Physical Forcing-Driven Productivity and Sediment Flux to the Deep Basin of Northern South China Sea: A Decadal Time Series Study

 ,
,  ,
, 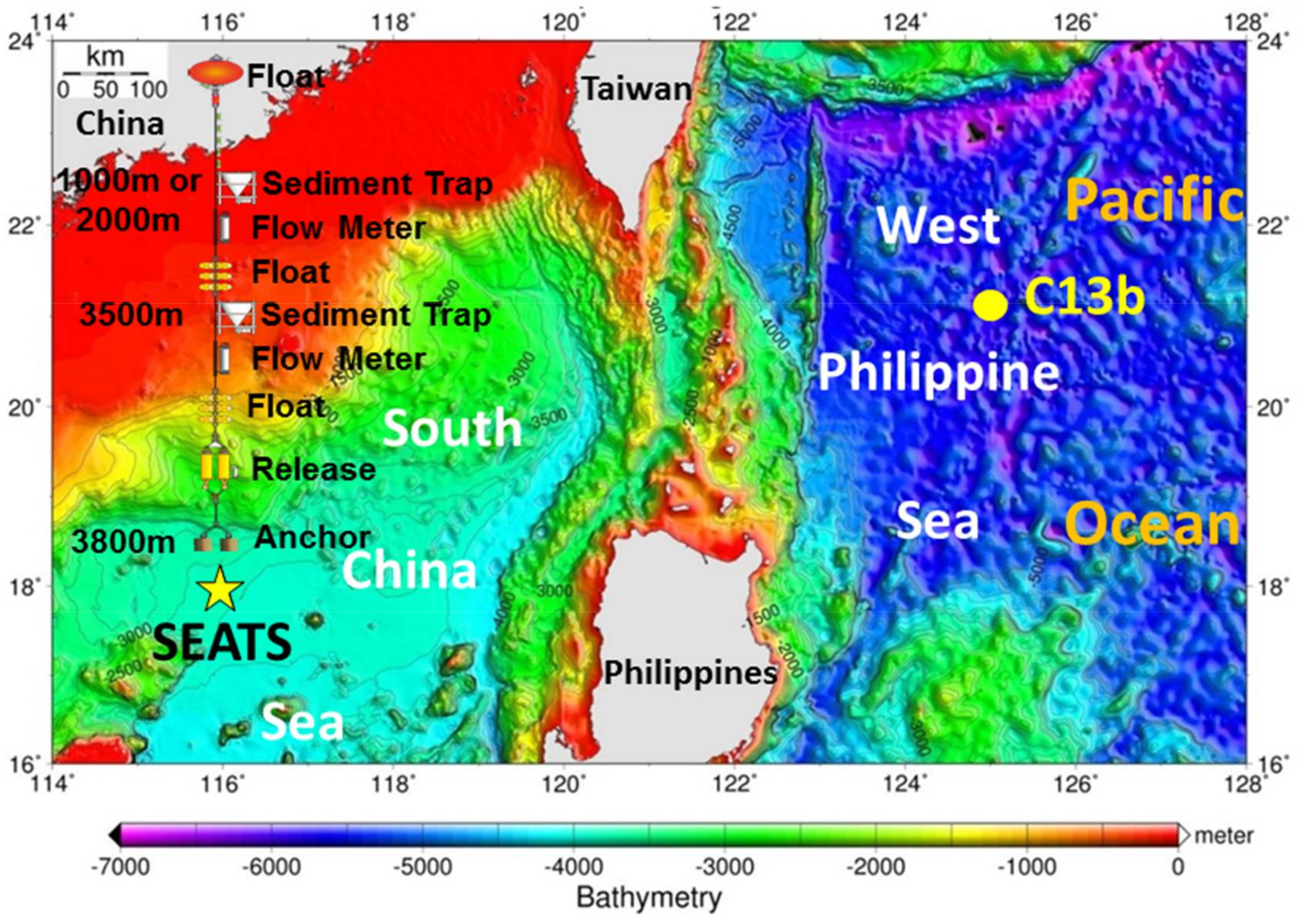{kind=link}
{kind=link}
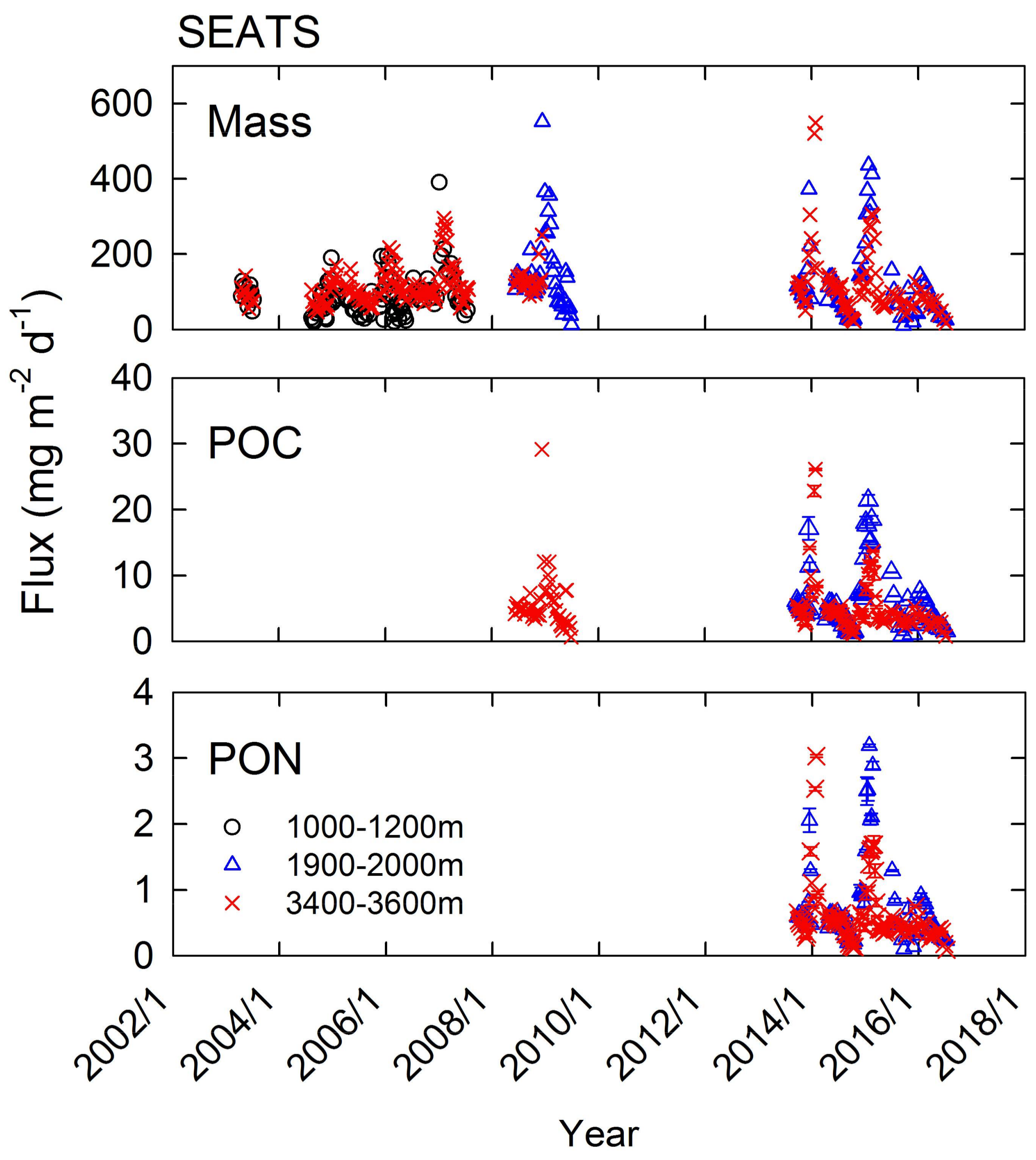{kind=link}
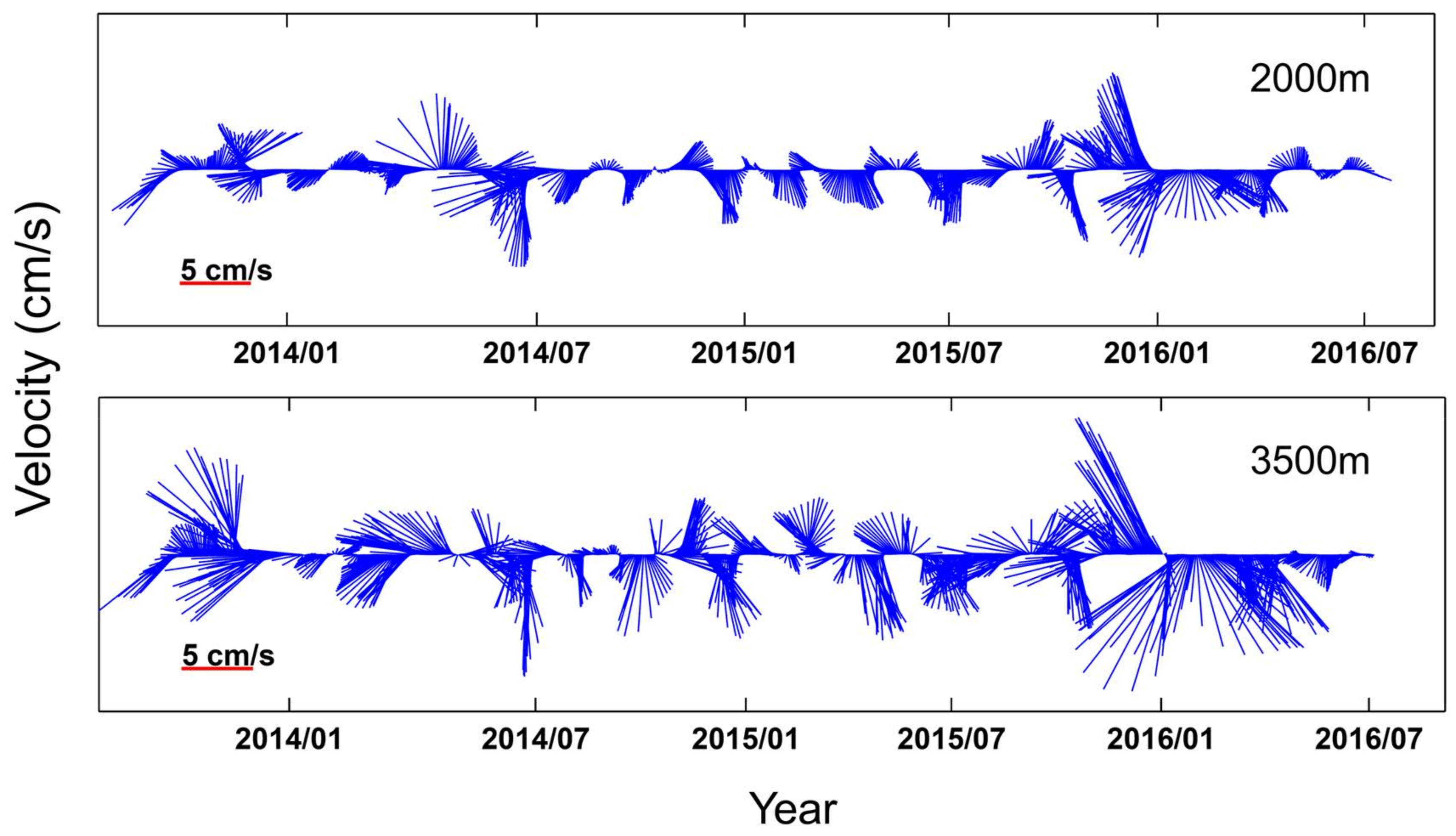{kind=link}
{kind=link}
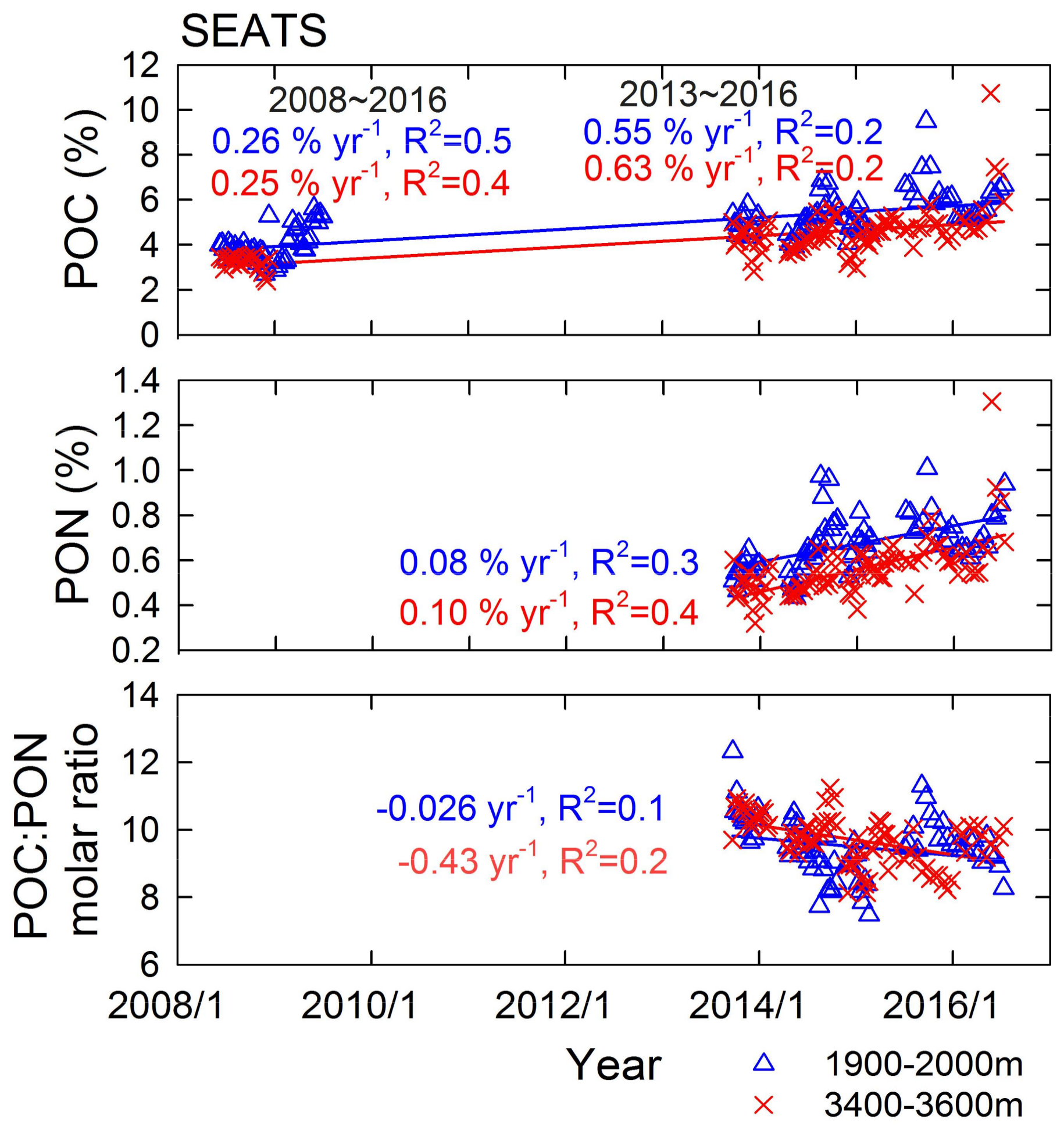{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Consistences in Seasonal and Interannual Variations between Mass Flux and chlorophyll-a
3.2. POC and PON Dynamics
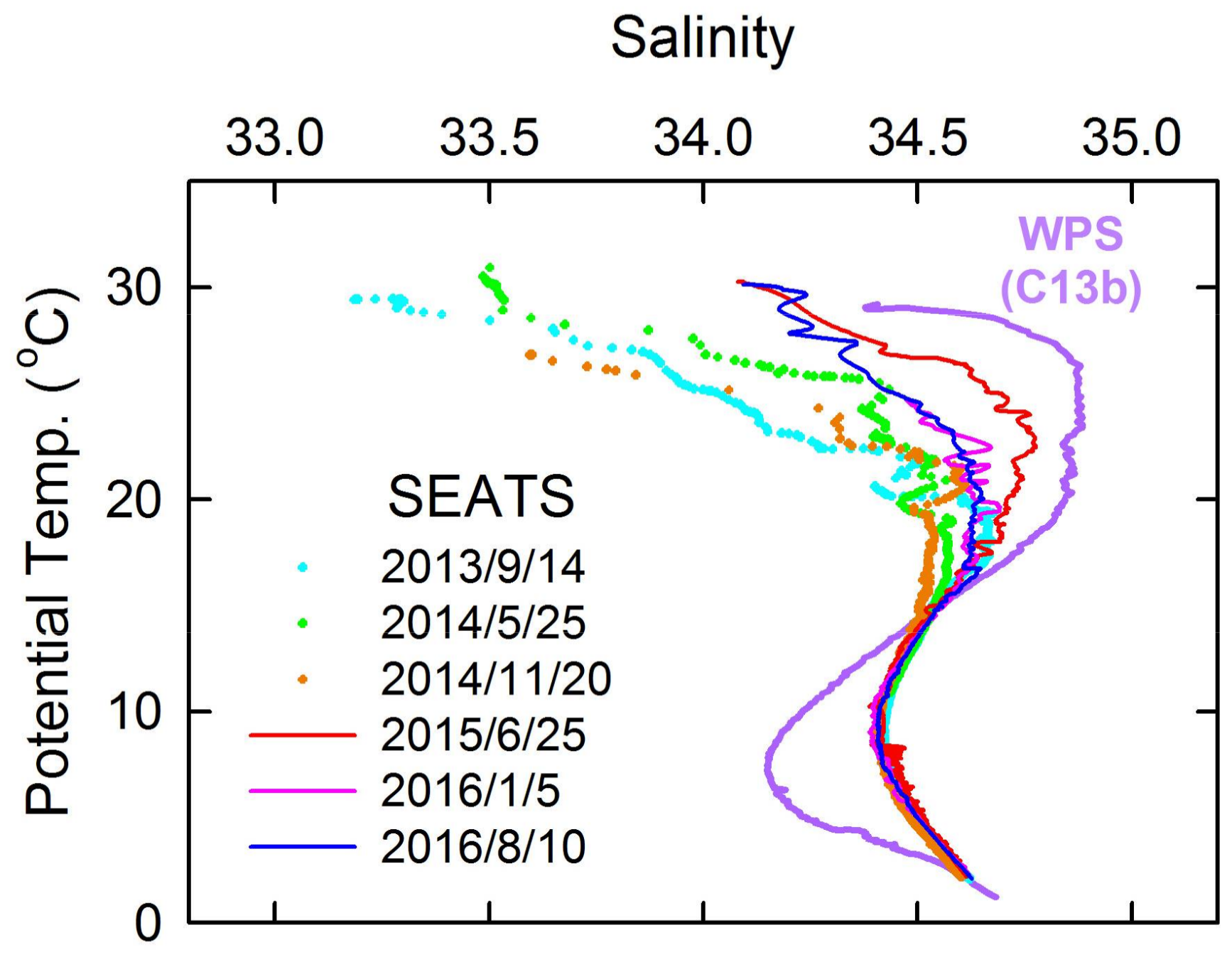
3.3. Intrusion of WPS Seawater Reduces Export Production and Mass Flux
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Le Quere, C.; Takahashi, T.; Buitenhuis, E.T.; Rodenbeck, C.; Sutherland, S.C. Impact of climate change and variability on the global oceanic sink of CO2. Glob. Biogeochem. Cycles 2010, 24. [Google Scholar] [CrossRef]
- Sabine, C.L.; Feely, R.A.; Gruber, N.; Key, R.M.; Lee, K.; Bullister, J.L.; Wanninkhof, R.; Wong, C.S.; Wallace, D.W.R.; Tilbrook, B.; et al. The oceanic sink for anthropogenic CO2. Science 2004, 305, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Sutherland, S.C.; Wanninkhof, R.; Sweeney, C.; Feely, R.A.; Chipman, D.W.; Hales, B.; Friederich, G.; Chavez, F.; Sabine, C.; et