
Population Dynamics, Plasma Cytokines and Platelet Centrifugation: Technical and Sociodemographic Aspects of ‘Ovarian Rejuvenation’



{kind=link}
Abstract
1. Introduction
2. Boundaries and Beginnings
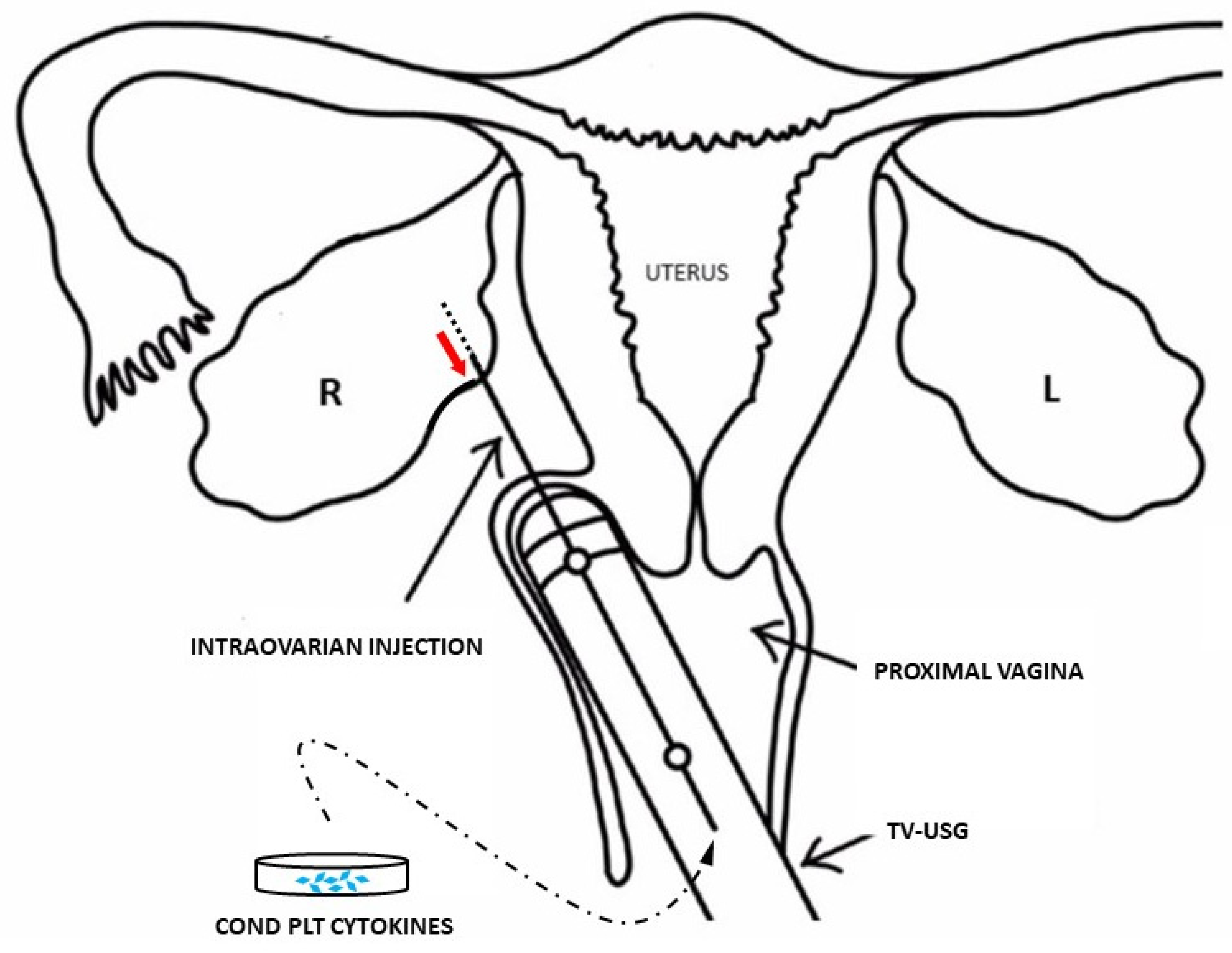
3. Centrifugation for Platelets and Their Stem-Cell Targets
4. Activation of Fresh Platelets
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tiniakos, D.G.; Kandilis, A.; Geller, S.A. Tityus: A forgotten myth of liver regeneration. J. Hepatol. 2010, 53, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Pantos, K.; Nitsos, N.; Kokkali, G.; Vaxevanoglou, T.; Markomichali, C.; Pantou, A.; Grammatis, M.; Lazaros, L.; Sfakianoudis, K. Ovarian Rejuvenation and Folliculogenesis Reactivation in Perimenopausal Women after Autologous Platelet-Rich Plasma Treatment [abstract]. ESHRE 32nd Annual Meeting (Helsinki). Hum. Reprod. 2016, i301. [Google Scholar]
- Storer, M.A.; Miller, F.D. A finger on the pulse of regeneration: Insights into cellular mechanisms of adult digit tip regeneration. Curr. Opin. Genet. Dev. 2021, 70, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Wang, C.; Zhou, Z.; Zhang, M.; Zhao, D.; Bai, X.; Leng, X. Comparative transcriptome analysis of the main beam and brow tine of sika deer antler provides insights into the molecular control of rapid antler growth. Cell. Mol. Biol. Lett. 2020, 25, 42. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Ohgo, S.; Yokoyama, H. Limb blastema cell: A stem cell for morphological regeneration. Dev. Growth Differ. 2010, 52, 89–99. [Google Scholar] [CrossRef]
- Hyun, J.S.; Chung, M.T.; Wong, V.W.; Montoro, D.; Longaker, M.T.; Wan, D.C. Rethinking the blastema. Plast. Reconstr. Surg. 2012, 129, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Hummitzsch, K.; Anderson, R.A.; Wilhelm, D.; Wu, J.; Telfer, E.E.; Russell, D.L.; Robertson, S.A.; Rodgers, R.J. Stem cells, progenitor cells, and lineage decisions in the ovary. Endocr. Rev. 2015, 36, 65–91. [Google Scholar] [CrossRef]
- Martin, J.J.; Woods, D.C.; Tilly, J.L. Implications and current limitations of oogenesis from female germline or oogonial stem cells in adult mammalian ovaries. Cells 2019, 8, 93. [Google Scholar] [CrossRef]
- Telfer, E.E.; Anderson, R.A. The existence and potential of germline stem cells in the adult mammalian ovary. Climacteric 2019, 22, 22–26. [Google Scholar] [CrossRef]
- Sfakianoudis, K.; Simopoulou, M.; Grigoriadis, S.; Pantou, A.; Tsioulou, P.; Maziotis, E.; Rapani, A.; Giannelou, P.; Nitsos, N.; Kokkali, G.; et al. Reactivating ovarian function through autologous platelet-rich plasma intraovarian infusion: Pilot data on premature ovarian insufficiency, perimenopausal, menopausal, and poor responder women. J. Clin. Med. 2020, 9, 1809. [Google Scholar] [CrossRef]
- Sills, E.S. The scientific and cultural journey to ovarian rejuvenation: Background, barriers, and beyond the biological clock. Medicines 2021, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Sun, J.; Zhang, L.; Chen, S.; Dong, W.; Yu, H.; Han, B.; Tan, M.; Li, X. Comparative effectiveness of minimally invasive nonsurgical treatments for plantar fasciitis: A network meta-analysis of 30 randomized controlled trials. Pain Physician 2021, 24, E955–E971. [Google Scholar] [PubMed]
