Unique Bee Communities within Vacant Lots and Urban Farms Result from Variation in Surrounding Urbanization Intensity


Abstract
1. Introduction
2. Methods
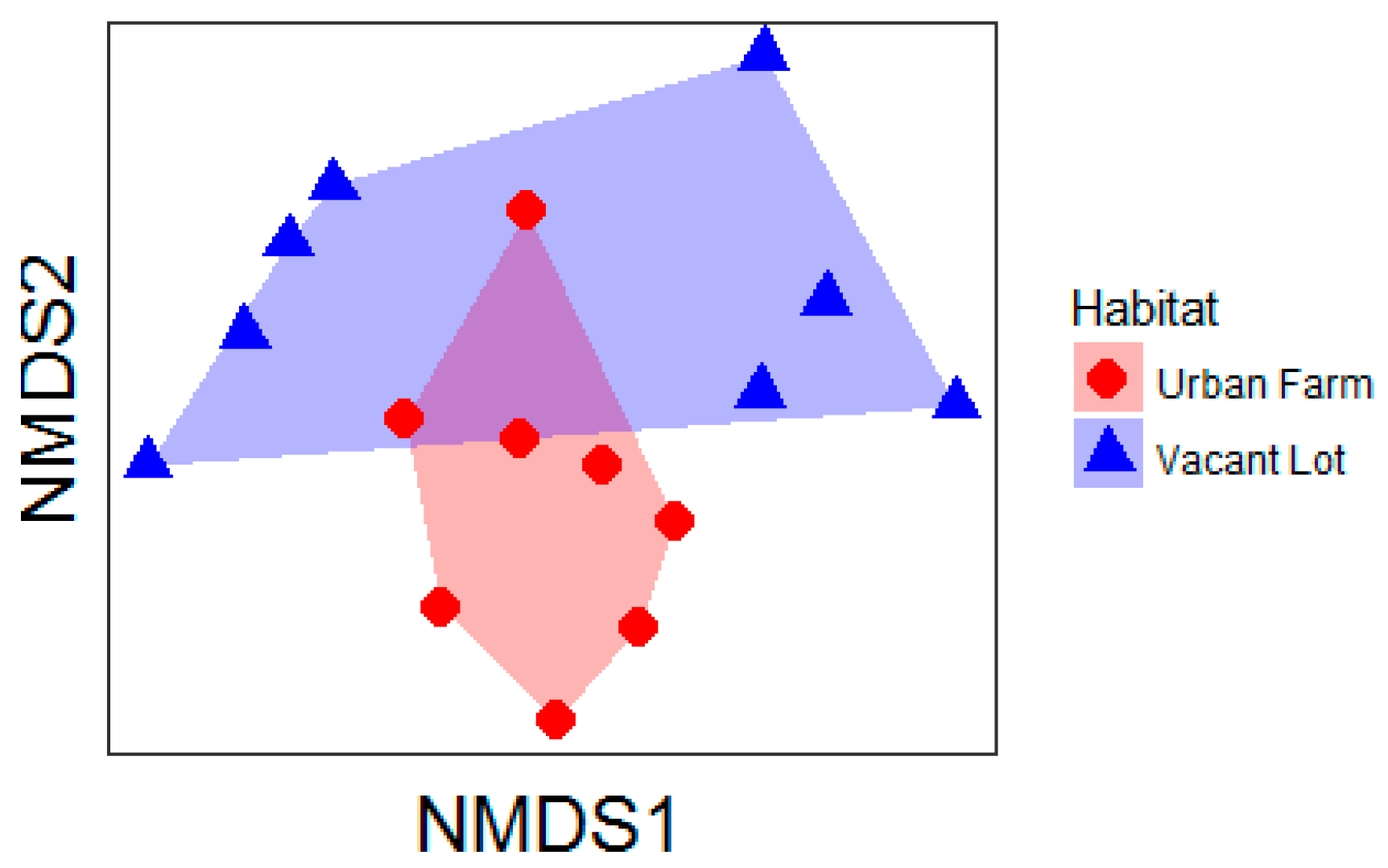
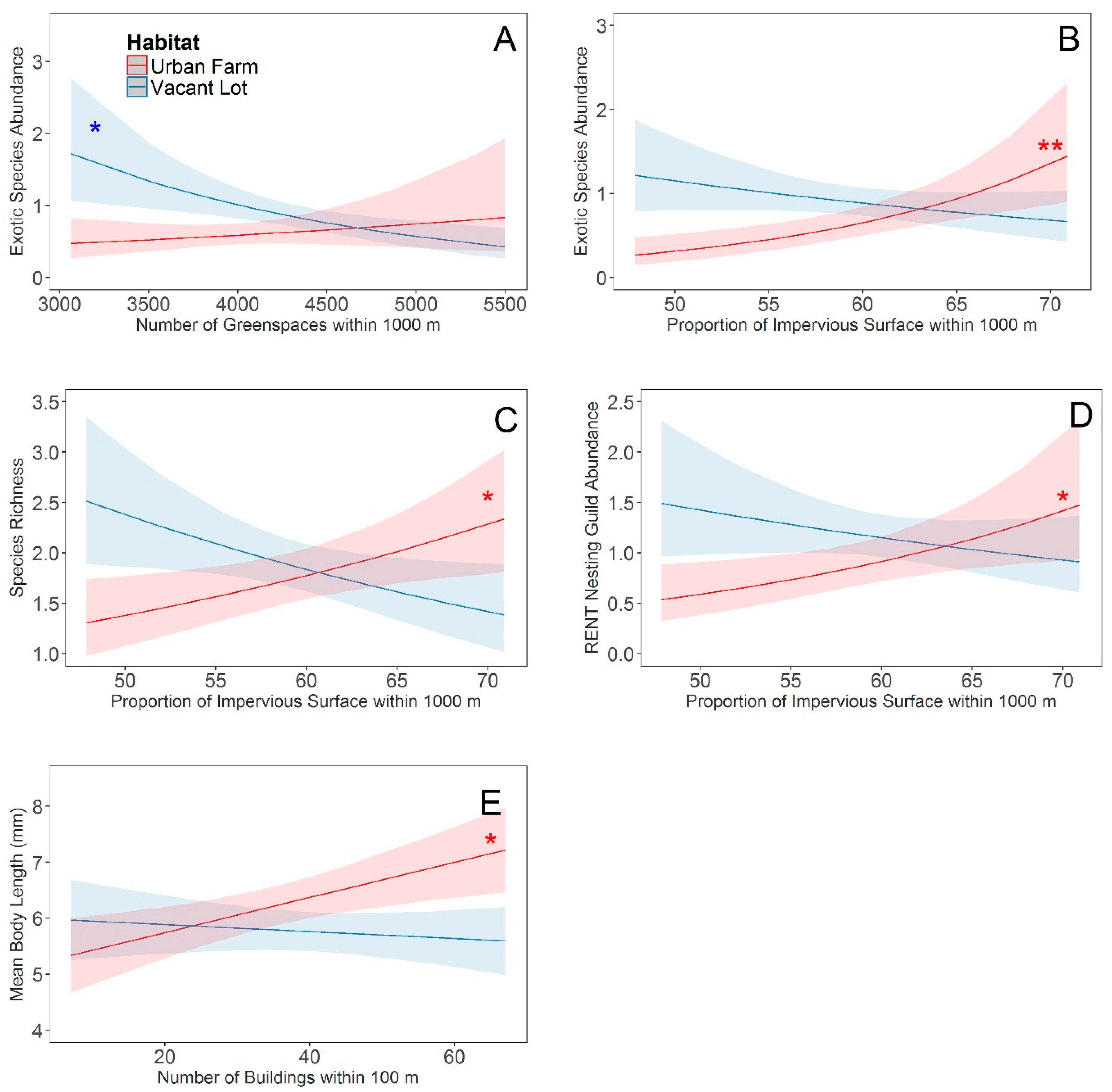
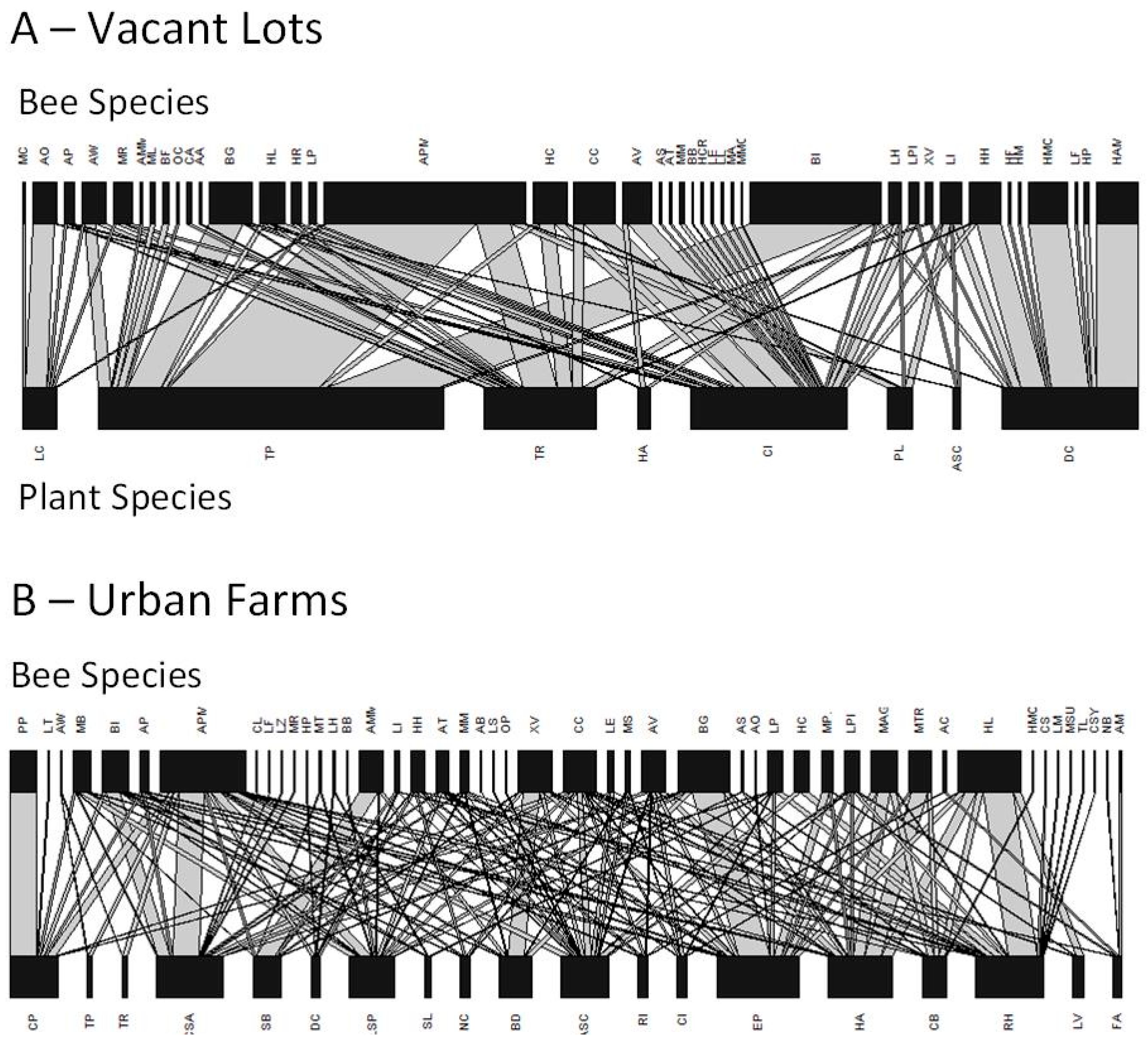
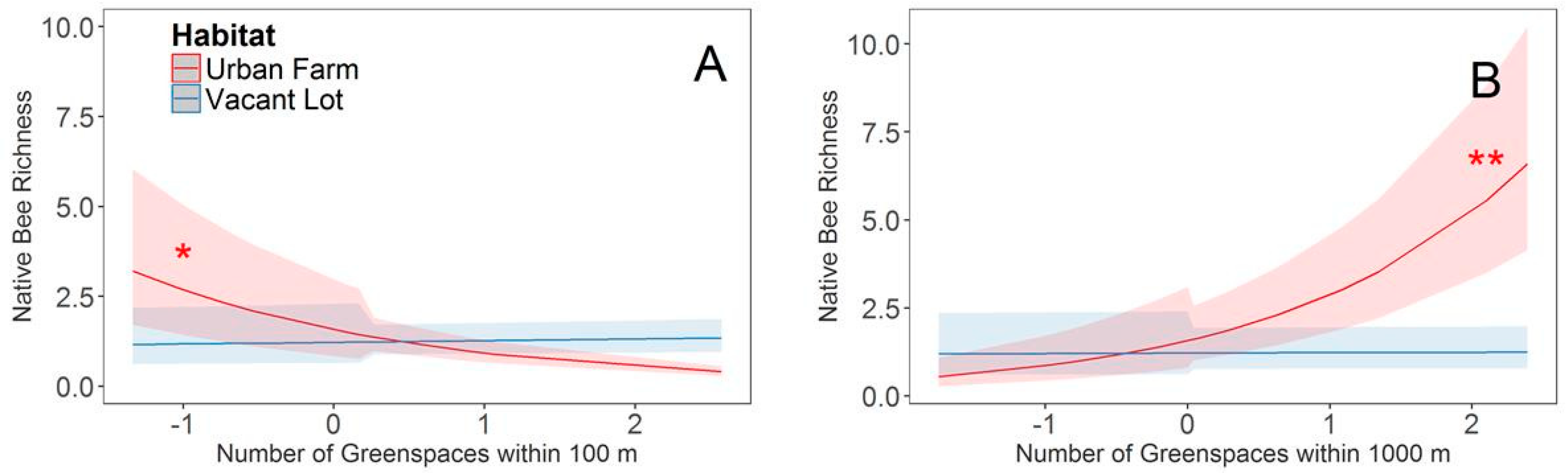
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Species Name | Nesting Guild a | Origin b |
|---|---|---|---|
| AA | Augochlorella aurata | MINER | Native |
| AB | Andrena barbilobris | MINER | Native |
| AC | Andrena commoda | MINER | Native |
| AM | Andrena miserabilis | MINER | Native |
| AMM | Anthidium manicatum | MINER | Exotic |
| AO | Anthidium oblongatum | MINER | Exotic |
| AP | Augochlora pura | RENT | Native |
| APM | Apis mellifera | RENT | Exotic |
| AS | Agapostemon sericeus | MINER | Native |
| AT | Agapostemon texanus | MINER | Native |
| AV | Agapostemon virescens | MINER | Native |
| AV | Andrena vicina | MINER | Native |
| AW | Andrena wilkella | MINER | Exotic |
| BB | Bombus bimaculatus | RENT | Native |
| BF | Bombus fervidus | RENT | Native |
| BG | Bombus griseocollis | RENT | Native |
| BI | Bombus impatiens | RENT | Native |
| CA | Calliopsis andreniformis | MINER | Native |
| CC | Ceratina calcarata | PITH | Native |
| CL | Colletes latitarsis | MINER | Native |
| CS | Ceratina stenua | PITH | Native |
| CSY | Coelioxys sayi | NONEST | Native |
| HAM | Hylaeus addinis/modestus | RENT | Native |
| HC | Halictus confusus | MINER | Native |
| HCR | Heriades carinata | RENT | Native |
| HF | Hylaeus fedorica | RENT | Native |
| HH | Hylaeus hyalinatus | RENT | Exotic |
| HL | Halictus ligatus | MINER | Native |
| HM | Hylaeus mesillae | RENT | Native |
| HMO | Hylaeus modestus | RENT | Native |
| HP | Hylaeus pictipes | RENT | Exotic |
| HR | Halictus rubicundus | MINER | Native |
| LE | Lasioglossum ephialtum | MINER | Native |
| LF | Lasioglossum fuscipenne | MINER | Native |
| LH | Lasioglossum hitchensi | MINER | Native |
| LI | Lasioglossum imitatum | MINER | Native |
| LL | Lasioglossum leucozonium | MINER | Native |
| LM | Lasioglossum mendica | MINER | Native |
| LP | Lasioglossum pectorale | MINER | Native |
| LPI | Lasioglossum pilosum | MINER | Native |
| LS | Lasioglossum smilacinae | MINER | Native |
| LT | Lasioglossum tegulare | MINER | Native |
| LZ | Lasioglossum zonulum | MINER | Native |