al. Climatological mean and decadal change in surface ocean pCO2, and net sea–air CO2 flux over the global oceans. Deep-Sea Res. Part II 2009, 56, 554–577. [Google Scholar] [CrossRef]
- Sarmiento, J.L.; Gruber, N. Sinks for Anthropogenic Carbon. Phys. Today 2002, 55, 30–36. [Google Scholar] [CrossRef]
- Wong, G.T.F.; Ku, T.L.; Mulholland, M.; Tseng, C.M.; Wang, D.P. The SouthEast Asian Time-series Study (SEATS) and the biogeochemistry of the South China Sea—An overview. Deep-Sea Res. Part II 2007, 54, 1434–1447. [Google Scholar] [CrossRef]
- O’Reilly, J.E.; Maritorena, S.; Mitchell, B.G.; Siegel, D.A.; Carder, K.L.; Garver, S.A.; Kahru, M.; McClain, C. Ocean color chlorophyll algorithms for SeaWiFS. J. Geophys. Res. 1998, 103, 24937–24953. [Google Scholar] [CrossRef]
- Hu, C.; Lee, Z.; Franz, B. Chlorophyllaalgorithms for oligotrophic oceans: A novel approach based on three-band reflectance difference. J. Geophys. Res. 2012, 117. [Google Scholar] [CrossRef]
- Abell, R.E.; Brand, T.; Dale, A.C.; Tilstone, G.H.; Beveridge, C. Variability of particulate flux over the Mid-Atlantic Ridge. Deep-Sea Res. Part II 2013, 98, 257–268. [Google Scholar] [CrossRef][Green Version]
