

Network Pharmacology Approaches Used to Identify Therapeutic Molecules for Chronic Venous Disease Based on Potential miRNA Biomarkers



Abstract
1. Introduction
2. Materials and Methods
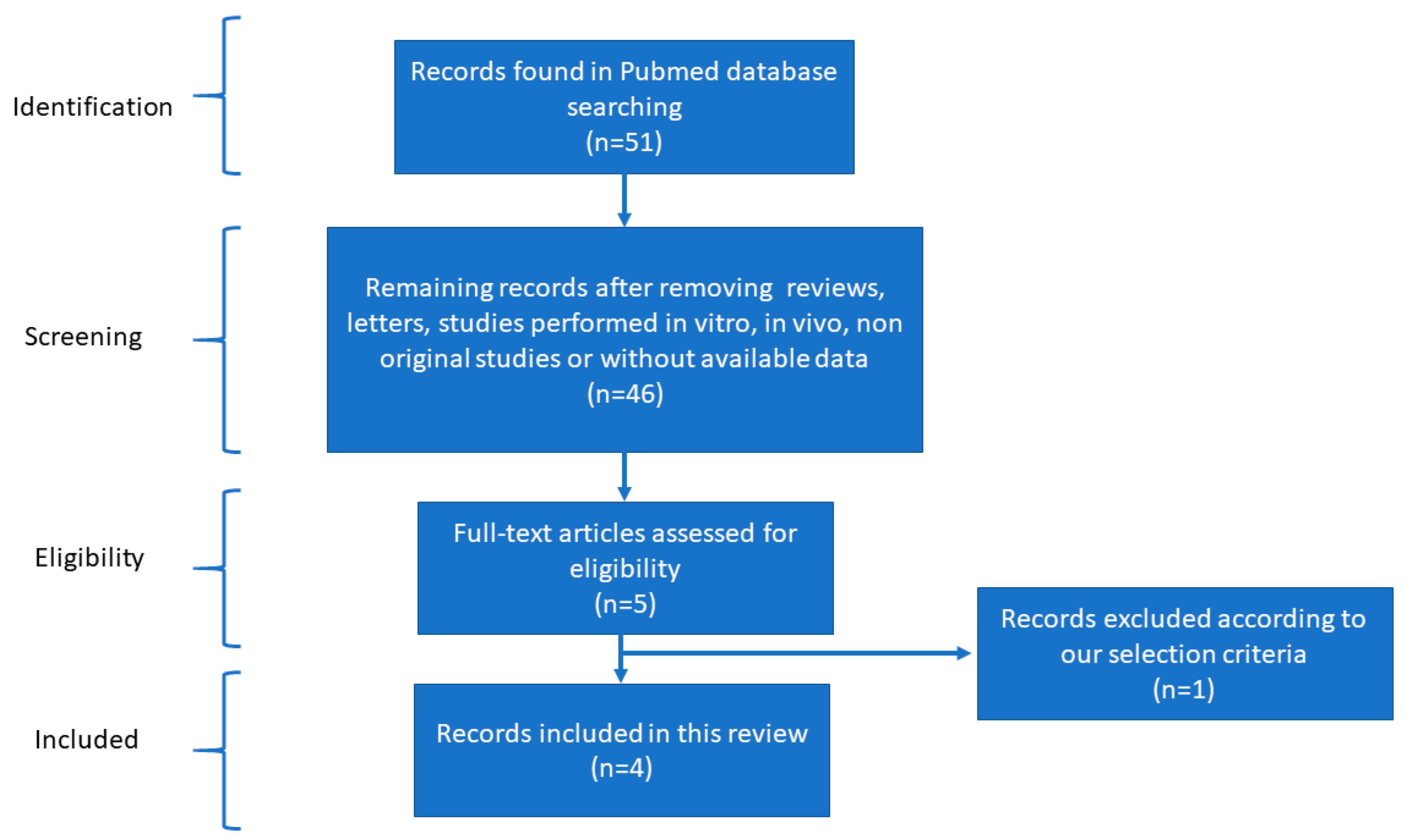
2.1. Reviewing
Bibliographic Screening to Select MicroRNAs Involved in CVD
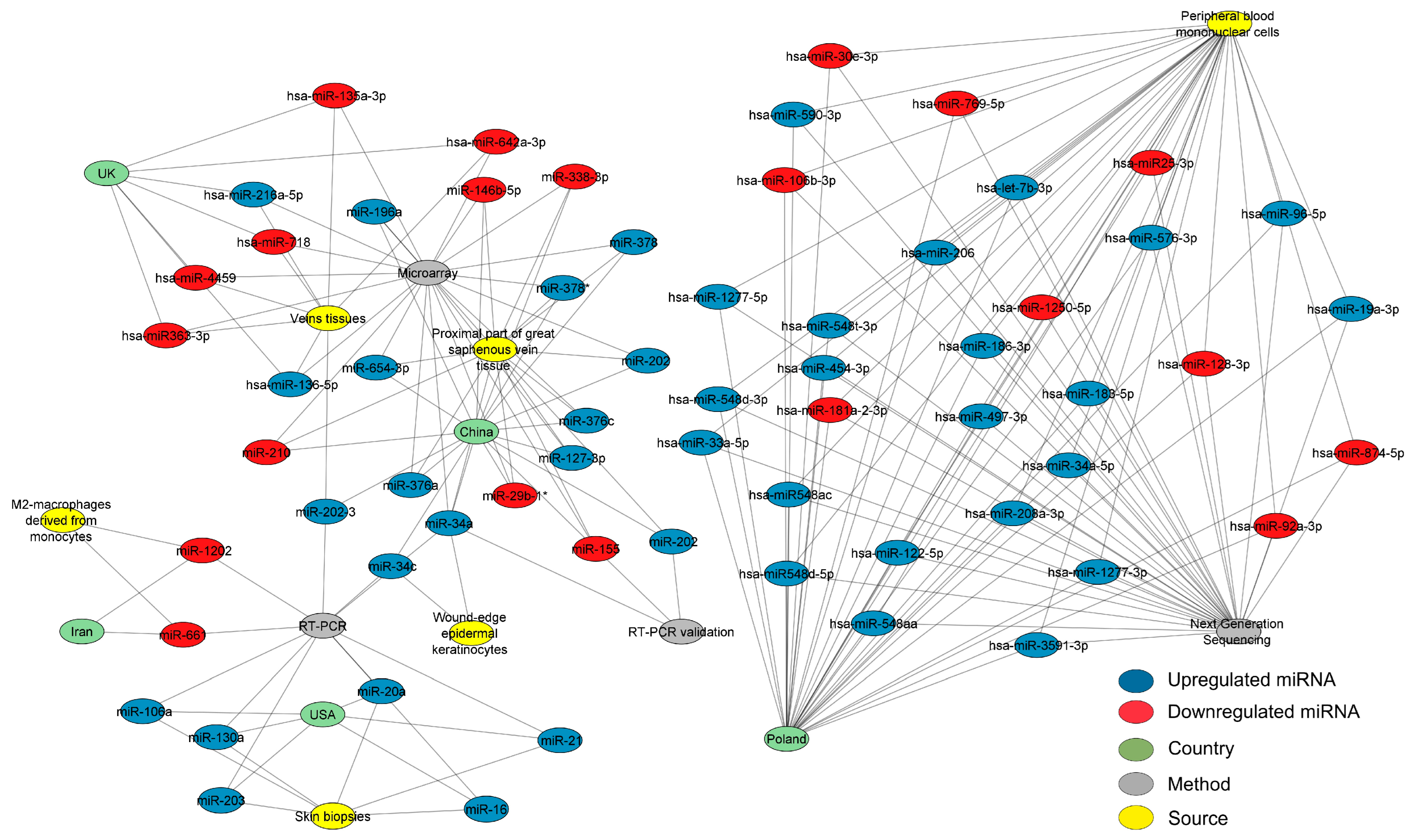
2.2. Data Collection
2.3. Network Structural Analysis
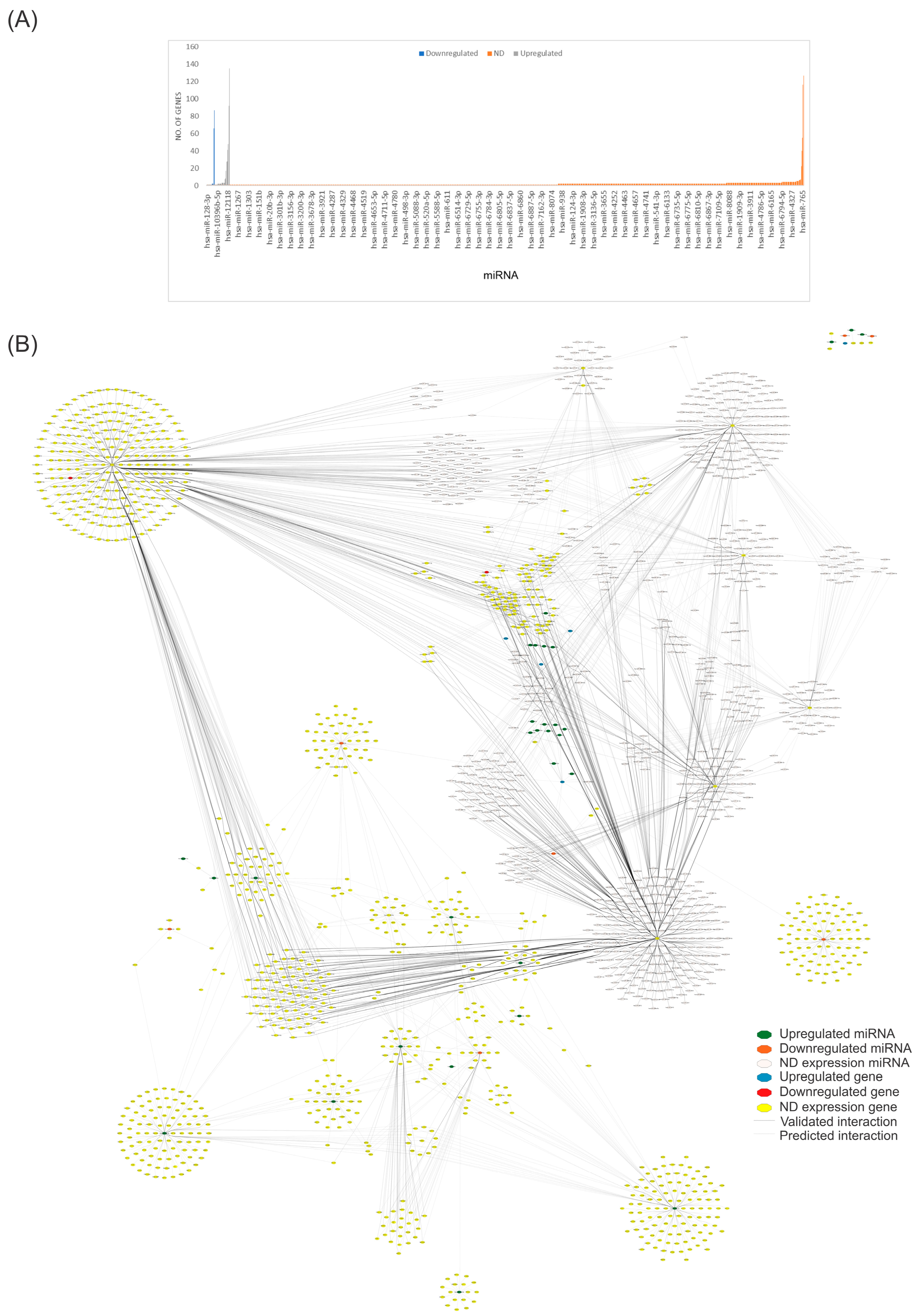
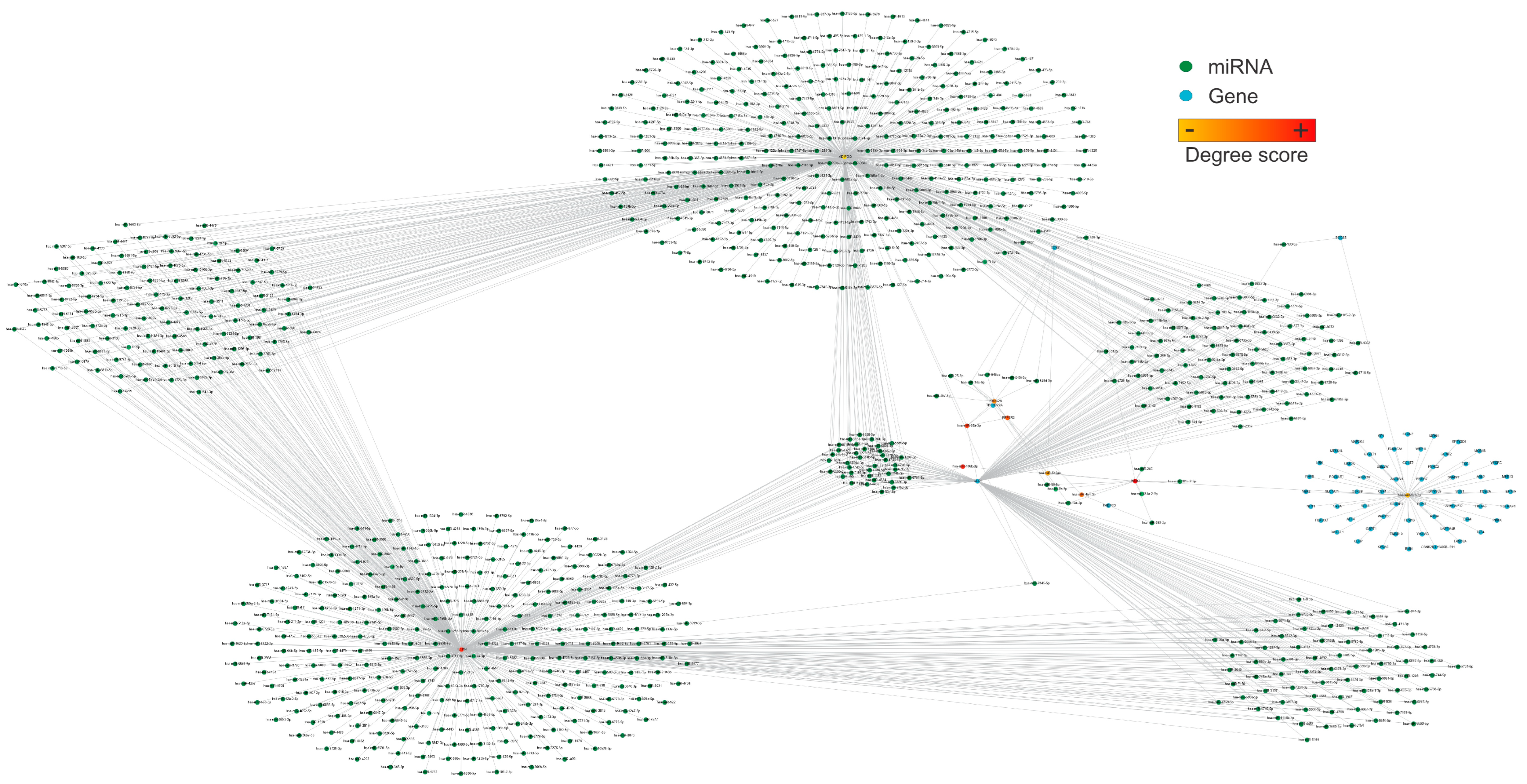
2.4. Network Pharmacology Analysis
2.5. Using QED Index to Determine Drug-like Candidates
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinikumpu, S.P.; Keränen, M.H.; Jokelainen, J.; Keinänen-Kiukaanniemi, S.; Huilaja, L. The association between chronic venous disease and measures of physical performance in older people: A population-based study. BMC Geriatr. 2021, 21, 556. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, A.N. The Most Severe Stage of Chronic Venous Disease: An Update on the Management of Patients with Venous Leg Ulcers. Adv. Ther. 2020, 37, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Duque, M.I.; Yosipovitch, G.; Chan, Y.H.; Smith, R.; Levy, P. Itch, pain, and burning sensation are common symptoms in mild to moderate chronic venous insufficiency with an impact on quality of life. Am. Acad. Dermatol. 2005, 53, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Zalewski, D.P.; Ruszel, K.P.; Stępniewski, A.; Gałkowski, D.; Bogucki, J.; Komsta, Ł.; Kołodziej, P.; Chmiel, P.; Zubilewicz, T.; Feldo, M.; et al. Dysregulations of MicroRNA and Gene Expression in Chronic Venous Disease. J. Clin. Med. 2020, 9, 1251. [Google Scholar] [CrossRef]
- Fernández-Hernando, C.; Suárez, Y. MicroRNAs in endothelial cell homeostasis and vascular disease. Curr. Opin. Hematol. 2018, 25, 227–236. [Google Scholar] [CrossRef]
- Qin, S.; Zhang, C. MicroRNAs in vascular disease. J. Cardiovasc. Pharmacol. 2011, 57, 8–12. [Google Scholar] [CrossRef]
- Pahlavan, Y.; Mohammadi Nasr, M.; Dalir Abdolahinia, E.; Pirdel, Z.; Razi Soofiyani, S.; Siahpoush, S.; Nejati, K. Prominent roles of microRNA-142 in cancer. Pathol. Res. Pract. 2020, 216, 153220. [Google Scholar] [CrossRef]
- Schulte, C.; Karakas, M.; Zeller, T. microRNAs in cardiovascular disease—Clinical application. Clin. Chem. Lab. Med. 2017, 55, 687–704. [Google Scholar] [CrossRef]
- Cui, C.; Liu, G.; Huang, Y.; Lu, X.; Lu, M.; Huang, X.; Li, W.; Jiang, M. MicroRNA profiling in great saphenous vein tissues of patients with chronic venous insufficiency. Tohoku J. Exp. Med. 2012, 228, 341–350. [Google Scholar] [CrossRef]
- Markovic, J.N.; Shortell, C.K. Genomics of varicose veins and chronic venous insufficiency. Semin. Vasc. Surg. 2013, 26, 2–13. [Google Scholar] [CrossRef]
- Pastar, I.; Khan, A.A.; Stojadinovic, O.; Lebrun, E.A.; Medina, M.C.; Brem, H.; Kirsner, R.S.; Jimenez, J.J.; Leslie, C.; Tomic-Canic, M. Induction of specific microRNAs inhibits cutaneous wound healing. J. Biol. Chem. 2012, 287, 29324–29335. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Xu, G.; Huang, J.; Zhou, D.; Huang, X.; Shen, L. Analysis of the association between an insertion/deletion polymorphism within the 3′ untranslated region of COL1A2 and chronic venous insufficiency. Ann. Vasc. Surg. 2013, 27, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Ssempijja, L.; Provenzano, M.; Andreucci, M. Genetic biomarkers in chronic venous disease. Biomark. Med. 2020, 14, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Jiang, N.; Jiang, Q.; Sun, X.; Wang, Y.; Zhang, H.; Hu, Z. Exploring microRNA-like small RNAs in the filamentous fungus Fusarium oxysporum. PLoS ONE 2014, 9, e104956. [Google Scholar] [CrossRef] [PubMed]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta 2010, 1803, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Galvão-Lima, L.J.; Morais, A.H.F.; Valentim, R.A.M.; Barreto, E. miRNAs as biomarkers for early cancer detection and their application in the development of new diagnostic tools. Biomed. Eng. Online 2021, 20, 21. [Google Scholar] [CrossRef]