| MA | Megachile apicalis | RENT | Exotic |
| MAG | Melissodes agilis | MINER | Native |
| MB | Melissodes bimaculata | MINER | Native |
| MC | Megachile concinna | RENT | Native |
| ML | Megachile latimanus | RENT | Native |
| MM | Megachile mendica | RENT | Native |
| MMO | Megachile montivaga | RENT | Native |
| MP | Megachile pugnata | RENT | Native |
| MR | Megachile rotundata | RENT | Exotic |
| MS | Megachile sculpturalis | RENT | Exotic |
| MSU | Melissodes subillatas | MINER | Native |
| MT | Megachile texana | RENT | Native |
| MTR | Melissodes trinodis | MINER | Native |
| NB | Nomada bidentate grp | NONEST | Native |
| OC | Osmia caerulescens | RENT | Exotic |
| OP | Osmia pumila | RENT | Native |
| PP | Peponapis pruinosa | MINER | Native |
| TL | Triepeolus lunatus | NONEST | Native |
| XV | Xylocopa virginica | CARPENTER | Native |
| Code | Species Name | Common Name |
|---|---|---|
| AS | Allium schoenoprasum | Chives |
| ASY | Asclepias syriaca | Common Milkweed |
| BD | Buddleja davidii | Butterfly Bush |
| CB | Cosmos bipinnatus | Cosmos |
| CI | Cichorium intybus | Chicory |
| CP | Cucurbita pepo | Squash |
| CS | Cucumis sativus | Cucumber |
| DC | Daucus carota | Queen Ann’s Lace |
| EP | Echinacea purpurea | Purple Coneflower |
| FA | Fragaria x ananassa | Strawberry |
| HA | Helianthus annuus | Sunflower |
| LC | Lotus corniculatus | birds-foot trefoil |
| LS | Lavandula spp. | Lavender |
| LV | Leucanthemum vulgare | Oxeye daisy |
| NC | Nepeta cataria | Catnip |
| PL | Plantago lanceolata | Narrowleaf Plantain |
| RH | Rudbeckia hirta | Black-eyed Susan |
| RI | Rubus idaeus | Raspberry |
| SB | Stachys byzantina | Lamb’s Ear |
| SL | Solanum lycopersicum | Tomato |
| TP | Trifolium pratense | Red Clover |
| TR | Trifolium repens | White Clover |
References
- Lepczyk, C.A.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S. Biodiversity in the City: Fundamental Questions for Understanding the Ecology of Urban Green Spaces for Biodiversity Conservation. BioScience 2017, 67, 799–807. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrné, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Aronson, M.F.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.; Cilliers, S.; Clarkson, B. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Banaszak-Cibicka, W.; Ratyńska, H.; Dylewski, L. Features of urban green space favourable for large and diverse bee populations (Hymenoptera: Apoidea: Apiformes). Urban For. Urban Green. 2016, 20, 448–452. [Google Scholar] [CrossRef]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.; Rayner, L. Cities are hotspots for threatened species. Glob. Ecol. Biogeogr. 2016, 25, 117–126. [Google Scholar] [CrossRef]
