

“Every Cloud Has a Silver Lining”: How Three Rare Diseases Defend Themselves from COVID-19 and What We Have Learnt from It

 , ,
, ,  and
and 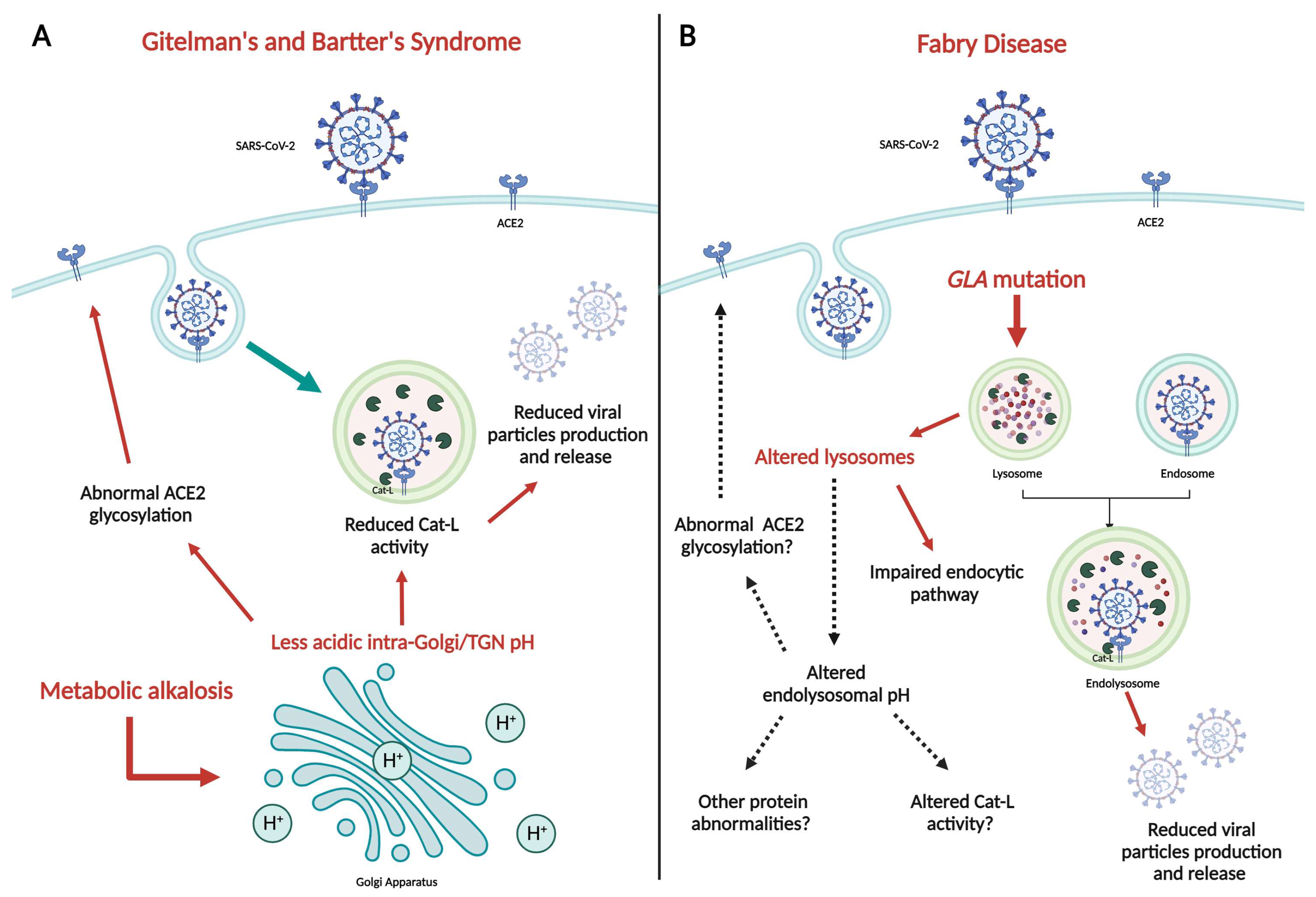{kind=link}
Abstract
1. Introduction
2. Mas Receptor and AT1R: The Good and the Bad
3. Gitelman and Bartter Syndromes: Two “In Vivo” Models of SARS-CoV-2 Innate Protection
4. Fabry Disease and SARS-CoV-2
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Senapati, S.; Banerjee, P.; Bhagavatula, S.; Kushwaha, P.P.; Kumar, S. Contributions of human ACE2 and TMPRSS2 in determining host–pathogen interaction of COVID-19. J. Genet. 2021, 100, 12. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Joyce, R.; Tan, H.; Hu, Y.; Wang, J. SARS-CoV-2 Main Protease Drug Design, Assay Development, and Drug Resistance Studies. Acc. Chem. Res. 2023, 56, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Chen, X.; Geiger, J.D. Role of Endolysosomes in Severe Acute Respiratory Syndrome Coronavirus-2 Infection and Coronavirus Disease 2019 Pathogenesis: Implications for Potential Treatments. Front. Pharmacol. 2020, 11, 595888. [Google Scholar] [CrossRef] [PubMed]
- Isobe, A.; Arai, Y.; Kuroda, D.; Okumura, N.; Ono, T.; Ushiba, S.; Nakakita, S.-I.; Daidoji, T.; Suzuki, Y.; Nakaya, T.; et al. ACE2 N-glycosylation modulates interactions with SARS-CoV-2 spike protein in a site-specific manner. Commun. Biol. 2022, 5, 1188. [Google Scholar] [CrossRef] [PubMed]
- Caputo, I.; Bertoldi, G.; Driussi, G.; Stefanelli, L.F.; Di Vico, V.; Carraro, G.; Nalesso, F.; Calò, L.A. The RAAS Goodfellas in Cardiovascular System. J. Clin. Med. 2023, 12, 6873. [Google Scholar] [CrossRef]
- Angeli, F.; Zappa, M.; Reboldi, G.; Trapasso, M.; Cavallini, C.; Spanevello, A.; Verdecchia, P. The pivotal link between ACE2 deficiency and SARS-CoV-2 infection: One year later. Eur. J. Intern. Med. 2021, 93, 28–34. [Google Scholar] [CrossRef]
- Calò, L.A.; Stefanelli, L.F.; Nalesso, F. Angiotensin II, RAS Activation, and RAS Blockers in COVID-19: Unambiguous Evidence. Kidney Blood Press. Res. 2022, 47, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Gnanenthiran, S.R.; Borghi, C.; Burger, D.; Caramelli, B.; Charchar, F.; Chirinoset, J.A.; Cohen, J.B.; Cremer, A.; Di Tanna, G.L.; Duvignaud, A.; et al. Renin-Angiotensin System Inhibitors in Patients with COVID-19: A Meta-Analysis of Randomized Controlled Trials Led by the International Society of Hypertension. J. Am. Heart Assoc. 