- Backes, C.; Meese, E.; Keller, A. Specific miRNA Disease Biomarkers in Blood, Serum and Plasma: Challenges and Prospects. Mol. Diagn. Ther. 2016, 20, 509–518. [Google Scholar] [CrossRef]
- Izzotti, A.; Carozzo, S.; Pulliero, A.; Zhabayeva, D.; Ravetti, J.L.; Bersimbaev, R. Extracellular MicroRNA in liquid biopsy: Applicability in cancer diagnosis and prevention. Am. J. Cancer Res. 2016, 6, 1461. [Google Scholar]
- Shigeyasu, K.; Toden, S.; Zumwalt, T.J.; Okugawa, Y.; Goel, A. Emerging role of microRNAs as liquid biopsy biomarkers in gastrointestinal cancers. Clin. Cancer Res. 2017, 23, 2391–2399. [Google Scholar] [CrossRef]
- Yu, D.; Kim, M.; Xiao, G.; Hwang, T.H. Review of biological network data and its applications. Genom. Inform. 2013, 11, 200–210. [Google Scholar] [CrossRef]
- Charitou, T.; Bryan, K.; Lynn, D.J. Using biological networks to integrate, visualize and analyze genomics data. Genet. Sel. Evol. 2016, 48, 27. [Google Scholar] [CrossRef] [PubMed]
- Barabási, A.-L.; Gulbahce, N.; Loscalzo, J. Network medicine: A network-based approach to human disease. Nat. Rev. Genet. 2011, 12, 56–68. [Google Scholar] [CrossRef]
- Lee, T.J.; Yuan, X.; Kerr, K.; Yoo, J.Y.; Kim, D.H.; Kaur, B.; Eltzschig, H.K. Strategies to Modulate MicroRNA Functions for the Treatment of Cancer or Organ Injury. Pharmacol. Rev. 2020, 72, 639–667. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Ijaz, B.; Ahmad, Z.; Farooq, N.; Sarwar, M.B.; Husnain, T. Modification of miRNA Expression through plant extracts and compounds against breast cancer: Mechanism and translational significance. Phytomedicine 2020, 68, 153168. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, E.M.; Ragle Aure, M.; Haugen, M.H.; Kleivi Sahlberg, K.; Kristensen, V.N.; Nebdal, D.; Børresen-Dale, A.L.; Lingjaerde, O.C.; Engebraaten, O. miRNA expression changes during the course of neoadjuvant bevacizumab and chemotherapy treatment in breast cancer. Mol. Oncol. 2019, 13, 2278–2296. [Google Scholar] [CrossRef]
- Arora, A. MicroRNA targets: Potential candidates for indirect regulation by drugs. Pharmacogenet. Genom. 2015, 25, 107–125. [Google Scholar] [CrossRef]
- Cervena, K.; Novosadova, V.; Pardini, B.; Naccarati, A.; Opattova, A.; Horak, J.; Vodenkova, S.; Buchler, T.; Skrobanek, P.; Levy, M.; et al. Analysis of MicroRNA Expression Changes During the Course of Therapy In Rectal Cancer Patients. Front. Oncol. 2021, 11, 702258. [Google Scholar] [CrossRef]
- Santiago, F. Quality of Life in Chronic Venous Disease: Bridging the Gap Between Patients and Physicians. Clin. Drug Investig. 2023, 43, 3–8. [Google Scholar] [CrossRef]
- Rabe, E.; Guex, J.-J.; Morrison, N.; Ramelet, A.-A.; Schuller-Petrovic, S.; Scuderi, A.; Staelens, I.; Pannier, F. Treatment of chronic venous disease with flavonoids: Recommendations for treatment and further studies. Phlebology 2013, 28, 308–319. [Google Scholar] [CrossRef]
- Martinez-Zapata, M.J.; Vernooij, R.W.; Simancas-Racines, D.; Uriona Tuma, S.M.; Stein, A.T.; Moreno Carriles, R.M.M.; Vargas, E.; Bonfill Cosp, X. Phlebotonics for venous insufficiency. Cochrane Database Syst. Rev. 2020, 11, Cd003229. [Google Scholar] [CrossRef]
- Imperatrice, M.; Cuijpers, I.; Troost, F.J.; Sthijns, M.M.J.P.E. Hesperidin Functions as an Ergogenic Aid by Increasing Endothelial Function and Decreasing Exercise-Induced Oxidative Stress and Inflammation, Thereby Contributing to Improved Exercise Performance. Nutrients 2022, 14, 2955. [Google Scholar] [CrossRef] [PubMed]
