris L.) stands. Plant Soil 2009, 314, 109–119. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. For. Ecol. Manag. 2011, 262, 2008–2023. [Google Scholar] [CrossRef]
- Helmisaari, H.S.; Makkonen, K.; Kellomaki, S.; Valtonen, E.; Malkonen, E. Below- and above-ground biomass, production and nitrogen use in Scots pine stands in eastern Finland. For. Ecol. Manag. 2002, 165, 317–326. [Google Scholar] [CrossRef]
- Cao, J.; Yan, W.D.; Xiang, W.H.; Chen, X.Y.; Lei, P.F. Chemical characteristics of soil carbon, nitrogen, and phosphorus in the plantation of three cunninghamia lanceolata plantations in hunan province. Sci. Silvae Sin. 2015, 51, 1–8. [Google Scholar]
- Fu, X.; Wang, J.; Di, Y.; Wang, H. Differences in fine-root biomass of trees and understory vegetation among stand types in subtropical forests. PLoS ONE 2015, 10, e0128894. [Google Scholar] [CrossRef] [PubMed]
- Hess, L.; Kroon, H.D. Effects of rooting volume and nutrient availability as an alternative explanation for root self/non-self discrimination. J. Ecol. 2007, 95, 241–251. [Google Scholar] [CrossRef]
- Bennett, J.N.; Andrew, B.; Prescott, C.E. Vertical fine root distributions of western redcedar, western hemlock, and salal in old-growth cedar–hemlock forests on northern Vancouver Island. Can. J. For. Res. 2002, 32, 1208–1216. [Google Scholar] [CrossRef]
- Bakker, M.R.; Augusto, L.; Achat, D.L. Fine root distribution of trees and understory in mature stands of maritime pine (Pinus pinaster) on dry and humid sites. Plant Soil 2006, 286, 37–51. [Google Scholar] [CrossRef]
- Hao, Y.R.; Peng, S.L. Variation of roots and its impact factors in succession. Ecol. Environ. Sci. 2005, 14, 762–767. [Google Scholar]
- Manes, F.; Ricotta, C.; Salvatori, E.; Bajocco, S.; Blasi, C. A multiscale analysis of canopy structure in Fagus sylvatica L. and Quercus cerris L. old-growth forests in the Cilento and Vallo di Diano National Park. Plant Biosyst. 2010, 144, 202–210. [Google Scholar] [CrossRef]
- Yang, Y.S.; Chen, G.S.; Lin, P.; Xie, J.S.; Guo, J.F. Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in Subtropical China. Ann. For. Sci. 2004, 61, 617–627. [Google Scholar] [CrossRef]
- Makkonen, K.; Helmisaari, H.S. Fine root biomass and production in Scots pine stands in relation to stand age. Tree Physiol. 2001, 21, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.Y.; Li, W.H. The underground root biomass and C storage in different forest ecosystems of Changbai Mountains in China. J. Nat. Resour. 2003, 18, 204–209. [Google Scholar]
- Chen, Y. The seasonal dynamics and distribution of fine root biomass in artificial ormosia xylocarpa forest. J. Subtrop. Resour. Environ. 2006, 1, 63–66. [Google Scholar]
- Yuan, Z.Y.; Chen, H.Y.H.; Ostle, N. Fine root dynamics with stand development in the boreal forest. Funct. Ecol. 2012, 26, 991–998. [Google Scholar] [CrossRef]
- Messier, C.; Puttonen, P. Coniferous and non-coniferous fine-root and rhizome production in Scots pine stands using the ingrowth bag method. Silva Fenn. 1993, 27, 245–256. [Google Scholar] [CrossRef]
- Finér, L.; Messier, C.; Grandpré, L.D. Fine-root dynamics in mixed boreal conifer—Broad-leafed forest stands at different successional stages after fire. Can. J. For. Res. 1997, 27, 304–314. [Google Scholar] [CrossRef]
- Lynch, J. Root architecture and plant productivity. Plant Physiol. 1995, 109, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ugawa, S.; Miura, S.; Iwamoto, K.; Kaneko, S.; Fukuda, K. Vertical patterns of fine root biomass, morphology and nitrogen concentration in a subalpine fir-wave forest. Plant Soil. 2010, 335, 469–478. [Google Scholar] [CrossRef]
- Bauhus, J.; Messier, C. Soil exploitation strategies of fine roots in different tree species of the southern boreal forest of eastern Canada. Can. J. For. Res. 1999, 29, 260–273. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).