Garden Pollinators and the Potential for Ecosystem Service Flow to Urban and Peri-Urban Agriculture

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
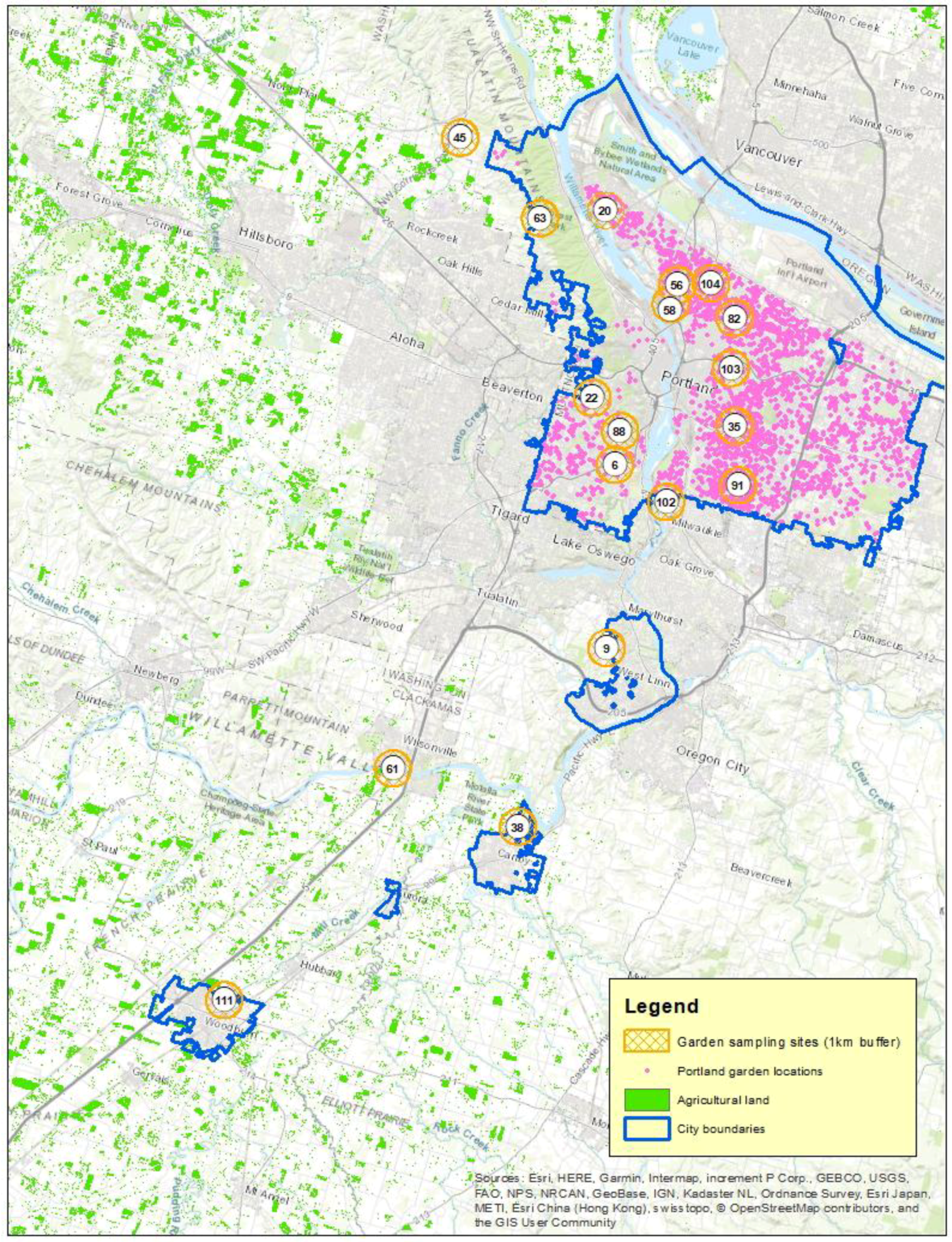
2.1. Study Sites
2.2. Bee Size and Foraging Distance
2.3. Agricultural Context
2.4. Residential Gardens
2.5. Ecosystem Service Flow between Gardens and Surrounding Agriculture
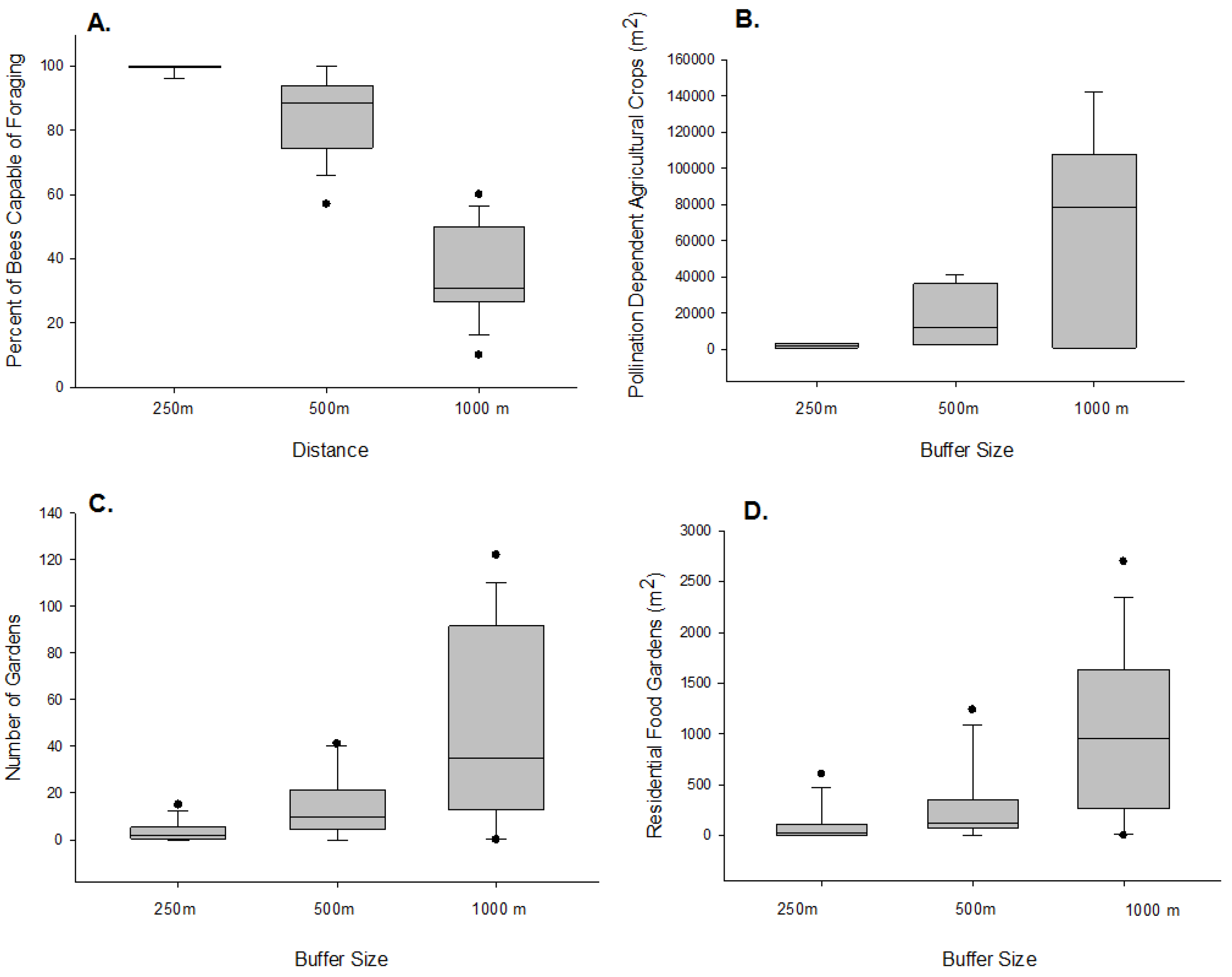
3. Results
4. Discussion
- What is the extent of pollinator movement between urban gardens and urban and peri-urban farms?
- Could management of urban bee habitat, in the form of residential gardens, parkland, and other managed green spaces within the city, positively influence pollinator movement between gardens and farms, to improve peri-urban and urban crop production?