- Magura, J.; Moodley, R.; Mackraj, I. The effect of hesperidin and luteolin isolated from Eriocephalus africanus on apoptosis, cell cycle and miRNA expression in MCF-7. J. Biomol. Struct. Dyn. 2022, 40, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Zhou, S.; Liu, M.; An, H.; Wang, Z.; Liu, T. Clinical Significance of miR-21-5p in Predicting Occurrence and Progression of Uremic Vascular Calcification in Patients with End-Stage Renal Disease. Yonsei Med. J. 2022, 63, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Toro, R.; Pérez-Serra, A.; Mangas, A.; Campuzano, O.; Sarquella-Brugada, G.; Quezada-Feijoo, M.; Ramos, M.; Alcalá, M.; Carrera, E.; García-Padilla, C.; et al. miR-16-5p Suppression Protects Human Cardiomyocytes against Endoplasmic Reticulum and Oxidative Stress-Induced Injury. Int. J. Mol. Sci. 2022, 23, 1036. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.C.; Liu, X.M.; Liang, L.R.; Wang, L.F.; Zhong, J.C. Targeting the microRNA-34a as a Novel Therapeutic Strategy for Cardiovascular Diseases. Front. Cardiovasc. Med. 2021, 8, 784044. [Google Scholar] [CrossRef]
- Wu, J.; Li, X.; Li, D.; Ren, X.; Li, Y.; Herter, E.K.; Qian, M.; Toma, M.A.; Wintler, A.M.; Sérézal, I.G.; et al. MicroRNA-34 Family Enhances Wound Inflammation by Targeting LGR4. J. Investig. Dermatol. 2020, 140, 465–476.e411. [Google Scholar] [CrossRef]
- Biranvand, A.S.; Khosravi, M.; Esfandiari, G.; Poursaleh, A.; Hosseini-Fard, S.R.; Amirfarhangi, A.; Najafi, M. Associations between miR-661, miR-1202, lncRNA-HOTAIR, lncRNA-GAS5 and MMP9 in differentiated M2-macrophages of patients with varicose veins. Int. Angiol. 2018, 37, 451–456. [Google Scholar] [CrossRef]
- Huang, X.; Liu, Z.; Shen, L.; Jin, Y.; Xu, G.; Zhang, Z.; Fang, C.; Guan, W.; Liu, C. Augmentation of miR-202 in varicose veins modulates phenotypic transition of vascular smooth muscle cells by targeting proliferator-activated receptor-γ coactivator-1α. J. Cell. Biochem. 2019, 120, 10031–10042. [Google Scholar] [CrossRef]
- Anwar, M.A.; Adesina-Georgiadis, K.N.; Spagou, K.; Vorkas, P.A.; Li, J.V.; Shalhoub, J.; Holmes, E.; Davies, A.H. A comprehensive characterisation of the metabolic profile of varicose veins; implications in elaborating plausible cellular pathways for disease pathogenesis. Sci. Rep. 2017, 7, 2989. [Google Scholar] [CrossRef]
- Ortega, M.A.; Fraile-Martínez, O.; García-Montero, C.; Álvarez-Mon, M.A.; Chaowen, C.; Ruiz-Grande, F.; Pekarek, L.; Monserrat, J.; Asúnsolo, A.; García-Honduvilla, N.; et al. Understanding Chronic Venous Disease: A Critical Overview of Its Pathophysiology and Medical Management. J. Clin. Med. 2021, 10, 3239. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8 (Suppl. 4), S11. [Google Scholar] [CrossRef] [PubMed]
- Winterhalter, C.; Widera, P.; Krasnogor, N. JEPETTO: A Cytoscape plugin for gene set enrichment and topological analysis based on interaction networks. Bioinformatics 2014, 30, 1029–1030. [Google Scholar] [CrossRef] [PubMed]
- Astore, C.; Zhou, H.; Ilkowski, B.; Forness, J.; Skolnick, J. LeMeDISCO is a computational method for large-scale prediction & molecular interpretation of disease comorbidity. Commun. Biol. 2022, 5, 870. [Google Scholar] [CrossRef]
- Griffiths-Jones, S. miRBase: The microRNA sequence database. Methods Mol. Biol. 2006, 342, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Li, B.; Sun, J.; Qi, J.; De Wilde, R.L.; Torres-de la Roche, L.A.; Li, C.; Ahmad, S.; Shi, W.; Li, X.; et al. PSRR: A Web Server for Predicting the Regulation of miRNAs Expression by Small Molecules. Front. Mol. Biosci. 2022, 9, 817294. [Google Scholar] [CrossRef]
- Bickerton, G.R.; Paolini, G.V.; Besnard, J.; Muresan, S.; Hopkins, A.L. Quantifying the chemical beauty of drugs. Nat. Chem. 2012, 4, 90–98. [Google Scholar] [CrossRef]