- Gardiner, M.M.; Burkman, C.E.; Prajzner, S.P. The Value of Urban Vacant Land to Support Arthropod Biodiversity and Ecosystem Services. Environ. Entomol. 2013, 42, 1123–1136. [Google Scholar] [CrossRef] [PubMed]
- Potter, A.; LeBuhn, G. Pollination service to urban agriculture in San Francisco, CA. Urban Ecosyst. 2015, 18, 885–893. [Google Scholar] [CrossRef]
- Burkman, C.E.; Gardiner, M.M. Urban greenspace composition and landscape context influence natural enemy community composition and function. Biol. Control 2014, 75, 58–67. [Google Scholar] [CrossRef]
- Gardiner, M.M.; Prajzner, S.P.; Burkman, C.E.; Albro, S.; Grewal, P.S. Vacant land conversion to community gardens: Influences on generalist arthropod predators and biocontrol services in urban greenspaces. Urban Ecosyst. 2014, 17, 101–122. [Google Scholar] [CrossRef]
- Riley, C.B.; Herms, D.A.; Gardiner, M.M. Exotic trees contribute to urban forest diversity and ecosystem services in inner-city Cleveland, OH. Urban For. Urban Green. 2018, 29, 367–376. [Google Scholar] [CrossRef]
- Delgado de la Flor, Y.A.; Burkman, C.E.; Eldredge, T.K.; Gardiner, M.M. Patch and landscape-scale variables influence the taxonomic and functional composition of beetles in urban greenspaces. Ecosphere 2017, 8, e02007. [Google Scholar] [CrossRef]
- McFrederick, Q.S.; LeBuhn, G. Are urban parks refuges for bumble bees Bombus spp. (Hymenoptera: Apidae)? Biol. Conserv. 2006, 129, 372–382. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- HilleRisLambers, J.; Adler, P.B.; Harpole, W.S.; Levine, J.M.; Mayfield, M.M. Rethinking Community assembly through the lens of coexistence theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 227–248. [Google Scholar] [CrossRef]
- Aronson, M.F.; Nilon, C.H.; Lepczyk, C.A.; Parker, T.S.; Warren, P.S.; Cilliers, S.S.; Goddard, M.A.; Hahs, A.K.; Herzog, C.; Katti, M. Hierarchical filters determine community assembly of urban species pools. Ecology 2016, 97, 2952–2963. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Harrison, T.; Winfree, R. Urban drivers of plant-pollinator interactions. Funct. Ecol. 2015, 29, 879–888. [Google Scholar] [CrossRef]