- Wang, S.L.; Liu, X.L.; Kang, Z.C.; Wang, Y.S. Platelet-rich plasma promotes peripheral nerve regeneration after sciatic nerve injury. Neural Regen. Res. 2023, 18, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Sills, E.S.; Wood, S.H. Growth factors, gene activation, and cell recruitment: From intraovarian condensed platelet cytokines to de novo oocyte development. J. Clin. Transl. Res. 2022, 8, 49–53. [Google Scholar] [PubMed]
- Virant-Klun, I.; Skutella, T.; Hren, M.; Gruden, K.; Cvjeticanin, B.; Vogler, A.; Sinkovec, J. Isolation of small SSEA-4-positive putative stem cells from the ovarian surface epithelium of adult human ovaries by two different methods. Biomed. Res. Int. 2013, 2013, 690415. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Zuba-Surma, E.; Wojakowski, W.; Suszynska, M.; Mierzejewska, K.; Liu, R.; Ratajczak, J.; Shin, D.M.; Kucia, M. Very small embryonic-like stem cells (VSELs) represent a real challenge in stem cell biology: Recent pros and cons in the midst of a lively debate. Leukemia 2014, 28, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Ratajczak, J.; Kucia, M. Very small embryonic-like stem cells (VSELs). Circ. Res. 2019, 124, 208–210. [Google Scholar] [CrossRef]
- Patel, H.; Bhartiya, D.; Parte, S.; Gunjal, P.; Yedurkar, S.; Bhatt, M. Follicle stimulating hormone modulates ovarian stem cells through alternately spliced receptor variant FSH-R3. J. Ovarian Res. 2013, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Sills, E.S.; Wood, S.H. Autologous activated platelet-rich plasma injection into adult human ovary tissue: Molecular mechanism, analysis, and discussion of reproductive response. Biosci. Rep. 2019, 39, BSR20190805. [Google Scholar] [CrossRef]
- Shaikh, A.; Anand, S.; Kapoor, S.; Ganguly, R.; Bhartiya, D. Mouse bone marrow VSELs exhibit differentiation into three embryonic germ lineages and germ & hematopoietic cells in culture. Stem Cell Rev. Rep. 2017, 13, 202–216. [Google Scholar] [CrossRef]
- Yang, S.; Liu, Z.; Wu, S.; Zou, L.; Cao, Y.; Xu, H.; Huang, J.; Tian, Q.; Wu, F.; Li, P.; et al. Meiosis resumption in human primordial germ cells from induced pluripotent stem cells by in vitro activation and reconstruction of ovarian nests. Stem Cell Res. Ther. 2022, 13, 339. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Feng, F.; Shi, F.; Huang, J.; Zhang, S.; Quan, Y.; Tu, T.; Liu, Y.; Wang, J.; Zhao, Y.; et al. Distinctive roles of Wnt signaling in chondrogenic differentiation of BMSCs under coupling of pressure and platelet-rich fibrin. Tissue Eng. Regen Med. 2022, 19, 823–837. [Google Scholar] [CrossRef]
- Qi, Y.; Tang, R.; Shi, Z.; Feng, G.; Zhang, W. Wnt5a/Platelet-rich plasma synergistically inhibits IL-1β-induced inflammatory activity through NF-κB signaling pathway and prevents cartilage damage and promotes meniscus regeneration. J. Tissue Eng. Regen Med. 2021, 15, 612–624. [Google Scholar] [CrossRef] [PubMed]