- Wei, C.L.; Chia, C.Y.; Chou, W.C.; Lee, W.H. Sinking fluxes of 210Pb and 210Po in the deep basin of the northern South China Sea. J. Environ. Radioact. 2017, 174, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.M. A unique seasonal pattern in phytoplankton biomass in low-latitude waters in the South China Sea. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Liu, K.K.; Wang, L.W.; Dai, M.; Tseng, C.M.; Yang, Y.; Sui, C.H.; Oey, L.; Tseng, K.Y.; Huang, S.M. Inter-annual variation of chlorophyll in the northern South China Sea observed at the SEATS Station and its asymmetric responses to climate oscillation. Biogeosciences 2013, 10, 7449–7462. [Google Scholar] [CrossRef]
- Cai, P.; Zhao, D.; Wang, L.; Huang, B.; Dai, M. Role of particle stock and phytoplankton community structure in regulating particulate organic carbon export in a large marginal sea. J. Geophys. Res. 2015, 120, 2063–2095. [Google Scholar] [CrossRef]
- Martin, P.; Lampitt, R.S.; Jane Perry, M.; Sanders, R.; Lee, C.; D'Asaro, E. Export and mesopelagic particle flux during a North Atlantic spring diatom bloom. Deep-Sea Res. Part. I 2011, 58, 338–349. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Zhao, Y.; Wang, W.; Li, J.; Xu, J. Mesoscale eddies transport deep-sea sediments. Sci. Rep. 2014, 4, 5937. [Google Scholar] [CrossRef] [PubMed]