- Antonini, Y.; Martins, R.P.; Aguiar, L.M.; Loyola, R.D. Richness, composition and trophic niche of stingless bee assemblages in urban forest remnants. Urban Ecosyst. 2013, 16, 527–541. [Google Scholar] [CrossRef]
- Braaker, S.; Ghazoul, J.; Obrist, M.K.; Moretti, M. Habitat connectivity shapes urban arthropod communities: The key role of green roofs. Ecology 2014, 95, 1010–1021. [Google Scholar] [CrossRef] [PubMed]
- Quistberg, R.D.; Bichier, P.; Philpott, S.M. Landscape and local correlates of bee abundance and species richness in urban gardens. Environ. Entomol. 2016, 45, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Wray, J.C.; Neame, L.A.; Elle, E. Floral resources, body size, and surrounding landscape influence bee community assemblages in oak-savannah fragments. Ecol. Entomol. 2014, 39, 83–93. [Google Scholar] [CrossRef]
- Banaszak-Cibicka, W.; Żmihorski, M. Wild bees along an urban gradient: Winners and losers. J. Insect Conserv. 2012, 16, 331–343. [Google Scholar] [CrossRef]
- Pardee, G.L.; Philpott, S.M. Native plants are the bee’s knees: Local and landscape predictors of bee richness and abundance in backyard gardens. Urban Ecosyst. 2014, 17, 641–659. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Matteson, K.C.; Xiao, I.; Silva, A.M.; Minor, E.S. Humans, bees, and pollination services in the city: The case of Chicago, IL (USA). Biodivers. Conserv. 2014, 23, 2857–2874. [Google Scholar] [CrossRef]
- Fischer, L.K.; Eichfeld, J.; Kowarik, I.; Buchholz, S. Disentangling urban habitat and matrix effects on wild bee species. PeerJ 2016, 4, e2729. [Google Scholar] [CrossRef] [PubMed]
- Frankie, G.W.; Thorp, R.W.; Pawelek, J.C.; Hernandez, J.; Coville, R. Urban bee diversity in a small residential garden in northern California. J. Hymenopt. Res. 2009, 18, 368–379. [Google Scholar]
- Garbuzov, M.; Samuelson, E.E.; Ratnieks, F.L. Survey of insect visitation of ornamental flowers in Southover Grange garden, Lewes, UK. Insect Sci. 2015, 22, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Sikora, A.; Micholap, P.; Kelm, M. Flowering plants preferred by bumblebees (Bombus Latr.) in the botanical garden of medicinal plants in Wroclaw. J. Apic. Sci. 2016, 60, 59–68. [Google Scholar] [CrossRef]