- Castrillon, D.H.; Quade, B.J.; Wang, T.Y.; Quigley, C.; Crum, C.P. The human VASA gene is specifically expressed in the germ cell lineage. Proc. Natl. Acad. Sci. USA 2000, 97, 9585–9590. [Google Scholar] [CrossRef]
- Gkountela, S.; Li, Z.; Vincent, J.J.; Zhang, K.X.; Chen, A.; Pellegrini, M.; Clark, A.T. The ontogeny of cKIT+ human primordial germ cells proves to be a resource for human germ line reprogramming, imprint erasure and in vitro differentiation. Nat. Cell Biol. 2013, 15, 113–122. [Google Scholar] [CrossRef]
- Gomes Fernandes, M.; Bialecka, M.; Salvatori, D.C.F.; Chuva de Sousa Lopes, S.M. Characterization of migratory primordial germ cells in the aorta-gonad-mesonephros of a 4.5-week-old human embryo: A toolbox to evaluate in vitro early gametogenesis. Mol. Hum. Reprod. 2018, 24, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Chitiashvili, T.; Hsu, F.M.; Dror, I.; Plath, K.; Clark, A. FGFR3 is expressed by human primordial germ cells and is repressed after meiotic initiation to form primordial oocytes. Stem Cell Rep. 2022, 17, 1268–1278. [Google Scholar] [CrossRef]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.K.; Tilly, J.L. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 428, 145–150. [Google Scholar] [CrossRef]
- Raz, E. The function and regulation of vasa-like genes in germ-cell development. Genome Biol. 2000, 1, REVIEWS1017. [Google Scholar] [CrossRef]
- Ozakpinar, O.B.; Maurer, A.M.; Ozsavci, D. Ovarian stem cells: From basic to clinical applications. World J. Stem. Cells 2015, 7, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Ratajczak, J.; Suszynska, M.; Miller, D.M.; Kucia, M.; Shin, D.M. A novel view of the adult stem cell compartment from the perspective of a quiescent population of very small embryonic-like stem cells. Circ. Res. 2017, 120, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Sfakianoudis, K.; Simopoulou, M.; Nitsos, N.; Rapani, A.; Pantou, A.; Vaxevanoglou, T.; Kokkali, G.; Koutsilieris, M.; Pantos, K. A case series on platelet-rich plasma revolutionary management of poor responder patients. Gynecol. Obstet. Investig. 2019, 84, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Sills, E.S.; Rickers, N.S.; Svid, C.S.; Rickers, J.M.; Wood, S.H. Normalized ploidy following 20 consecutive blastocysts with chromosomal error: Healthy 46, XY pregnancy with IVF after intraovarian injection of autologous enriched platelet-derived growth factors. Int. J. Mol. Cell. Med. 2019, 8, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Sills, E.S. Why might ovarian rejuvenation fail? Decision analysis of variables impacting reproductive response after autologous platelet-rich plasma. Minerva Obstet. Gynecol. 2022, 74, 377–385. [Google Scholar] [CrossRef]
- Bhartiya, D.; Sharma, D. Ovary does harbor stem cells—Size of the cells matter! J. Ovarian Res. 2020, 13, 39. [Google Scholar] [CrossRef]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11, 1147. [Google Scholar] [CrossRef]