- Schneider, B.; Schlitzer, R.; Fischer, G.; Nöthig, E.-M. Depth-dependent elemental compositions of particulate organic matter (POM) in the ocean. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Martiny, A.C.; Vrugt, J.A.; Lomas, M.W. Concentrations and ratios of particulate organic carbon, nitrogen, and phosphorus in the global ocean. Sci. Data 2014, 1, 140048. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; He, X.; Yu, S.; Chen, C.T.A. Changes in the Ecological Environment of the Marginal Seas along the Eurasian Continent from 2003 to 2014. Sustainability 2018, 10. [Google Scholar] [CrossRef]
- Martin, J.H.; Knauer, G.A.; Karl, D.M.; Broenkow, W.W. VERTEX: Carbon cycling in the northeast Pacific. Deep-Sea Res. Part A 1987, 34, 267–285. [Google Scholar] [CrossRef]
- Yang, J.-Y.T.; Kao, S.-J.; Dai, M.; Yan, X.; Lin, H.-L. Examining N cycling in the northern South China Sea from N isotopic signals in nitrate and particulate phases. J. Geophys. Res. 2017, 122, 2118–2136. [Google Scholar] [CrossRef]
- Liu, J.; Clift, P.D.; Yan, W.; Chen, Z.; Chen, H.; Xiang, R.; Wang, D. Modern transport and deposition of settling particles in the northern South China Sea: Sediment trap evidence adjacent to Xisha Trough. Deep-Sea Res. Part I 2014, 93, 145–155. [Google Scholar] [CrossRef]