- How far could the effects of urban land management extend out to agriculture in the surrounding peri-urban landscape, and what is the extent that current agriculture is benefitting from these services?
- How do effects of urban land management influence wild bees, versus managed bees?
- What is the spatial and temporal extent (i.e., quantity) and importance (i.e., quality) of urban gardens, relative to other forms of pollinator habitat (e.g., natural areas, other agricultural land) surrounding farms?
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Site # | Site Size (m2) | Max. Site Distance (m) | # of Bees Collected and Measured | Min.–Max. Foraging Range (m) of Bee Community | Land Area Planted to Pollination Dependent Commercial Crops (m2) | Land Area Planted as Gardens (m2) | Number of Gardens | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 250 m | 500 m | 1000 m | 250 m | 500 m | 1000 m | 250 m | 500 m | 1000 m | |||||
| 6 | 688 | 11 | 16 | 440–1120 | 0 | 0 | 0 | 19 | 59 | 329 | 1 | 7 | 19 |
| 9 | 20,234 | 60 | 51 | 190–1140 | 0 | 0 | 900 | n.c | n.c | n.c | n.c | n.c | n.c |
| 20 | 581 | 7 | 22 | 200–1270 | 0 | 0 | 0 | 606 | 1237 | 2697 | 15 | 39 | 73 |
| 22 | 1982 | 25 | 21 | 360–1030 | 0 | 0 | 0 | 62 | 99 | 230 | 3 | 5 | 15 |
| 35 | 486 | 9 | 12 | 350–1160 | 0 | 0 | 0 | 267 | 868 | 1,724 | 8 | 41 | 93 |
| 38 | 929 | 18 | 26 | 480–1250 | 3600 | 21,600 | 78,300 | n.c | n.c | n.c | n.c | n.c | n.c |
| 45 | 18,939 | 37 | 67 | 310–1120 | 1800 | 41,400 | 108,000 | 0 | 0 | 0 | 0 | 0 | 0 |
| 56 | 465 | 7 | 60 | 330–1070 | 0 | 0 | 0 | 153 | 403 | 953 | 8 | 23 | 61 |
| 58 | 445 | 5 | 16 | 270–1140 | 0 | 0 | 0 | 0 | 86 | 537 | 0 | 7 | 33 |
| 61 | 2670 | 29 | 41 | 460–1270 | 900 | 2,700 | 142,200 | n.c | n.c | n.c | n.c | n.c | n.c |
| 63 | 13,273 | 20 | 10 | 380–1080 | 0 | 0 | 8100 | 0 | 0 | 17 | 0 | 0 | 1 |
| 82 | 445 | 7 | 16 | 470–1130 | 0 | 0 | 0 | 27 | 304 | 1,270 | 2 | 20 | 91 |
| 88 | 1295 | 20 | 46 | 260–1190 | 0 | 0 | 0 | 5 | 99 | 298 | 1 | 4 | 11 |
| 91 | 51,894 | 135 | 48 | 210–1150 | 0 | 0 | 0 | 21 | 281 | 1,816 | 2 | 18 | 122 |
| 102 | 1740 | 22 | 22 | 460–1120 | 0 | 0 | 900 | n.c | n.c | n.c | n.c | n.c | n.c |
| 103 | 445 | 12 | 26 | 210–1110 | 0 | 0 | 0 | 47 | 120 | 1,074 | 2 | 10 | 35 |
| 104 | 167 | 8 | 36 | 350–1160 | 0 | 0 | 0 | 47 | 224 | 1,549 | 3 | 20 | 92 |
| 111 | 441 | 7 | 15 | 470–1110 | 0 | 2,700 | 81,000 | n.c | n.c | n.c | n.c | n.c | n.c |
References
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Ratnieks, F.L.W.; Carreck, N.L. Clarity on honey bee collapse? Science 2010, 327, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Brittain, C.; Potts, S.G. The potential impacts of insecticides on the life-history traits of bees and the consequences for pollination. Basic Appl. Ecol. 2011, 12, 321–331. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Kunin, W.E.; Keil, P.; Aguirre-Gutiérrez, J.; Ellis, W.N.; Fox, R.; Groom, Q.; Hennekens, S.; Van Landuyt, W.; Maes, D.; et al. Species richness declines and biotic homogenisation have slowed down for NW-European pollinators and plants. Ecol. Lett. 2013, 16, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Koh, I.; Lonsdorf, E.V.; Williams, N.M.; Brittain, C.; Isaacs, R.; Gibbs, J.; Ricketts, T.H. Modeling the status, trends, and impacts of wild bee abundance in the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Miles, A. Ecosystem services in biologically diversified versus conventional farming systems: Benefits, externalities, and trade-offs. Ecol. Soc. 2012, 17, 388–395. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Morandin, L.A.; Kremen, C. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Blaauw, B.R.; Isaacs, R. Larger patches of diverse floral resources increase insect pollinator density, diversity, and their pollination of native wildflowers. Basic Appl. Ecol. 2014, 15, 701–711. [Google Scholar] [CrossRef]
- Ponisio, L.C.; M’Gonigle, L.K.; Kremen, C. On-farm habitat restoration counters biotic homogenization in intensively managed agriculture. Glob. Chang. Biol. 2016, 22, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Park, M.G.; Blitzer, E.J.; Gibbs, J.; Losey, J.E.; Danforth, B.N. Negative effects of pesticides on wild bee communities can be buffered by landscape context. Proc. Natl. Acad. Sci. USA 2015, 282, 20150299. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.K.; Otieno, M.; Rajotte, E.G.; Fleischer, S.J.; Biddinger, D.J. Proximity to woodland and landscape structure drives pollinator visitation in apple orchard ecosystem. Front. Ecol. Evol. 2016, 4, 38. [Google Scholar] [CrossRef]
- Williams, N.M.; Ward, K.L.; Pope, N.; Isaacs, R.; Wilson, J.; May, E.A.; Ellis, J.; Daniels, J.; Pence, A.; Ullmann, K.; et al. Native wildflower plantings support wild bee abundance and diversity in agricultural landscapes across the United States. Ecol. Appl. 2015, 25, 2119–2131. [Google Scholar] [CrossRef] [PubMed]
- Francis, C.A.; Hansen, T.E.; Fox, A.A.; Hesje, P.J.; Nelson, H.E.; Lawseth, A.E.; English, A. Farmland conversion to non-agricultural uses in the US and Canada: Current impacts and concerns for the future. Int. J. Agric. Sustain. 2012, 10, 8–24. [Google Scholar] [CrossRef]
- Condon, P.M.; Mullinix, K.; Fallick, A.; Harcourt, M. Agriculture on the edge: Strategies to abate urban encroachment onto agricultural lands by promoting viable human-scale agriculture as an integral element of urbanization. Int. J. Agric. Sustain. 2010, 8, 104–115. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Fetridge, E.J.; Ascher, J.S.; Langellotto, G.A. The bee fauna of residential gardens in a suburb of New York City (Hymenoptera: Apoidea). Ann. Entomol. Soc. Am. 2008, 101, 1067–1077. [Google Scholar] [CrossRef]
- Matteson, K.C.; Ascher, J.S.; Langellotto, G.A. Bee richness and abundance in New York City urban gardens. Ann. Entomol. Soc. Am. 2008, 101, 140–150. [Google Scholar] [CrossRef]
- Frankie, G.W.; Thorp, R.W.; Pawelek, J.C.; Hernandez, J.; Coville, R. Urban bee diversity in a small residential garden in northern California. J. Hymenopt. Res. 2009, 18, 368–379. Available online: http://biostor.org/reference/111478 (accessed on 13 April 2018).