- Matteson, K.C.; Grace, J.B.; Minor, E.S. Direct and indirect effects of land use on floral resources and flower-visiting insects across an urban landscape. Oikos 2013, 122, 682–694. [Google Scholar] [CrossRef]
- Hinners, S.J.; Hjelmroos-Koski, M.K. Receptiveness of foraging wild bees to exotic landscape elements. Am. Midland Nat. 2009, 162, 253–265. [Google Scholar] [CrossRef]
- MacIvor, J.S.; Ruttan, A.; Salehi, B. Exotics on exotics: Pollen analysis of urban bees visiting Sedum on a green roof. Urban Ecosyst. 2015, 18, 419–430. [Google Scholar] [CrossRef]
- Larson, J.L.; Kesheimer, A.J.; Potter, D.A. Pollinator assemblages on dandelions and white clover in urban and suburban lawns. J. Insect Conserv. 2014, 18, 863–873. [Google Scholar] [CrossRef]
- Hanley, M.E.; Awbi, A.J.; Franco, M. Going native? Flower use by bumblebees in English urban gardens. Ann. Bot. 2014, 113, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Hicks, D.M.; Ouvrard, P.; Baldock, K.C.R.; Baude, M.; Goddard, M.A.; Kunin, W.E.; Mitschunas, N.; Memmott, J.; Morse, H.; Nikolitsi, M.; et al. Food for pollinators: Quantifying the nectar and pollen resources of urban flower meadows. PLoS ONE 2016, 11, e0158117. [Google Scholar] [CrossRef] [PubMed]
- Frankie, G.W.; Thorp, R.W.; Schindler, M.; Hernandez, J.; Ertter, B.; Rizzardi, M. Ecological patterns of bees and their host ornamental flowers in two northern California cities. J. Kansas Entomol. Soc. 2005, 78, 227–246. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Walker, K.; Williams, N.S.; Hahs, A.K.; Mata, L.; Stork, N.; Livesley, S.J. The conservation value of urban green space habitats for Australian native bee communities. Biol. Conserv. 2015, 187, 240–248. [Google Scholar] [CrossRef]
- Matteson, K.C.; Langellotto, G.A. Determinates of inner city butterfly and bee species richness. Urban Ecosyst. 2010, 13, 333–347. [Google Scholar] [CrossRef]
- Oswalt, P.; Rieniets, T. Atlas of Shrinking Cities; Hatje Cantz: Berlin, Germany, 2006. [Google Scholar]
- Greene, J. (Commissioner of Neighborhood Development, Cleveland, OH, USA). Personal communication, 2008.
- Droege, S. The Very Handy Manual: How to Catch and Identify Bees and Manage a Collection; USGS Bee Inventory and Monitoring Lab, Patuxent Wildlife Research Center: Beltsville, MA, USA, 2009. [Google Scholar]
- Ascher, J.S.; Pickering, J. Bee Species Guide (Hymenoptera: Apoidea: Anthophila). Available online: http://www.discoverlife.org/mp/20q (accessed on 7 June 2018).