- Ho, T.Y.; Chou, W.C.; Lin, H.L.; Sheu, D.D. Trace metal cycling in the deep water of the South China Sea: The composition, sources, and fluxes of sinking particles. Limnol. Oceanogr. 2011, 56, 1225–1243. [Google Scholar] [CrossRef]
- Lin, H.L.; Sheu, D.D.D.; Yang, Y.; Chou, W.C.; Hung, G.W. Stable isotopes in modern planktonic foraminifera: Sediment trap and plankton tow results from the South China Sea. Mar. Micropaleontol. 2011, 79, 15–23. [Google Scholar] [CrossRef]
- Tseng, C.M.; Gong, G.C.; Wang, L.W.; Liu, K.K.; Yang, Y. Anomalous biogeochemical conditions in the northern South China Sea during the El-Niño events between 1997 and 2003. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef]
- Tseng, C.M.; Wong, G.T.F.; Chou, W.C.; Lee, B.S.; Sheu, D.D.; Liu, K.K. Temporal variations in the carbonate system in the upper layer at the SEATS station. Deep-Sea Res. Part II 2007, 54, 1448–1468. [Google Scholar] [CrossRef]
- Tseng, C.M.; Liu, K.K.; Wang, L.W.; Gong, G.C. Anomalous hydrographic and biological conditions in the northern South China Sea during the 1997–1998 El Niño and comparisons with the equatorial Pacific. Deep-Sea Res. Part I 2009, 56, 2129–2143. [Google Scholar] [CrossRef]