- Pawelek, J.C.; Frankie, G.W.; Thorp, R.W.; Przybylski, M. Modification of a community garden to attract native bee pollinators in urban San Luis Obispo, California. Cities Environ. 2009, 2. Article 7. Available online: http://digitalcommons.lmu.edu/cate/vol2/iss1/7 (accessed on 13 April 2018). [CrossRef]
- Pardee, G.L.; Philpott, S.M. Native plants are the bee’s knees: Local and landscape predictors of bee richness and abundance in backyard gardens. Urban Ecosyst. 2014, 17, 641–659. [Google Scholar] [CrossRef]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; Stone, G.N.; et al. Where Is the UK’s pollinator biodiversity? the importance of urban areas for flower-visiting insects. Proc. Biol. Sci. 2015, 282, 20142849. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, M.H.; Jackson, J.; Edwards, M.; Ollerton, J. Diversity and abundance of solitary and primitively eusocial bees in an urban centre: A case study from Northampton (England). J. Insect Conserv. 2015, 19, 487–500. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Shortall, C.R.; Todd, A.D.; Goulson, D.; Knight, M.E.; Hale, R.J.; Sanderson, R.A. Quantifying and comparing bumblebee nest densities in gardens and countryside habitats. J. Appl. Ecol. 2007, 45, 784–792. [Google Scholar] [CrossRef]
- Goulson, D.; Lepais, O.; O’Connor, S.; Osborne, J.L.; Sanderson, R.A.; Cussans, J.; Goffe, L.; Darvill, B. Effects of land use at a landscape scale on bumblebee nest density and survival. J. Appl. Ecol. 2010, 47, 1207–1215. [Google Scholar] [CrossRef]
- Wojcik, V.A.; McBride, J.R. Common factors influence bee foraging in urban and wildland landscapes. Urban Ecosyst. 2012, 15, 581–598. [Google Scholar] [CrossRef]
- Angold, P.G.; Sadler, J.P.; Hill, M.O.; Pullin, A.; Rushton, S.; Austin, K.; Small, E.; Wood, B.; Wadsworth, R.; Sanderson, R.; et al. Biodiversity in urban habitat patches. Sci. Total Environ. 2006, 360, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Hope, D.; Gries, C.; Zhu, W.; Fagan, W.F.; Redman, C.L.; Grimm, N.B.; Nelson, A.L.; Martin, C.; Kinzig, A. Socioeconomics drive urban plant diversity. Proc. Natl. Acad. Sci. USA 2003, 100, 8788–8792. [Google Scholar] [CrossRef] [PubMed]
- Grove, J.M.; Troy, A.R.; O’Neil-Dunne, J.P.M.; Burch, W.R.; Cadenasso, M.L.; Pickett, S.T.A. Characterization of households and its implications for the vegetation of urban ecosystems. Ecosystems 2006, 9, 578–597. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, L. The city in the country: Growing alternative food networks in metropolitan areas. J. Rural Stud. 2008, 24, 231–244. [Google Scholar] [CrossRef]
- Kaluza, B.F.; Wallace, H.; Heard, T.A.; Klein, A.-M.; Leonhardt, S.D. Urban gardens promote bee foraging over natural habitats and plantations. Ecol. Evol. 2016, 6, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Somme, L.; Moquet, L.; Quinet, M.; Vanderplanck, M.; Michez, D.; Lognay, G.; Jacquemart, A.-L. Food in a row: Urban trees offer valuable floral resources to pollinating insects. Urban Ecosyst. 2016, 19, 1149–1161. [Google Scholar] [CrossRef]
- David, A.; Botías, C.; Abdul-Sada, A.; Nicholls, E.; Rotheray, E.L.; Hill, E.M.; Goulson, D. Widespread contamination of wildflower and bee-collected pollen with complex mixtures of neonicotinoids and fungicides commonly applied to crops. Environ. Int. 2016, 88, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J.; Roulston, T.H.; Neal, M. Complex responses within a desert bee guild ( Hymenoptera: Apiformes) to urban habitat fragmentation. Ecol. Appl. 2006, 16, 632–644. [Google Scholar] [CrossRef]
- Carré, G.; Roche, P.; Chifflet, R.; Morison, N.; Bommarco, R.; Harrison-Cripps, J.; Krewenka, K.; Potts, S.G.; Roberts, S.P.M.; Rodet, G.; et al. Landscape context and habitat type as drivers of bee diversity in European annual crops. Agric. Ecosyst. Environ. 2009, 133, 40–47. [Google Scholar] [CrossRef]
- Matteson, K.C.; Langellotto, G.A. Bumble bee abundance in New York City community gardens: Implications for urban agriculture. Cities Environ. 2009, 2. Article 5. Available online: http://digitalcommons.lmu.edu/cate/vol2/iss1/5 (accessed on 13 April 2018). [CrossRef]
- Lowenstein, D.M.; Matteson, K.C.; Minor, E.S. Diversity of wild bees supports pollination services in an urbanized landscape. Oecologia 2015, 179, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Potter, A.; LeBuhn, G. Pollination service to urban agriculture in San Francisco, CA. Urban Ecosyst. 2015, 18, 885–893. [Google Scholar] [CrossRef]
- Ackerman, K.; Conard, M.; Culligan, P.; Plunz, R.; Sutto, M.-P.; Whittinghill, L. Sustainable food systems for future cities: The potential of urban agriculture. Econ. Soc. Rev. 2014, 45, 189–206. Available online: http://www.esr.ie/article/view/136 (accessed on 13 April 2018).