2022, 11, e026143. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, A.; Bockenhauer, D.; Bolignano, D.; Calò, L.A.; Cosyns, E.; Devuyst, O.; Ellison, D.H.; Frankl, F.E.K.; Knoers, N.V.; Konrad, M.; et al. Gitelman syndrome: Consensus and guidance from a kidney disease: Improving global outcomes (KDIGO) controversies conference. Kidney Int. 2017, 91, 24–33. [Google Scholar] [CrossRef]
- Konrad, M.; Nijenhuis, T.; Ariceta, G.; Bertholet-Thomas, A.; Calo, L.A.; Capasso, G.; Emma, F.; Schlingmann, K.P.; Singh, M.; Trepiccione, F.; et al. Diagnosis and management of Bartter syndrome: Executive summary of the consensus and recommendations from the European Rare Kidney Disease Reference Network Working Group for tubular disorders. Kidney Int. 2021, 99, 324–335. [Google Scholar] [CrossRef]
- Calò, L.A.; Davis, P.A.; Rossi, G.P. Understanding the mechanisms of angiotensin II signaling involved in hypertension and its long-term sequelae. J. Hypertens. 2014, 32, 2109–2119. [Google Scholar] [CrossRef] [PubMed]
- Calò, L.A.; Rigato, M.; Sgarabotto, L.; Gianesello, L.; Bertoldi, G.; Ravarotto, V.; Davis, P.A. ACE2 and SARS-CoV-2 Infection Risk: Insights from Patients with Two Rare Genetic Tubulopathies, Gitelman’s and Bartter’s Syndromes. Front. Med. 2021, 8, 647319. [Google Scholar] [CrossRef] [PubMed]
- Bertoldi, G.; Ravarotto, V.; Sgarabotto, L.; Davis, P.A.; Gobbi, L.; Calò, L.A. Impaired ACE2 glycosylation and protease activity lowers COVID-19 susceptibility in Gitelman’s and Bartter’s syndromes. J. Intern. Med. 2022, 291, 522–524. [Google Scholar] [CrossRef] [PubMed]
- Vincent, M.J.; Bergeron, E.; Benjannet, S.; Erickson, B.R.; Rollin, P.E.; Ksiazek, T.G.; Seidah, N.G.; Nichol, S.T. Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol. J. 2005, 2, 69. [Google Scholar] [CrossRef]
- Abedi, F.; Hayes, A.W.; Reiter, R.; Karimi, G. Acute lung injury: The therapeutic role of Rho kinase inhibitors. Pharmacol. Res. 2020, 155, 104736. [Google Scholar] [CrossRef] [PubMed]
- Calò, L.A.; Bertoldi, G.; Davis, P.A. Rho kinase inhibitors for SARS-CoV-2 induced acute respiratory distress syndrome: Support from Bartter’s and Gitelman’s syndrome patients. Pharmacol. Res. 2020, 158, 104903. [Google Scholar] [CrossRef] [PubMed]
- Kok, K.; Zwiers, K.C.; Boot, R.G.; Overkleeft, H.S.; Aerts, J.M.F.G.; Artola, M. Fabry Disease: Molecular Basis, Pathophysiology, Diagnostics and Potential Therapeutic Directions. Biomolecules 2021, 11, 271. [Google Scholar] [CrossRef] [PubMed]