- Chen, C.T.A.; Wang, S.L.; Wang, B.J.; Pai, S.C. Nutrient budgets for the South China Sea basin. Mar. Chem. 2001, 75, 281–300. [Google Scholar] [CrossRef]
- Wu, C.R.; Hsin, Y.C. The forcing mechanism leading to the Kuroshio intrusion into the South China Sea. J. Geophys. Res. 2012, 117. [Google Scholar] [CrossRef]
- Wu, C.R.; Wang, Y.L.; Lin, Y.F.; Chiang, T.L.; Wu, C.C. Weakening of the Kuroshio Intrusion Into the South China Sea under the Global Warming Hiatus. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2016, 9, 5064–5070. [Google Scholar] [CrossRef]
- Huang, T.H.; Chen, C.T.A.; Zhang, W.Z.; Zhuang, X.F. Varying intensity of Kuroshio intrusion into Southeast Taiwan Strait during ENSO events. Cont. Shelf Res. 2015, 103, 79–87. [Google Scholar] [CrossRef]
- Chen, C.T.A.; Yeh, Y.T.; Chen, Y.C.; Huang, T.H. Seasonal and ENSO-related interannual variability of subsurface fronts separating West Philippine Sea waters from South China Sea waters near the Luzon Strait. Deep-Sea Res. Part I 2015, 103, 13–23. [Google Scholar] [CrossRef]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lui, H.-K.; Chen, K.-Y.; Chen, C.