- Barrera-Vázquez, O.S.; Montenegro-Herrera, S.A.; Martínez-Enríquez, M.E.; Escobar-Ramírez, J.L.; Magos-Guerrero, G.A. Selection of Mexican Medicinal Plants by Identification of Potential Phytochemicals with Anti-Aging, Anti-Inflammatory, and Anti-oxidant Properties through Network Analysis and Chemoinformatic Screening. Biomolecules 2023, 13, 1673. [Google Scholar] [CrossRef]
- Rathod, N.B.; Elabed, N.; Punia, S.; Ozogul, F.; Kim, S.K.; Rocha, J.M. Recent Developments in Polyphenol Applications on Human Health: A Review with Current Knowledge. Plants 2023, 12, 1217. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Safa, A.; Bahroudi, Z.; Shoorei, H.; Majidpoor, J.; Abak, A.; Taheri, M.; Ghafouri-Fard, S. miR-1: A comprehensive review of its role in normal development and diverse disorders. Biomed. Pharmacother. 2020, 132, 110903. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Guo, Q.; Fu, F.J.; Wang, Z.; Yin, Z.; Wei, Y.B.; Yang, J.R. The role of miR-29b in cancer: Regulation, function, and signaling. OncoTargets Ther. 2015, 8, 539–548. [Google Scholar] [CrossRef]
- Katoh, M. Cardio-miRNAs and onco-miRNAs: Circulating miRNA-based diagnostics for non-cancerous and cancerous diseases. Front. Cell Dev. Biol. 2014, 2, 61. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.; Yao, X.; Engel, T.; Tiwari, D.; Xing, L.; Rowley, S.; Danielson, S.W.; Thomas, K.T.; Jimenez-Mateos, E.M.; Schroeder, L.M.; et al. MicroRNA-Mediated Downregulation of the Potassium Channel Kv4.2 Contributes to Seizure Onset. Cell Rep. 2016, 17, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Smalheiser, N.R.; Lugli, G.; Rizavi, H.S.; Torvik, V.I.; Turecki, G.; Dwivedi, Y. MicroRNA expression is down-regulated and reorganized in prefrontal cortex of depressed suicide subjects. PLoS ONE 2012, 7, e33201. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, D.L.; Long, B.; An, T.; Zhang, J.; Zhou, L.Y.; Liu, C.Y.; Li, P.F. NFAT4-dependent miR-324-5p regulates mitochondrial morphology and cardiomyocyte cell death by targeting Mtfr1. Cell Death Dis. 2015, 6, e2007. [Google Scholar] [CrossRef]
- Hervé, M.; Ibrahim, E.C. MicroRNA screening identifies a link between NOVA1 expression and a low level of IKAP in familial dysautonomia. Dis. Model. Mech. 2016, 9, 899–909. [Google Scholar] [CrossRef]
- Mas-Capdevila, A.; Teichenne, J.; Domenech-Coca, C.; Caimari, A.; Del Bas, J.M.; Escoté, X.; Crescenti, A. Effect of Hesperidin on Cardiovascular Disease Risk Factors: The Role of Intestinal Microbiota on Hesperidin Bioavailability. Nutrients 2020, 12, 1488. [Google Scholar] [CrossRef]
- Bian, H.; Zhou, Y.; Zhou, D.; Zhang, Y.; Shang, D.; Qi, J. The latest progress on miR-374 and its functional implications in physiological and pathological processes. J. Cell. Mol. Med. 2019, 23, 3063–3076. [Google Scholar] [CrossRef]
- Di Mauro, G.; Frontini, F.; Torreggiani, E.; Iaquinta, M.R.; Caselli, A.; Mazziotta, C.; Esposito, V.; Mazzoni, E.; Libener, R.; Grosso, F.; et al. Epigenetic investigation into circulating microRNA 197-3p in sera from patients affected by malignant pleural mesothelioma and workers ex-exposed to asbestos. Sci. Rep. 2023, 13, 6501. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Jung, C.H.; Ha, S.Y.; Seo, H.D.; Kim, Y.I.; Ha, T. 6-Gingerol Ameliorates Hepatic Steatosis via HNF4α/miR-467b-3p/GPAT1 Cascade. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1201–1213. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wang, X.; Zhao, H.; Jiang, F.; Li, Y.; Yao, Y.; Shi, C.; Yang, Y. MiR-291a/b-5p inhibits autophagy by targeting Atg5 and Becn1 during mouse preimplantation embryo development. RSC Adv. 2019, 9, 9331–9341. [Google Scholar] [CrossRef] [PubMed]
- Rider, B.J. 5 Fluorouracil. In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: New York, NY, USA, 2007; pp. 1–5. [Google Scholar]
- Hantzidiamantis, P.J.; Awosika, A.O.; Lappin, S.L. Physiology, Glucose. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/books/NBK545201/ (accessed on 22 July 2024).