- Bertoldi, G.; Caputo, I.; Driussi, G.; Stefanelli, L.F.; Di Vico, V.; Carraro, G.; Nalesso, F.; Calò, L.A. Biochemical Mechanisms beyond Glycosphingolipid Accumulation in Fabry Disease: Might They Provide Additional Therapeutic Treatments? J. Clin. Med. 2023, 12, 2063. [Google Scholar] [CrossRef] [PubMed]
- Pieroni, M.; Pieruzzi, F.; Mignani, R.; Graziani, F.; Olivotto, I.; Riccio, E.; Ciabatti, M.; Limongelli, G.; Manna, R.; Bolognese, L.; et al. Potential resistance to SARS-CoV-2 infection in lysosomal storage disorders. Clin. Kidney J. 2021, 14, 1488–1490. [Google Scholar] [CrossRef]
- Caputo, I.; Bertoldi, G.; Driussi, G.; Sgarabotto, L.; Carraro, G.; Stefanelli, L.F.; Davis, P.A.; Calò, L.A. Impaired ACE2 glycosylation and protease activity in Fabry disease protects from COVID-19. J. Intern. Med. 2023, 294, 238–240. [Google Scholar] [CrossRef]
- Resnick, S.J.; Iketani, S.; Hong, S.J.; Zask, A.; Liu, H.; Kim, S.; Melore, S.; Lin, F.-Y.; Nair, M.S.; Huang, Y.; et al. Inhibitors of coronavirus 3CL proteases protect cells from protease-mediated cytotoxicity. J. Virol. 2021, 95, e0237420. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cacciapuoti, M.; Caputo, I.; Stefanelli, L.F.; Davis, P.A.; Nalesso, F.; Calò, L.A. “Every Cloud Has a Silver Lining”: How Three Rare Diseases Defend Themselves from COVID-19 and What We Have Learnt from It. Clin. Pract. 2024, 14, 614-618. https://doi.org/10.3390/clinpract14020048
Cacciapuoti M, Caputo I, Stefanelli LF, Davis PA, Nalesso F, Calò LA. “Every Cloud Has a Silver Lining”: How Three Rare Diseases Defend Themselves from COVID-19 and What We Have Learnt from It. Clinics and Practice. 2024; 14(2):614-618. https://doi.org/10.3390/clinpract14020048
Chicago/Turabian StyleCacciapuoti, Martina, Ilaria Caputo, Lucia Federica Stefanelli, Paul A. Davis, Federico Nalesso, and Lorenzo A. Calò. 2024. "“Every Cloud Has a Silver Lining”: How Three Rare Diseases Defend Themselves from COVID-19 and What We Have Learnt from It" Clinics and Practice 14, no. 2: 614-618. https://doi.org/10.3390/clinpract14020048
APA StyleCacciapuoti, M., Caputo, I., Stefanelli, L. F., Davis, P. A., Nalesso, F., & Calò, L. A. (2024). “Every Cloud Has a Silver Lining”: How Three Rare Diseases Defend Themselves from COVID-19 and What We Have Learnt from It. Clinics and Practice, 14(2), 614-618. https://doi.org/10.3390/clinpract14020048