- Moretti, M.; de Bello, F.; Roberts, S.P.M.; Potts, S.G. Taxonomical vs. functional responses of bee communities to fire in two contrasting climatic regions. J Anim. Ecol. 2009, 78, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.H.; Knott, B.; Eberlein, T.; Memmott, J. Sampling method influences the structure of plant–pollinator networks. Oikos 2011, 120, 822–831. [Google Scholar] [CrossRef]
- Blaschke, T. Object based image analysis for remote sensing. ISPRS J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef]
- De Pinho, C.M.D.; Fonseca, L.M.G.; Korting, T.S.; Almeida, C.M.; de Kux, H.J.H. Land-cover classification of an intra-urban environment using high-resolution images and object-based image analysis. Int. J. Remote Sens. 2012, 33, 5973–5995. [Google Scholar] [CrossRef]
- Zhou, W.; Troy, A. An object-oriented approach for analysing and characterizing urban landscape at the parcel level. Int. J. Remote Sens. 2008, 29, 3119–3135. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10.1; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Trimble Documentation. Trimble eCognition Developer 8.7 Reference; Trimble Germany GmbH: Munich, Germany, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv, 2014; arXiv:1406.5823. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R package Version 1.17-2; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite package: Analysing ecological networks. R News 2008, 8, 8–11. [Google Scholar]
- Dormann, C.F.; Fründ, J.; Blüthgen, N.; Gruber, B. Indices, graphs and null models: Analyzing bipartite ecological networks. Open Ecol. J. 2009, 2, 7–24. [Google Scholar] [CrossRef]
- Dormann, C.F. How to be a specialist? Quantifying specialisation in pollination networks. Netw. Biol. Hong Kong 2011, 1, 1. [Google Scholar] [CrossRef]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissière, B.E. Decreasing abundance, increasing diversity and changing structure of the wild bee community (Hymenoptera: Anthophila) along an urbanization gradient. PLoS ONE 2014, 9, e104679. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, B.C.; Shuster, W.D.; Garmestani, A.S.; Furio, B.; Albro, S.L.; Gardiner, M.; Spring, M.; Green, O.O. A tale of two rain gardens: Barriers and bridges to adaptive management of urban stormwater in Cleveland, Ohio. J. Environ. Manag. 2016, 183, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Cameron, R.W.; Blanuša, T. Green infrastructure and ecosystem services–is the devil in the detail? Ann. Bot. 2016, 118, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Matteson, K.C.; Ascher, J.S.; Langellotto, G.A. Bee richness and abundance in New York City urban gardens. Ann. Entomol. Soc. Am. 2008, 101, 140–150. [Google Scholar] [CrossRef]
- Tonietto, R.; Fant, J.; Ascher, J.; Ellis, K.; Larkin, D. A comparison of bee communities of Chicago green roofs, parks and prairies. Landsc. Urban Plan. 2011, 103, 102–108. [Google Scholar] [CrossRef]
- Burkman, C.E.; Gardiner, M.M. Spider assemblages within greenspaces of a deindustrialized urban landscape. Urban Ecosyst. 2015, 18, 793–818. [Google Scholar] [CrossRef]
- Uno, S.; Cotton, J.; Philpott, S.M. Diversity, abundance, and species composition of ants in urban green spaces. Urban Ecosyst. 2010, 13, 425–441. [Google Scholar] [CrossRef]
- Sheffield, C.S.; Pindar, A.; Packer, L.; Kevan, P.G. The potential of cleptoparasitic bees as indicator taxa for assessing bee communities. Apidologie 2013, 44, 501–510. [Google Scholar] [CrossRef]
- Richards, M.H.; Packer, L. Annual variation in survival and reproduction of the primitively eusocial sweat bee Halictus ligatus (Hymenoptera: Halictidae). Can. J. Zool. 1995, 73, 933–941. [Google Scholar] [CrossRef]
- Gibbs, J.; Brady, S.G.; Kanda, K.; Danforth, B.N. Phylogeny of halictine bees supports a shared origin of eusociality for Halictus and Lasioglossum (Apoidea: Anthophila: Halictidae). Mol. Phylogenet. Evol. 2012, 65, 926–939. [Google Scholar] [CrossRef] [PubMed]
- Onuferko, T.M.; Kutby, R.; Richards, M.H. A list of bee species (Hymenoptera: Apoidea) recorded from three municipalities in the Niagara region of Ontario, including a new record of Lasioglossum furunculum Gibbs (Halictidae) in Canada. J. Entomol. Soc. Ont. 2015, 146, 3–22. [Google Scholar]
- MacIvor, J.S.; Cabral, J.M.; Packer, L. Pollen specialization by solitary bees in an urban landscape. Urban Ecosyst. 2014, 17, 139–147. [Google Scholar] [CrossRef]
- Nicholls, C.I.; Altieri, M.A. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef]