- Manohar, S.; Leung, N. Cisplatin nephrotoxicity: A review of the literature. J. Nephrol. 2018, 31, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yu, S.; Xing, X.; Qiao, J.; Yin, Y.; Wang, J.; Liu, M.; Zhang, W. Ginsenoside Rh2 stimulates the production of mitochondrial reactive oxygen species and induces apoptosis of cervical cancer cells by inhibiting mitochondrial electron transfer chain complex. Mol. Med. Rep. 2021, 24, 873. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, L.; Barbosa, E.; Charão, M.F.; Brucker, N. Formaldehyde toxicity reports from in vitro and in vivo studies: A review and updated data. Drug Chem. Toxicol. 2022, 45, 972–984. [Google Scholar] [CrossRef]
- Reed, C.E.; Fenton, S.E. Exposure to diethylstilbestrol during sensitive life stages: A legacy of heritable health effects. Birth Defects Res. Part C Embryo Today Rev. 2013, 99, 134–146. [Google Scholar] [CrossRef]
- Jałbrzykowska, K.; Chrzanowska, A.; Roszkowski, P.; Struga, M. The New Face of a Well-Known Antibiotic: A Review of the Anticancer Activity of Enoxacin and Its Derivatives. Cancers 2022, 14, 3056. [Google Scholar] [CrossRef]
- Zhou, H.; Xie, Y.; Baloch, Z.; Shi, Q.; Huo, Q.; Ma, T. The effect of atorvastatin, 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitor (HMG-CoA), on the prevention of osteoporosis in ovariectomized rabbits. J. Bone Miner. Metab. 2017, 35, 245–254. [Google Scholar] [CrossRef]
- Jiang, Y.; Shen, X.; Zhi, F.; Wen, Z.; Gao, Y.; Xu, J.; Yang, B.; Bai, Y. An overview of arsenic trioxide-involved combined treatment algorithms for leukemia: Basic concepts and clinical implications. Cell Death Discov. 2023, 9, 266. [Google Scholar] [CrossRef]
- Le, V.K.H.; Pham, T.P.D.; Truong, D.H. Delivery systems for vorinostat in cancer treatment: An updated review. J. Drug Deliv. Technol. 2021, 61, 102334. [Google Scholar] [CrossRef]
- Yaşar, P.; Ayaz, G.; User, S.D.; Güpür, G.; Muyan, M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod. Med. Biol. 2017, 16, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Ping, Y.H.; Lee, H.C.; Lee, J.Y.; Wu, P.H.; Ho, L.K.; Chi, C.W.; Lu, M.F.; Wang, J.J. Anticancer effects of low-dose 10-hydroxycamptothecin in human colon cancer. Oncol. Rep. 2006, 15, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Sritharan, S.; Sivalingam, N. A comprehensive review on time-tested anticancer drug doxorubicin. Life Sci. 2021, 278, 119527. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Herrera-Bravo, J.; Kamiloglu, S.; Petroni, K.; Mishra, A.P.; Monserrat-Mesquida, M.; Sureda, A.; Martorell, M.; Aidarbekovna, D.S.; Yessimsiitova, Z.; et al. Recent advances in the therapeutic potential of emodin for human health. Biomed. Pharmacother. 2022, 154, 113555. [Google Scholar] [CrossRef] [PubMed]
- Abdulfattah, A.Y.; Tajuddin, S.; Akkari, N.; Elsayed, O.I.; Graham-Hill, S. A Rare Case of Bicalutamide-Induced Severe Congestive Heart Failure in a Patient With Advanced Prostate Cancer. Cureus 2024, 16, e60298. [Google Scholar] [CrossRef]
- Bouyahya, A.; El Omari, N.; Bakha, M.; Aanniz, T.; El Menyiy, N.; El Hachlafi, N.; El Baaboua, A.; El-Shazly, M.; Alshahrani, M.M.; Al Awadh, A.A.; et al. Pharmacological Properties of Trichostatin A, Focusing on the Anticancer Potential: A Comprehensive Review. Pharmaceuticals 2022, 15, 1235. [Google Scholar] [CrossRef]
- Combs, G.F. Chapter 9—Vitamin C. In The Vitamins, 4th ed.; Combs, G.F., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 233–259. [Google Scholar]
- Felippe, M.J.B. Chapter 66—Immunotherapy. In Equine Infectious Diseases, 2nd ed.; Sellon, D.C., Long, M.T., Eds.; W.B. Saunders: St. Louis, MO, USA, 2014; pp. 584–597.e585. [Google Scholar]
- Liang, C.; Qiao, G.; Liu, Y.; Tian, L.; Hui, N.; Li, J.; Ma, Y.; Li, H.; Zhao, Q.; Cao, W.; et al. Overview of all-trans-retinoic acid (ATRA) and its analogues: Structures, activities, and mechanisms in acute promyelocytic leukaemia. Eur. J. Med. Chem. 2021, 220, 113451. [Google Scholar] [CrossRef]
- Majeed, M.; Hakeem, K.R.; Rehman, R.U. Mistletoe lectins: From interconnecting proteins to potential tumour inhibiting agents. Phytomed. Plus 2021, 1, 100039. [Google Scholar] [CrossRef]
- Fan, G.; Wei, X.; Xu, X. Is the era of sorafenib over? A review of the literature. Ther. Adv. Med. Oncol. 2020, 12, 1758835920927602. [Google Scholar] [CrossRef]
- Saleem, K.; Franz, J.; Klem, M.L.; Yabes, J.G.; Boyiadzis, M.; Jones, J.R.; Shaikh, N.; Lontos, K. Second primary malignancies in patients with haematological cancers treated with lenalidomide: A systematic review and meta-analysis. Lancet Haematol. 2022, 9, e906–e918. [Google Scholar] [CrossRef]
- Kirstine, W.V.; Galbally, I.E. Ethanol in the Environment: A Critical Review of Its Roles as a Natural Product, a Biofuel, and a Potential Environmental Pollutant. Crit. Rev. Environ. Sci. Technol. 2012, 42, 1735–1779. [Google Scholar] [CrossRef]
- Lyseng-Williamson, K.A.; Fenton, C. Docetaxel: A review of its use in metastatic breast cancer. Drugs 2005, 65, 2513–2531. [Google Scholar] [CrossRef]
- Pourmadadi, M.; Aslani, A.; Hassan, D.; Sani, A.; Rahdar, A.; Medina, D.I.; Abdouss, M.; Romanholo Ferreira, L.F. Recent advancements in the targeted delivery of Gemcitabine: Harnessing nanomedicine for enhanced cancer therapy. OpenNano 2023, 13, 100177. [Google Scholar] [CrossRef]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and safety of trastuzumab as a single agent in first-line treatment of HER2-overexpressing metastatic breast cancer. J. Clin. Oncol. 2002, 20, 719–726. [Google Scholar] [CrossRef]