- De Roo, C.; Lierman, S.; Tilleman, K.; Peynshaert, K.; Braeckmans, K.; Caanen, M.; Lambalk, C.B.; Weyers, S.; T’Sjoen, G.; Cornelissen, R.; et al. Ovarian tissue cryopreservation in female-to-male transgender people: Insights into ovarian histology and physiology after prolonged androgen treatment. Reprod. Biomed. Online 2017, 34, 557–566. [Google Scholar] [CrossRef]
- Johnson, B.S.; Krishna, M.B.; Padmanabhan, R.A.; Pillai, S.M.; Jayakrishnan, K.; Laloraya, M. Derailed peripheral circadian genes in polycystic ovary syndrome patients alters peripheral conversion of androgen synthesis. Hum. Reprod. 2022, 37, 1835–1855. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Abudu, A.; Salinas, I.; Sinha, N.; Cline-Fedewa, H.; Yaw, A.M.; Qi, W.; Lydic, T.A.; Takahashi, D.L.; Hennebold, J.D.; et al. Androgen-mediated perturbation of the hepatic circadian system through epigenetic modulation promotes NAFLD in PCOS mice. Endocrinology 2022, 163, bqac127. [Google Scholar] [CrossRef]
- Hikabe, O.; Hamazaki, N.; Nagamatsu, G.; Obata, Y.; Hirao, Y.; Hamada, N.; Shimamoto, S.; Imamura, T.; Nakashima, K.; Saitou, M.; et al. Reconstitution in vitro of the entire cycle of the mouse female germ line. Nature 2016, 539, 299–303. [Google Scholar] [CrossRef]
- Abban, G.; Johnson, J. Stem cell support of oogenesis in the human. Hum. Reprod. 2009, 24, 2974–2978. [Google Scholar] [CrossRef]
- Rickers, N.S.; Sills, E.S. Is Autologous Platelet Activation the Key Step in Ovarian Therapy for Fertility Recovery and Menopause Reversal? Biomedicine 2022, 12, 1–8. [Google Scholar] [CrossRef]
- Suades, R.; Padró, T.; Vilahur, G.; Badimon, L. Platelet-released extracellular vesicles: Effects of thrombin activation. Cell. Mol. Life Sci. 2022, 79, 190. [Google Scholar] [CrossRef]
- Shoeib, H.M.; Keshk, W.A.; Foda, A.M.; Abo El Noeman, S.E.A.E. A study on the regenerative effect of platelet-rich plasma on experimentally induced hepatic damage in albino rats. Can. J. Physiol. Pharmacol. 2018, 96, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, J.; Zhou, F.; Wang, W.; Chen, N. PGC-1α ameliorates kidney fibrosis in mice with diabetic kidney disease through an antioxidative mechanism. Mol. Med. Rep. 2018, 17, 4490–4498. [Google Scholar] [CrossRef]
- Keshk, W.A.; Zahran, S.M. Mechanistic role of cAMP and hepatocyte growth factor signaling in thioacetamide-induced nephrotoxicity: Unraveling the role of platelet rich plasma. Biomed. Pharmacother. 2019, 109, 1078–1084. [Google Scholar] [CrossRef]
- Wood, S.H.; Sills, E.S. Intraovarian vascular enhancement via stromal injection of platelet-derived growth factors: Exploring subsequent oocyte chromosomal status and in vitro fertilization outcomes. Clin. Exp. Reprod. Med. 2020, 47, 94–100. [Google Scholar] [CrossRef]
- Raslan, Z.; Aburima, A.; Naseem, K.M. The spatiotemporal regulation of cAMP signaling in blood platelets—Old friends and new players. Front. Pharmacol. 2015, 6, 266. [Google Scholar] [CrossRef]
- Marschalek, J.; Ott, J.; Aitzetmueller, M.; Mayrhofer, D.; Weghofer, A.; Nouri, K.; Walch, K. The impact of repetitive oocyte retrieval on ovarian reserve: A retrospective cohort study. Arch. Gynecol. Obstet. 2019, 299, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Sills, E.S.; Rickers, N.S.; Petersen, J.L.; Li, X.; Wood, S.H. Regenerative effect of intraovarian injection of autologous platelet rich plasma: Serum anti-Mullerian hormone levels measured among poor-prognosis in vitro fertilization patients. Int. J. Regen. Med. 2020, 3, 1–5. [Google Scholar] [CrossRef]