- Moore, L.J.; Kosut, M. Buzz: Urban Beekeeping and the Power of the Bee; New York University Press: New York, NY, USA; London, UK, 2013. [Google Scholar]
- Edmunds, B.; Little, R.; Sagilli, R. Nurturing Mason Bees in Your Backyard in Western Oregon; Extension Service: Corvallis, OR, USA, 2016. [Google Scholar]
- Kreyer, D.; Oed, A.; Walther-Hellwig, K.; Frankl, R. Are forests potential landscape barriers for foraging bumblebees? Landscape scale experiments with Bombus terrestris agg. and Bombus pascuorum (Hymenoptera, Apidae). Biol. Conserv. 2004, 116, 111–118. [Google Scholar] [CrossRef]
- Krewenka, K.M.; Holzschuh, A.; Tscharntke, T.; Dormann, C.F. Landscape elements as potential barriers and corriders for bees, wasps and parasitoids. Biol. Conserv. 2011, 144, 1816–1825. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Primack, R.B.; Gerwing, J. Are roads and railroads barriers to bumblebee movement in a temperate suburban conservation area? Biol. Conserv. 2003, 109, 37–45. [Google Scholar] [CrossRef]
- McClintock, N.; Young, J.; Santos, J.; Evans, T.; Simpson, M. Periodic atlas of the metroscape: Food production. In Metroscape; Institute of Portland Metropolitan Studies, Portland State University: Portland, OR, USA, 2013; pp. 13–19. [Google Scholar]
- Zillow. Available online: https://www.zillow.com/ (accessed on 3 April 2018).
- Cane, J.H. Estimation of bee body size using intertegula span (Apoidea). J. Kansas Entomol. Soc. 1987, 60, 145–147. [Google Scholar]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee foraging ranges and their relationship to body size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Environmental Systems Research Institute, (ESRI). ArcGIS Release 10.6; Environmental Systems Research Institute, (ESRI): Redlands, CA, USA, 2018. [Google Scholar]
- USDA National Agricultural Statistics Service Cropland Data Layer; USDA-NASS: Washington, DC, USA, 2018. Available online: https://nassgeodata.gmu.edu/CropScape/ (accessed on 14 March 2018).
- Kleijn, D.; Winfree, R.; Bartomeus, I.; Carvalheiro, L.G.; Henry, M.; Isaacs, R.; Klein, A.M.; Kremen, C.; M’Gonigle, L.K.; Rader, R.; et al. Delivery of crop pollination services is an insufficient argument for wild pollinator conservation. Nat. Commun. 2015, 6, 7414. [Google Scholar] [CrossRef] [PubMed]
- Oberholtzer, L.; Dimitri, C.; Pressman, A. Urban agriculture in the United States: Characteristics, challenges, and technical assistance needs. J. Ext. 2014, 52, 6FEA1. Available online: https://joe.org/joe/2014december/a1.php (accessed on 16 April 2018).