- Zanwar, A.A.; Badole, S.L.; Shende, P.S.; Hegde, M.V.; Bodhankar, S.L. Chapter 21—Anti-oxidant Role of Catechin in Health and Disease. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 267–271. [Google Scholar]
- Jeong, S.P.; Sharma, N.; An, S.S.A. Role of Calcitriol and Vitamin D Receptor (VDR) Gene Polymorphisms in Alzheimer’s Disease. Int. J. Mol. Sci. 2024, 25, 4806. [Google Scholar] [CrossRef] [PubMed]
- Kolatorova, L.; Vitku, J.; Suchopar, J.; Hill, M.; Parizek, A. Progesterone: A Steroid with Wide Range of Effects in Physiology as Well as Human Medicine. Int. J. Mol. Sci. 2022, 23, 7989. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Chakrabortty, A.; Patton, D.J.; Smith, B.F.; Agarwal, P. miRNAs: Potential as Biomarkers and Therapeutic Targets for Cancer. Genes 2023, 14, 1375. [Google Scholar] [CrossRef]
- Zalewski, D.; Chmiel, P. MicroRNA expression biomarkers of chronic venous disease. Curr. Issues Pharm. Med. Sci. 2022, 35, 21–26. [Google Scholar] [CrossRef]
- Raucci, A.; Macrì, F.; Castiglione, S.; Badi, I.; Vinci, M.C.; Zuccolo, E. MicroRNA-34a: The bad guy in age-related vascular diseases. Cell. Mol. Life Sci. 2021, 78, 7355–7378. [Google Scholar] [CrossRef]
- Guo, Y.; Xing, L.; Chen, N.; Gao, C.; Ding, Z.; Jin, B. Total flavonoids from the Carya cathayensis Sarg. leaves inhibit HUVEC senescence through the miR-34a/SIRT1 pathway. J. Cell. Biochem. 2019, 120, 17240–17249. [Google Scholar] [CrossRef] [PubMed]
- Niederer, F.; Trenkmann, M.; Ospelt, C.; Karouzakis, E.; Neidhart, M.; Stanczyk, J.; Kolling, C.; Gay, R.E.; Detmar, M.; Gay, S.; et al. Down-regulation of microRNA-34a* in rheumatoid arthritis synovial fibroblasts promotes apoptosis resistance. Arthritis Rheum. 2012, 64, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Ducasse, E.; Giannakakis, K.; Speziale, F.; Midy, D.; Sbarigia, E.; Baste, J.C.; Faraggiana, T. Association of primary varicose veins with dysregulated vein wall apoptosis. Eur. J. Vasc. Endovasc. Surg. 2008, 35, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Mikuła-Pietrasik, J.; Uruski, P.; Aniukiewicz, K.; Sosińska, P.; Krasiński, Z.; Tykarski, A.; Książek, K. Serum from Varicose Patients Induces Senescence-Related Dysfunction of Vascular Endothelium Generating Local and Systemic Proinflammatory Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 2069290. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.B.; An, Z.; Zheng, H.K.; Wang, X.P.; Shan, R.T.; Mao, C.Y.; Zhang, W.Q. MicroRNA-34c suppresses proliferation of vascular smooth muscle cell via modulating high mobility group box protein 1. J. Clin. Lab. Anal. 2020, 34, e23293. [Google Scholar] [CrossRef]
- Kim, S.; Han, J.; Ahn, Y.H.; Ha, C.H.; Hwang, J.J.; Lee, S.E.; Kim, J.J.; Kim, N. Protective Role of miR-34c in Hypoxia by Activating Autophagy through BCL2 Repression. Mol. Cells 2022, 45, 403–412. [Google Scholar] [CrossRef]
- Harati, R.; Hafezi, S.; Mabondzo, A.; Tlili, A. Silencing miR-202-3p increases MMP-1 and promotes a brain invasive phenotype in metastatic breast cancer cells. PLoS ONE 2020, 15, e0239292. [Google Scholar] [CrossRef]
- Papazafiropoulou, A.; Tentolouris, N. Matrix metalloproteinases and cardiovascular diseases. Hippokratia 2009, 13, 76–82. [Google Scholar]
- Dbouk, H.A.; Weil, L.M.; Perera, G.K.; Dellinger, M.T.; Pearson, G.; Brekken, R.A.; Cobb, M.H. Actions of the protein kinase WNK1 on endothelial cells are differentially mediated by its substrate kinases OSR1 and SPAK. Proc. Natl. Acad. Sci. USA 2014, 111, 15999–16004. [Google Scholar] [CrossRef]
- Shiiba, M.; Saito, K.; Yamagami, H.; Nakashima, D.; Higo, M.; Kasamatsu, A.; Sakamoto, Y.; Ogawara, K.; Uzawa, K.; Takiguchi, Y.; et al. Interleukin-1 receptor antagonist (IL1RN) is associated with suppression of early carcinogenic events in human oral malignancies. Int. J. Oncol. 2015, 46, 1978–1984. [Google Scholar] [CrossRef]
- Tavernier, N.; Sicheri, F.; Pintard, L. Aurora A kinase activation: Different means to different ends. J. Cell Biol. 2021, 220, e202106128. [Google Scholar] [CrossRef]
- Al-Nbaheen, M.S. Effect of Genetic Variations in the ADIPOQ Gene on Susceptibility to Type 2 Diabetes Mellitus. Diabetes Metab. Syndr. Obes. 2022, 15, 2753–2761. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, H.; Hwang, K.H.; Chang, J.S.; Park, K.S.; Cha, S.K.; Kong, I.D. WNK1 kinase is essential for insulin-stimulated GLUT4 trafficking in skeletal muscle. FEBS Open Bio 2018, 8, 1866–1874. [Google Scholar] [CrossRef]
- Henriques, A.F.A.; Matos, P.; Carvalho, A.S.; Azkargorta, M.; Elortza, F.; Matthiesen, R.; Jordan, P. WNK1 phosphorylation sites in TBC1D1 and TBC1D4 modulate cell surface expression of GLUT1. Arch. Biochem. Biophys. 2020, 679, 108223. [Google Scholar] [CrossRef] [PubMed]
- Jaykumar, A.B.; Plumber, S.; Barry, D.M.; Binns, D.; Wichaidit, C.; Grzemska, M.; Earnest, S.; Goldsmith, E.J.; Cleaver, O.; Cobb, M.H. WNK1 collaborates with TGF-β in endothelial cell junction turnover and angiogenesis. Proc. Natl. Acad. Sci. USA 2022, 119, e2203743119. [Google Scholar] [CrossRef] [PubMed]