- Ahrne, K.; Bengtsson, J.; Elmqvist, T. Bumble bees (Bombus spp.) along a gradient of increasing urbanization. PLoS ONE 2009, 4, e5574. [Google Scholar] [CrossRef] [PubMed]
- Garbuzov, M.; Ratnieks, F.L. Quantifying variation among garden plants in attractiveness to bees and other flower-visiting insects. Funct. Ecol. 2014, 28, 364–374. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M. Landscape effects on crop pollination services: Are there general patterns? Ecol. Let. 2008, 11, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.M.; Winfree, R. Local habitat characteristics but not landscape urbanization drive pollinator visitation and native plant pollination in forest remnants. Biol. Conserv. 2013, 160, 10–18. [Google Scholar] [CrossRef]
- Carper, A.L.; Adler, L.S.; Warren, P.S.; Irwin, R.E. Effects of suburbanization on forest bee communities. Environ. Entomol. 2014, 43, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Geslin, B.; Oddie, M.; Folschweiller, M.; Legras, G.; Seymour, C.L.; Van Veen, F.F.; Thébault, E. Spatiotemporal changes in flying insect abundance and their functional diversity as a function of distance to natural habitats in a mass flowering crop. Agric. Ecosyst. Environ. 2016, 229, 21–29. [Google Scholar] [CrossRef]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J.; Williams, N.M. Complex responses within a desert bee guild (Hymenoptera: Apiformes) to urban habitat fragmentation. Ecol. Appl. 2006, 16, 632–644. [Google Scholar] [CrossRef]
- Veddeler, D.; Klein, A.-M.; Tscharntke, T. Contrasting responses of bee communities to coffee flowering at different spatial scales. Oikos 2006, 112, 594–601. [Google Scholar] [CrossRef]
- Concepción, E.D.; Díaz, M.; Baquero, R.A. Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landsc. Ecol. 2008, 23, 135–148. [Google Scholar] [CrossRef]
- Woltz, J.M.; Isaacs, R.; Landis, D.A. Landscape structure and habitat management differentially influence insect natural enemies in an agricultural landscape. Agric. Ecosyst. Environ. 2012, 152, 40–49. [Google Scholar] [CrossRef]




| Family | Species | Urban Farms | Vacant Lots | Nesting Guild | Status | Size (mm) |
|---|---|---|---|---|---|---|
| Andrenidae | Andrena barbilabris | 0 | 1 | MINER | Native | 10.50 |
| Andrena commoda | 4 | 19 | MINER | Native | 11.00 | |
| Andrena cressonii | 1 | 3 | MINER | Native | 9.50 | |
| Andrena dunningi | 1 | 0 | MINER | Native | 11.50 | |
| Andrena erigeniae | 3 | 2 | MINER | Native | 7.50 | |
| Andrena miserabilis | 3 | 0 | MINER | Native | 7.60 | |
| Andrena nasonii | 5 | 0 | MINER | Native | 7.50 | |
| Andrena perplexa | 0 | 2 | MINER | Native | 12.00 | |
| Andrena violae | 0 | 9 | MINER | Native | 10.00 | |
| Andrena wilkella | 2 | 3 | MINER | Exotic | 10.50 | |
| Andrena ziziae | 0 | 1 | MINER | Native | 6.00 | |
| Calliopsis andreniformis | 4 | 15 | MINER | Native | 6.50 | |
| Apidae | Anthophora terminalis | 1 | 1 | MINER | Native | 11.75 |
| Apis mellifera | 40 | 35 | RENT | Exotic | 13.50 | |
| Bombus fervidus | 3 | 13 | RENT | Native | 10.50 | |
| Bombus griseocollis | 4 | 4 | RENT | Native | 13.75 | |
| Bombus impatiens | 15 | 2 | RENT | Native | 12.25 | |
| Bombus perplexus | 0 | 1 | RENT | Native | 13.00 | |
| Bombus vagans | 0 | 1 | RENT | Native | 10.75 | |
| Ceratina calcarata | 43 | 107 | PITH | Native | 6.50 | |
| Ceratina mikmaqi | 0 | 2 | PITH | Native | 6.50 | |
| Ceratina strenua | 0 | 1 | PITH | Native | 5.50 | |
| Holcopasites calliopsidis | 1 | 0 | NONEST | Native | 5.50 | |
| Melissodes bimaculatus | 70 | 49 | MINER | Native | 13.00 | |
| Melissodes druriellus | 0 | 1 | MINER | Native | 9.90 | |
| Melissodes trinodis | 1 | 2 | MINER | Native | 10.50 | |
| Nomada articulata | 1 | 2 | NONEST | Native | 9.00 | |
| Nomada bidentate group | 1 | 1 | NONEST | Native | 7.35 | |
| Nomada cressonii | 0 | 1 | NONEST | Native | 7.50 | |
| Nomada imbricata | 1 | 0 | NONEST | Native | 9.50 | |
| Nomada luteoloides | 1 | 2 | NONEST | Native | 7.00 | |
| Nomada maculata | 1 | 0 | NONEST | Native | 9.75 | |
| Nomada pygmaea | 0 | 1 | NONEST | Native | 7.25 | |
| Peponapis pruinosa | 25 | 34 | MINER | Native | 12.49 | |
| Triepeolus remigatus | 0 | 1 | NONEST | Native | 13.00 | |
| Xylocopa virginica | 3 | 5 | CARPENTER | Native | 20.00 | |
| Colletidae | Hylaeus affinis | 0 | 5 | RENT | Native | 5.50 |
| Hylaeus affinis/modestus | 0 | 5 | RENT | Native | 5.75 | |
| Hylaeus hyalinatus | 12 | 7 | RENT | Exotic | 5.89 | |
| Hylaeus modestus | 0 | 2 | RENT | Native | 5.75 | |