- Henderson, J.P. Introduction to Thomas Robert Malthus, Principles of political economy, 1820. In Essays in the History of Mainstream Political Economy; Palgrave Macmillan: London, UK, 1992. [Google Scholar] [CrossRef]
- McDonald, P. Low Fertility Not Politically Sustainable; Population Reference Bureau: Washington, DC, USA, 2001; Available online: https://www.prb.org/resources/low-fertility-not-politically-sustainable/ (accessed on 16 January 2023).
- Mauldin, W.P.; Ross, J.A. Prospects and Programs for Fertility Reduction, 1990–2015. Stud. Fam. Plan. 1994, 25, 77–95. [Google Scholar] [CrossRef]
- Leete, R. Fertility Trends and Prospects in East and South-East Asian Countries and Implications for Policies and Programs. Popul. Res. Leads 1991, 39, 1–17. [Google Scholar]
- Liu, Z.; Liu, L. An Analysis of Social Consequences of Rapid Fertility Decline in China. Popul. Res. 1988, 5, 17–30. [Google Scholar] [PubMed]
- Lee, R.; Mason, A.; Amporfu, E.; An, C.-B.; Bixby, L.R.; Bravo, J.; Bucheli, M.; Chen, Q.; Comelatto, P.; Coy, D.; et al. Is low fertility really a problem? Population aging, dependency, and consumption. Science 2014, 346, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Department for Education (United Kingdom). Survey of Childcare and Early Year Providers; Department for Education: London, UK, 2021; pp. 1–23. [Google Scholar]
- Faulconbridge, G. UK Population Growth to Slow Dramatically. Reuters [Wireservice]. 12 January 2022. Available online: https://www.reuters.com/world/uk/uk-population-growth-slow-dramatically-2022-01-12/ (accessed on 16 January 2023).
- Sills, E.S.; Tan, S.L. Preliminary cost variance modeling to compare autologous intraovarian platelet-rich plasma vs. standard hormone replacement therapy for menopause management. Ceska Gynekol. 2022, 87, 28–34. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sills, E.S.; Tan, S.L. Population Dynamics, Plasma Cytokines and Platelet Centrifugation: Technical and Sociodemographic Aspects of ‘Ovarian Rejuvenation’. Clin. Pract. 2023, 13, 435-441. https://doi.org/10.3390/clinpract13020039
Sills ES, Tan SL. Population Dynamics, Plasma Cytokines and Platelet Centrifugation: Technical and Sociodemographic Aspects of ‘Ovarian Rejuvenation’. Clinics and Practice. 2023; 13(2):435-441. https://doi.org/10.3390/clinpract13020039
Chicago/Turabian StyleSills, E. Scott, and Seang Lin Tan. 2023. "Population Dynamics, Plasma Cytokines and Platelet Centrifugation: Technical and Sociodemographic Aspects of ‘Ovarian Rejuvenation’" Clinics and Practice 13, no. 2: 435-441. https://doi.org/10.3390/clinpract13020039
APA StyleSills, E. S., & Tan, S. L. (2023). Population Dynamics, Plasma Cytokines and Platelet Centrifugation: Technical and Sociodemographic Aspects of ‘Ovarian Rejuvenation’. Clinics and Practice, 13(2), 435-441. https://doi.org/10.3390/clinpract13020039