- Fejfarová, V.; Roztočil, K.; Svědínková, A.; Čermáková, H.; Dubský, M.; Bém, R.; Jirkovská, A.; Wosková, V.; Němcová, A.; Skibová, J. The relationship between chronic venous insufficiency and diabetes mellitus. Int. Angiol. 2017, 36, 90–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, Y.; He, X.; Yang, P.; Zong, T.; Sun, P.; Sun, R.C.; Yu, T.; Jiang, Z. The cellular function and molecular mechanism of formaldehyde in cardiovascular disease and heart development. J. Cell. Mol. Med. 2021, 25, 5358–5371. [Google Scholar] [CrossRef]
- Augenreich, M.; Stickford, J.; Stute, N.; Koontz, L.; Cope, J.; Bennett, C.; Ratchford, S.M. Vascular dysfunction and oxidative stress caused by acute formaldehyde exposure in female adults. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H1369–H1379. [Google Scholar] [CrossRef]
- Wang, S.; Han, Q.; Wei, Z.; Wang, Y.; Deng, L.; Chen, M. Formaldehyde causes an increase in blood pressure by activating ACE/AT1R axis. Toxicology 2023, 486, 153442. [Google Scholar] [CrossRef]
- Bergan, J.J.; Pascarella, L.; Schmid-Schönbein, G.W. Pathogenesis of primary chronic venous disease: Insights from animal models of venous hypertension. J. Vasc. Surg. 2008, 47, 183–192. [Google Scholar] [CrossRef]
- Raju, S.; Knight, A.; Lamanilao, L.; Pace, N.; Jones, T. Peripheral venous hypertension in chronic venous disease. J. Vasc. Surg. Venous Lymphat. Disord. 2019, 7, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Perrin, M.; Ramelet, A.A. Pharmacological Treatment of Primary Chronic Venous Disease: Rationale, Results and Unanswered Questions. Eur. J. Vasc. Endovasc. Surg. 2011, 41, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Santillán-Aguayo, E.; Rivera Sánchez, J.d.J.; Carbajal-Robles, V.; Duarte-Acuña, J.; Piña-Avilés, F.A.; Ramírez-Berumen, M.V. Chronic venous insufficiency symptoms and its potential causes: Are we doing it right? Rev. Mex. Angiol. 2021, 49, 57–66. [Google Scholar] [CrossRef]
- Awata, W.M.C.; Alves, J.V.; Costa, R.M.; Bruder-Nascimento, A.; Singh, S.; Barbosa, G.S.; Tirapelli, C.R.; Bruder-Nascimento, T. Vascular injury associated with ethanol intake is driven by AT1 receptor and mitochondrial dysfunction. Biomed. Pharmacother. 2023, 169, 115845. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Gallelli, L.; Perri, P.; De Francesco, E.M.; Rigiracciolo, D.C.; Mastroroberto, P.; Maggiolini, M.; de Franciscis, S. Estrogen Receptors and Chronic Venous Disease. Eur. J. Vasc. Endovasc. Surg. 2016, 52, 114–118. [Google Scholar] [CrossRef]
- Kalfert, D.; Ludvikova, M.; Pesta, M.; Ludvik, J.; Dostalova, L.; Kholová, I. Multifunctional Roles of miR-34a in Cancer: A Review with the Emphasis on Head and Neck Squamous Cell Carcinoma and Thyroid Cancer with Clinical Implications. Diagnostics 2020, 10, 563. [Google Scholar] [CrossRef]
- Fu, J.; Imani, S.; Wu, M.Y.; Wu, R.C. MicroRNA-34 Family in Cancers: Role, Mechanism, and Therapeutic Potential. Cancers 2023, 15, 4723. [Google Scholar] [CrossRef]
- Takashima, Y.; Kawaguchi, A.; Iwadate, Y.; Hondoh, H.; Fukai, J.; Kajiwara, K.; Hayano, A.; Yamanaka, R. miR-101, miR-548b, miR-554, and miR-1202 are reliable prognosis predictors of the miRNAs associated with cancer immunity in primary central nervous system lymphoma. PLoS ONE 2020, 15, e0229577. [Google Scholar] [CrossRef]
- Lohcharoenkal, W.; Li, C.; Das Mahapatra, K.; Lapins, J.; Homey, B.; Sonkoly, E.; Pivarcsi, A. MiR-130a Acts as a Tumor Suppressor MicroRNA in Cutaneous Squamous Cell Carcinoma and Regulates the Activity of the BMP/SMAD Pathway by Suppressing ACVR1. J. Investig. Dermatol. 2021, 141, 1922–1931. [Google Scholar] [CrossRef]
- Sagar, S.K. miR-106b as an emerging therapeutic target in cancer. Genes. Dis. 2022, 9, 889–899. [Google Scholar] [CrossRef]
- Larrue, R.; Fellah, S.; Boukrout, N.; De Sousa, C.; Lemaire, J.; Leboeuf, C.; Goujon, M.; Perrais, M.; Mari, B.; Cauffiez, C.; et al. miR-92a-3p regulates cisplatin-induced cancer cell death. Cell Death Dis. 2023, 14, 603. [Google Scholar] [CrossRef] [PubMed]
- Shao, N.; Xue, L.; Wang, R.; Luo, K.; Zhi, F.; Lan, Q. miR-454-3p Is an Exosomal Biomarker and Functions as a Tumor Suppressor in Glioma. Mol. Cancer Ther. 2019, 18, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Yang, Y.; Liu, J. MicroRNA-548ac induces apoptosis in laryngeal squamous cell carcinoma cells by targeting transmembrane protein 158. Oncol. Lett. 2020, 20, 69. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Li, Y.; Lian, B.; Yin, X. microRNA-128-3p inhibits proliferation and accelerates apoptosis of gastric cancer cells via inhibition of TUFT1. World J. Surg. Oncol. 2023, 21, 47. [Google Scholar] [CrossRef]
- Sala, A.C.A.; Crespo, A.C.F.; Stelini, R.F.; Leite, O.; Moraes, A.M.; Velho, P.; Souza, E.M.; Cintra, M.L. Chronic Venous Insufficiency as a Predisposing Factor for Basal Cell Carcinoma on Legs. Ann. Vasc. Surg. 2020, 68, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Buffone, G.; Miglietta, A.M.; Abonante, S.; Giordano, V.; Renne, M.; Lugarà, M.; de Franciscis, S. Breast cancer and venous disease: A retrospective cohort study. Ann. Vasc. Surg. 2013, 27, 762–766. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Babiker, A.Y.; Anwar, S. Hesperidin, a Bioflavonoid in Cancer Therapy: A Review for a Mechanism of Action through the Modulation of Cell Signaling Pathways. Molecules 2023, 28, 5152. [Google Scholar] [CrossRef]