| Hylaeus pictipes | 0 | 3 | RENT | Exotic | 4.35 | |
| Halictidae | Agapostemon sericeus | 12 | 20 | MINER | Native | 9.41 |
| Agapostemon texanus | 39 | 61 | MINER | Native | 10.00 | |
| Agapostemon virescens | 279 | 436 | MINER | Native | 10.50 | |
| Augochlora pura | 1 | 6 | RENT | Native | 8.00 | |
| Augochlorella aurata | 10 | 22 | MINER | Native | 5.25 | |
| Halictus confusus | 38 | 82 | MINER | Native | 7.00 | |
| Halictus ligatus | 65 | 109 | MINER | Native | 8.50 | |
| Halictus rubicundus | 0 | 2 | MINER | Native | 10.50 | |
| Lasioglossum admirandum | 11 | 5 | MINER | Native | 5.25 | |
| Lasioglossum anomalum | 11 | 0 | MINER | Native | 4.00 | |
| Lasioglossum apocyni | 0 | 1 | MINER | Native | 4.56 | |
| Lasioglossum bruneri | 5 | 5 | MINER | Native | 6.30 | |
| Lasioglossum cattellae | 0 | 6 | MINER | Native | 5.10 | |
| Lasioglossum cinctipes | 0 | 1 | MINER | Native | 8.00 | |
| Lasioglossum coeruleum | 6 | 5 | MINER | Native | 6.18 | |
| Lasioglossum cressonii | 0 | 2 | MINER | Native | 5.96 | |
| Lasioglossum ellisiae | 3 | 3 | MINER | Native | 4.32 | |
| Lasioglossum ephialtum | 54 | 28 | MINER | Native | 5.31 | |
| Lasioglossum hitchensi | 23 | 40 | MINER | Native | 4.85 | |
| Lasioglossum illinoense | 5 | 4 | MINER | Native | 4.70 | |
| Lasioglossum imitatum | 39 | 45 | MINER | Native | 4.18 | |
| Lasioglossum laevissimum | 2 | 14 | MINER | Native | 6.11 | |
| Lasioglossum leucocomum | 19 | 55 | MINER | Native | 5.43 | |
| Lasioglossum leucozonium | 17 | 74 | MINER | Native | 7.50 | |
| Lasioglossum lineatulum | 2 | 6 | MINER | Native | 6.00 | |
| Lasioglossum lionotum | 0 | 1 | MINER | Native | 4.30 | |
| Lasioglossum pectorale | 24 | 26 | MINER | Native | 5.50 | |
| Lasioglossum pilosum | 65 | 53 | MINER | Native | 6.00 | |
| Lasioglossum subviridatum | 1 | 24 | MINER | Native | 10.79 | |
| Lasioglossum tegulare | 10 | 3 | MINER | Native | 8.30 | |
| Lasioglossum versatum | 1 | 2 | MINER | Native | 5.92 | |
| Lasioglossum weemsi | 8 | 8 | MINER | Native | 9.14 | |
| Lasioglossum zephyrum | 4 | 2 | MINER | Native | 6.08 | |
| Lasioglossum zonulum | 0 | 2 | MINER | Native | 7.50 | |
| Sphecodes atlantis | 0 | 1 | NONEST | Native | 5.63 | |
| Sphecodes coronus | 1 | 1 | NONEST | Native | 5.85 | |
| Sphecodes cressonii | 2 | 1 | NONEST | Native | 5.25 | |
| Sphecodes mandibularis | 3 | 0 | NONEST | Native | 5.36 | |
| Sphecodes minor | 0 | 1 | NONEST | Native | 6.75 | |
| Megachilidae | Anthidium manicatum | 1 | 1 | MINER | Exotic | 14.00 |
| Anthidium oblongatum | 1 | 22 | MINER | Exotic | 8.55 | |
| Megachile apicalis | 2 | 0 | RENT | Exotic | 7.50 | |
| Megachile brevis | 1 | 0 | RENT | Native | 9.50 | |
| Megachile centuncularis | 1 | 3 | RENT | Native | 9.50 | |
| Megachile frigida | 1 | 0 | RENT | Native | 13.00 | |
| Megachile mendica | 2 | 3 | RENT | Native | 10.50 | |
| Megachile pugnata | 0 | 1 | RENT | Native | 14.50 | |
| Megachile relativa | 0 | 1 | RENT | Native | 10.00 | |
| Megachile rotundata | 12 | 28 | RENT | Exotic | 8.00 | |
| Megachile texana | 1 | 2 | RENT | Native | 12.00 | |
| Osmia atriventris | 0 | 1 | RENT | Native | 7.50 | |
| Osmia bucephala | 1 | 1 | RENT | Native | 14.50 | |
| Osmia caerulescens | 4 | 33 | RENT | Exotic | 10.00 | |
| Osmia georgica | 9 | 13 | RENT | Native | 8.25 | |
| Osmia pumila | 3 | 14 | RENT | Native | 7.50 | |
| Osmia subfasciata | 1 | 0 | RENT | Native | 7.50 | |
| Osmia taurus | 2 | 0 | RENT | Exotic | 11.10 | |
| TOTAL | 1048 | 1623 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivakoff, F.S.; Prajzner, S.P.; Gardiner, M.M. Unique Bee Communities within Vacant Lots and Urban Farms Result from Variation in Surrounding Urbanization Intensity. Sustainability 2018, 10, 1926. https://doi.org/10.3390/su10061926
Sivakoff FS, Prajzner SP, Gardiner MM. Unique Bee Communities within Vacant Lots and Urban Farms Result from Variation in Surrounding Urbanization Intensity. Sustainability. 2018; 10(6):1926. https://doi.org/10.3390/su10061926
Chicago/Turabian StyleSivakoff, Frances S., Scott P. Prajzner, and Mary M. Gardiner. 2018. "Unique Bee Communities within Vacant Lots and Urban Farms Result from Variation in Surrounding Urbanization Intensity" Sustainability 10, no. 6: 1926. https://doi.org/10.3390/su10061926
APA StyleSivakoff, F. S., Prajzner, S. P., & Gardiner, M. M. (2018). Unique Bee Communities within Vacant Lots and Urban Farms Result from Variation in Surrounding Urbanization Intensity. Sustainability, 10(6), 1926. https://doi.org/10.3390/su10061926