- Taylor, J.R.; Lovell, S.T. Mapping public and private spaces of urban agriculture in Chicago through the analysis of high-resolution aerial images in Google Earth. Landsc. Urban Plan. 2012, 108, 57–70. [Google Scholar] [CrossRef]
- McClintock, N.; Mahmoudi, D.; Simpson, M.; Santos, J.P. Socio-spatial differentiation in the sustainable city: A mixed-methods assessment of residential gardens in metropolitan Portland, Oregon, USA. Landsc. Urban Plan. 2016, 148, 1–16. [Google Scholar] [CrossRef]
- Harvey, T.; Works, M.A. Urban sprawl and rural landscapes: Perceptions of landscape as amenity in Portland, Oregon. Local Environ. 2002, 7, 381–396. [Google Scholar] [CrossRef]
- Chun, N. An emerging contradiction: Non-farm activity within exclusive farm use zones. In Metroscap; Institute of Portland Metropolitan Studies, Portland State University: Portland, OR, USA, 2018; Available online: https://metroscape.imspdx.org/an-emerging-contradiction-non-farm-activity-within-exclusive-farm-use-zones (accessed on 16 April 2018).
- Schtickzelle, N.; Baguette, M. Behavioural responses to habitat patch boundaries restrict dispersal and generate emigration-patch area relationships in fragmented landscapes. J. Anim. Ecol. 2003, 72, 533–545. [Google Scholar] [CrossRef]
- Krauss, J.; Alfert, T.; Steffan-Dewenter, I. Habitat area but not habitat age determines wild bee richness in limestone quarries. J. Appl. Ecol. 2009, 46, 194–202. [Google Scholar] [CrossRef]
- Winfree, R.; Dushoff, J.; Crone, E.E.; Schultz, C.B.; Budny, R.V.; Williams, N.M.; Kremen, C. Testing simple indices of habitat proximity. Am. Nat. 2005, 165, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Bugg, R.L.; Fay, J.P.; Thorp, R.W. The area requirements of an ecosystem service: Crop pollination by native bee communities in California. Ecol. Lett. 2004, 7, 1109–1119. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum foraging ranges in solitary bees: Only few individuals have the capability to cover long foraging distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]
- Beekman, M.; Ratnieks, F.L.W. Long-range foraging by the honey-bee, Apis mellifera L. Funct. Ecol. 2000, 14, 490–496. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Carreck, N.L.; Swain, J.L.; Knight, M.E.; Goulson, D.; Hale, R.J.; Sanderson, R.A. Bumblebee flight distances in relation to the forage landscape. J. Anim. Ecol. 2008, 77, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Leong, M.; Ponisio, L.C.; Kremen, C.; Thorp, R.W.; Roderick, G.K. Temporal dynamics influenced by global change: Bee community phenology in urban, agricultural, and natural landscapes. Glob. Chang. Biol. 2016, 22, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- McFrederick, W.S.; LeBuhn, G. Are urban parks refuges for bumble bees Bombus spp. (Hymenoptera: Apidae)? Biol. Conserv. 2006, 129, 372–382. [Google Scholar] [CrossRef]
- Ahrné, K.; Bengtsson, J.; Elmqvist, T. Bumble bees (Bombus spp.) along a gradient of increasing urbanization. PLoS ONE 2009, 4, e5574. [Google Scholar] [CrossRef] [PubMed]
- Melathopoulos, A.P.; Rodia, R.; Holt, J.; Sagili, R. Residential Beekeeping: Best-practice Guidelines for Nuisance-free Beekeeping in Oregon; Oregon State University: Corvallis, OR, USA, 2018; EM9186. [Google Scholar]
- Winter Bee Losses of Oregon Backyard Beekeepers, 2018. Available online: http://www.webcitation.org/706hgm01U (accessed on 11 June 2018).