-T.A.; Wang, B.-S.; Lin, H.-L.; Ho, S.-H.; Tseng, C.-J.; Yang, Y.; Chan, J.-W. Physical Forcing-Driven Productivity and Sediment Flux to the Deep Basin of Northern South China Sea: A Decadal Time Series Study. Sustainability 2018, 10, 971. https://doi.org/10.3390/su10040971
Lui H-K, Chen K-Y, Chen C-TA, Wang B-S, Lin H-L, Ho S-H, Tseng C-J, Yang Y, Chan J-W. Physical Forcing-Driven Productivity and Sediment Flux to the Deep Basin of Northern South China Sea: A Decadal Time Series Study. Sustainability. 2018; 10(4):971. https://doi.org/10.3390/su10040971
Chicago/Turabian StyleLui, Hon-Kit, Kuang-Yu Chen, Chen-Tung Arthur Chen, Bo-Shian Wang, Hui-Ling Lin, Shih-Hu Ho, Chiung-Jung Tseng, Yih Yang, and Jui-Wen Chan. 2018. "Physical Forcing-Driven Productivity and Sediment Flux to the Deep Basin of Northern South China Sea: A Decadal Time Series Study" Sustainability 10, no. 4: 971. https://doi.org/10.3390/su10040971
APA StyleLui, H.-K., Chen, K.-Y., Chen, C.-T. A., Wang, B.-S., Lin, H.-L., Ho, S.-H., Tseng, C.-J., Yang, Y., & Chan, J.-W. (2018). Physical Forcing-Driven Productivity and Sediment Flux to the Deep Basin of Northern South China Sea: A Decadal Time Series Study. Sustainability, 10(4), 971. https://doi.org/10.3390/su10040971