- Matteson, K.C.; Langellotto, G.A. Evaluating community gardens as habitat for an urban butterfly. Cities Environ. 2012, 5. Article 10. Available online: http://digitalcommons.lmu.edu/cate/vol5/iss1/10 (accessed on 16 April 2018). [CrossRef]
- Hennig, E.I.; Ghazoul, J. Pollinating animals in the urban environment. Urban Ecosyst. 2012, 15, 149–166. [Google Scholar] [CrossRef]
- Heinrich, B. Do bumblebees forage optimally, and does it matter? Am. Zool. 1983, 23, 273–281. [Google Scholar] [CrossRef]
- Hicks, D.M.; Ouvrard, P.; Baldock, K.C.R.; Baude, M.; Goddard, M.A.; Kunin, W.E.; Mitschunas, N.; Memmott, J.; Morse, H.; Nikolitsi, M.; et al. Food for pollinators: Quantifying the nectar and pollen resources of urban flower meadows. PLoS ONE 2016, 11, e0158117. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrné, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef] [PubMed]
- López-Uribe, M.M.; Oi, C.A.; Del Lama, M.A. Nectar-foraging behavior of Euglossine bees (Hymenoptera: Apidae) in urban areas. Apidologie 2008, 39, 410–418. [Google Scholar] [CrossRef]
- Real, L.A. Uncertainty and pollinator-plant interactions: The foraging behavior of bees and wasps on artificial flowers. Ecology 1981, 62, 20–26. [Google Scholar] [CrossRef]
- Bennett, J.A.; Gensler, G.C.; Cahill, J.F., Jr. Small-scale bee patch use is affected equally by flower availability and local habitat configuration. Basic Appl. Ecol. 2014, 15, 260–268. [Google Scholar] [CrossRef]
- Sharp, J.; Smith, M. Social capital and farming at the rural–urban interface: The importance of nonfarmer and farmer relations. Agric. Syst. 2003, 76, 913–927. [Google Scholar] [CrossRef]
- Hanes, S.P.; Collum, K.K.; Hoshide, A.K.; Asare, E. Grower perceptions of native pollinators and pollination strategies in the lowbush blueberry industry. Renew. Agric. Food Syst. 2015, 30, 124–131. [Google Scholar] [CrossRef]
- Wilson, J.S.; Forister, M.L.; Carril, O.M. Interest exceeds understanding in public support of bee conservation. Front. Ecol. Environ. 2017, 15, 460–466. [Google Scholar] [CrossRef]
- Wu, J.; Jenerette, G.D.; Buyantuyev, A.; Redman, C.L. Quantifying spatiotemporal patterns of urbanization: The case of the two fastest growing metropolitan regions in the United States. Ecol. Complex. 2011, 8, 1–8. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Müller, N.; Ignatieva, M.; Nilon, C.H.; Werner, P.; Zipperer, W.C. Patterns and Trends in Urban Biodiversity and Landscape Design. In Urbanization, Biodiversity and Ecosystem Services: Challenges and Opportunities; Springer: Dordrecht, The Netherlands, 2013; pp. 123–174. ISBN 978-94-007-7088-1. [Google Scholar]
- McKinney, M.L. Urbanization, biodiversity, and conservation: The impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Kareiva, P.; Watts, S.; McDonald, R.; Boucher, T. Domesticated nature: Shaping landscapes and ecosystems for human welfare. Science 2007, 316, 1866–1869. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Langellotto, G.A.; Melathopoulos, A.; Messer, I.; Anderson, A.; McClintock, N.; Costner, L. Garden Pollinators and the Potential for Ecosystem Service Flow to Urban and Peri-Urban Agriculture. Sustainability 2018, 10, 2047. https://doi.org/10.3390/su10062047
Langellotto GA, Melathopoulos A, Messer I, Anderson A, McClintock N, Costner L. Garden Pollinators and the Potential for Ecosystem Service Flow to Urban and Peri-Urban Agriculture. Sustainability. 2018; 10(6):2047. https://doi.org/10.3390/su10062047
Chicago/Turabian StyleLangellotto, Gail Ann, Andony Melathopoulos, Isabella Messer, Aaron Anderson, Nathan McClintock, and Lucas Costner. 2018. "Garden Pollinators and the Potential for Ecosystem Service Flow to Urban and Peri-Urban Agriculture" Sustainability 10, no. 6: 2047. https://doi.org/10.3390/su10062047
APA StyleLangellotto, G. A., Melathopoulos, A., Messer, I., Anderson, A., McClintock, N., & Costner, L. (2018). Garden Pollinators and the Potential for Ecosystem Service Flow to Urban and Peri-Urban Agriculture. Sustainability, 10(6), 2047. https://doi.org/10.3390/su10062047