- Barrera-Vázquez, O.S.; Escobar-Ramírez, J.L.; Santiago-Mejía, J.; Carrasco-Ortega, O.F.; Magos-Guerrero, G.A. Discovering Potential Compounds for Venous Disease Treatment through Virtual Screening and Network Pharmacology Approach. Molecules 2023, 28, 7937. [Google Scholar] [CrossRef]
- Han, Q.F.; Liu, J.H.; Mo, D.G. The Effects of Dexamethasone on Cardiovascular Disease: Friend or Foe? Drugs Ther. 2023, 37, 847–848. [Google Scholar] [CrossRef]
- Ospina-Quintero, L.; Jaramillo, J.C.; Tabares-Guevara, J.H.; Ramírez-Pineda, J.R. Reformulating Small Molecules for Cardiovascular Disease Immune Intervention: Low-Dose Combined Vitamin D/Dexamethasone Promotes IL-10 Production and Atheroprotection in Dyslipidemic Mice. Front. Immunol. 2020, 11, 743. [Google Scholar] [CrossRef]
- Wang, H.; Yao, X.; Huang, K.; Zhang, J.; Xiao, J.; Guo, J.; Wei, D.; Xiang, B. Low-dose dexamethasone in combination with luteolin improves myocardial infarction recovery by activating the antioxidative response. Biomed. Pharmacother. 2022, 151, 113121. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhu, Q.; Chen, Y.; Ji, K.; Li, S.; Wu, Q.; Pan, Q.; Li, J. Review of the Protective Mechanism of Curcumin on Cardiovascular Disease. Drug Des. Dev. Ther. 2024, 18, 165–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhong, Y.; Wang, S.; Xu, S.; Chen, J.; Cheng, X.; Yang, X. Reversine inhibits proliferation and induces apoptosis of human osteosarcoma cells through targeting MEK1. J. Bone Oncol. 2024, 46, 100601. [Google Scholar] [CrossRef] [PubMed]
- Lips, D.J.; Bueno, O.F.; Wilkins, B.J.; Purcell, N.H.; Kaiser, R.A.; Lorenz, J.N.; Voisin, L.; Saba-El-Leil, M.K.; Meloche, S.; Pouysségur, J.; et al. MEK1-ERK2 signaling pathway protects myocardium from ischemic injury in vivo. Circulation 2004, 109, 1938–1941. [Google Scholar] [CrossRef]
- Khachigian, L.M. The MEK-ERK-Egr-1 axis and its regulation in cardiovascular disease. Vasc. Pharmacol. 2023, 153, 107232. [Google Scholar] [CrossRef]
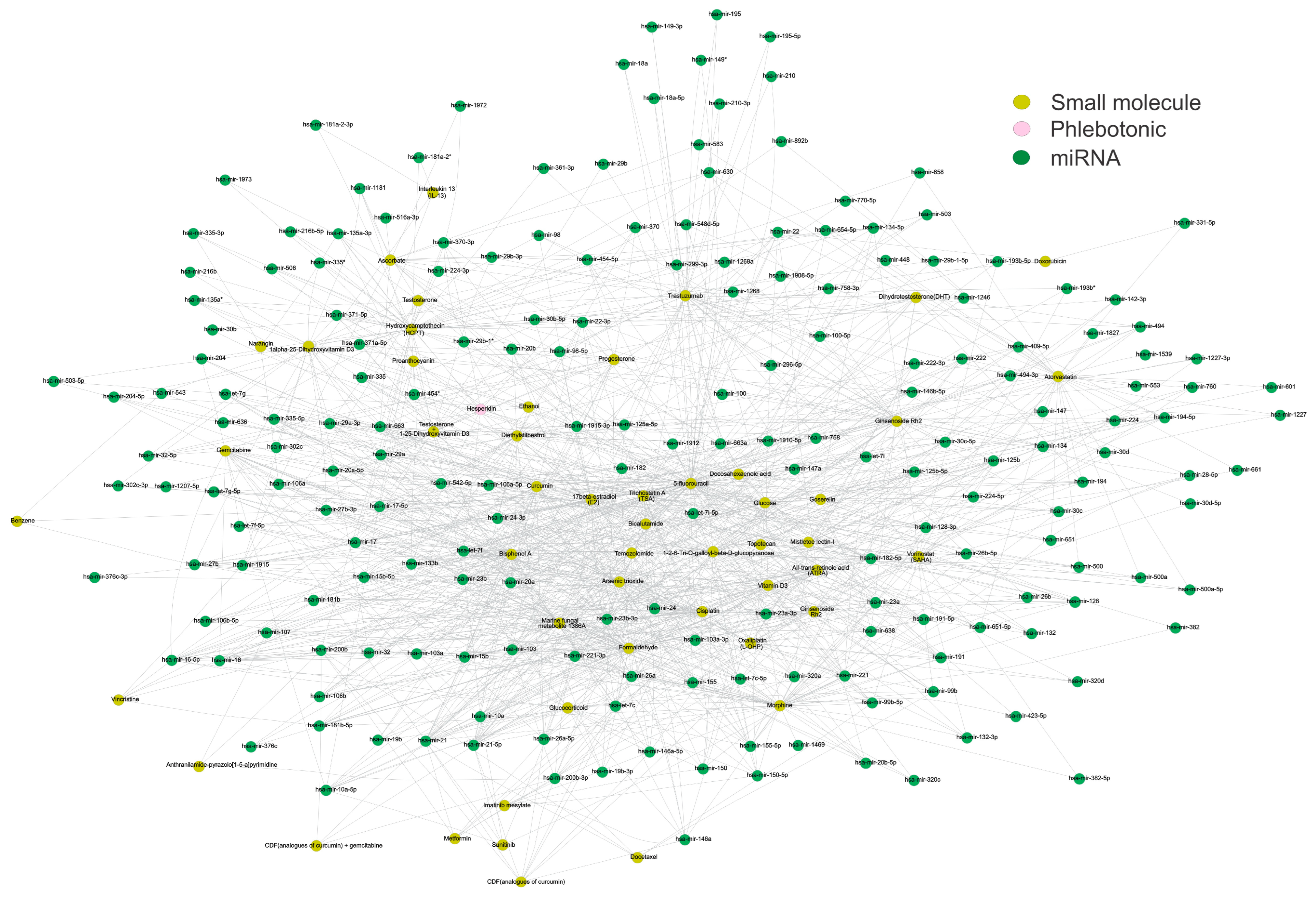
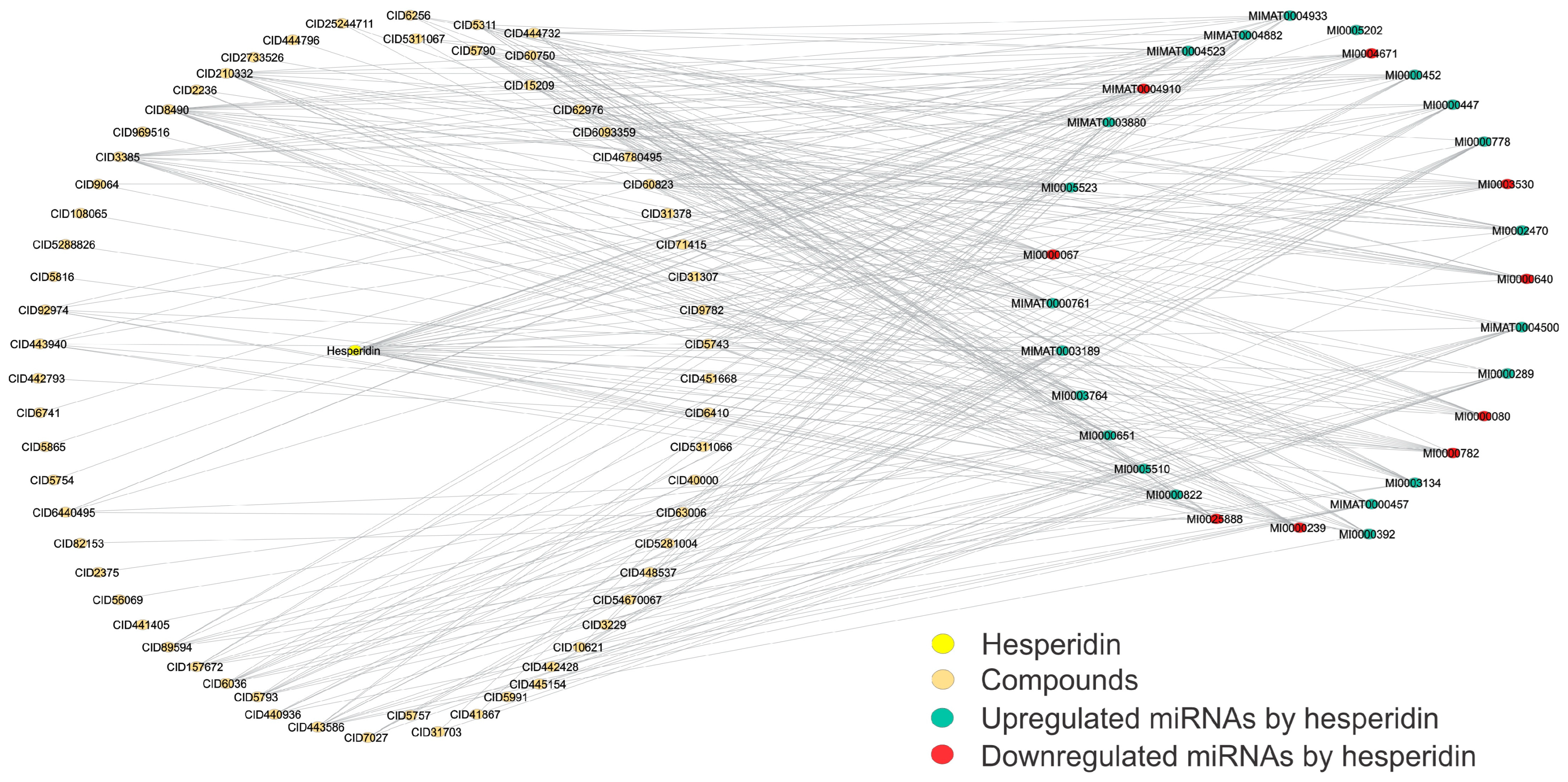

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway or Process | XD Score | q-Value | Overlap/Size |
|---|---|---|---|
| Activation of the AP1 family of transcription factors | 0.38079 | 1 | 1/10 |
| Activation of chaperones by IRE1 alpha | 0.38079 | 1 | 1/10 |
| P38MAPK events | 0.31412 | 1 | 1/12 |
| Amino acid transport across the plasma membrane | 0.29702 | 1 | 1/13 |
| Activated TAK1 mediation of p38 MAPK activation | 0.29702 | 1 | 1/13 |
| Cytochrome p450 arranged by substrate type | 0.28848 | 1 | 1/13 |
| Pyrimidine metabolism | 0.28848 | 1 | 1/13 |
| Intrinsic pathway for apoptosis | 0.2665 | 1 | 2/28 |
| Activation of BH3-only proteins | 0.24745 | 1 | 1/15 |
| Small Molecule | Pharmacological Activity | Shared miRNAs | Functions |
|---|---|---|---|
| Curcumin CID 969516  | Polyphenol has multiple signaling targets and activity at the cellular level, and it has multiple health benefits [49]. It positively affects inflammatory conditions, metabolic syndrome, and pain, and helps manage inflammatory and degenerative eye conditions and kidneys [50]. | miR-1 | It plays a crucial role in the physiological processes of smooth and skeletal muscles and other tissues, thereby contributing to the pathogenesis of a broad spectrum of disorders [51]. |
| miR-29b | This miRNA regulates PTEN gene expression by downregulating DNMT expression and hypomethylating the PTEN promoter, resulting in tumor-suppressive effects [52]. | ||
| miR-324-5p | This miRNA is implicated in cell growth, apoptosis, cancer, epilepsy, neuronal differentiation, psychiatric conditions, cardiac disease pathology, and other physiological processes [53,54,55,56,57]. | ||
| Hesperidin CID 10621  | This compound is used to treat several blood vessel disorders, such as hemorrhoids, varicose veins, and impaired circulation (venous stasis). It also has demonstrated anticancer and hepatoprotective activities [58]. | miR-374 | It involves malignant processes like digestive system carcinoma, including proliferation, apoptosis, invasion, and metastasis [59]. |
| miR-197 | It plays a crucial role in the post-transcriptional regulation of gene expression in multicellular organisms by influencing both the stability and translation of mRNAs [60]. | ||
| miR-467b | This miRNA reduces hepatocyte lipid accumulation by directly binding to the 3′UTR of Gpat1 and inhibiting GPAT1 expression at the mRNA and protein levels [61]. | ||
| miR-291b-5p | It participates during the preimplantation stage of mouse embryo development; the inhibition of autophagy is achieved by targeting Atg5 and Becn1 [62]. |
| Name | Activity | Score |
|---|---|---|
| 5-fluorouracil | This pyrimidine derivative serves as an antineoplastic agent. 5-FU demonstrates a broad-spectrum antimicrobial activity and effectively inhibits virulence factors and biofilm production [63]. | 260 |
| Glucose | The 6-carbon structure is a prevalent energy source for all organisms globally and plays a pivotal role in facilitating aerobic and anaerobic cellular respiration [64]. | 83 |
| Cisplatin | The platinum-based drug was initially approved as a cancer-fighting agent. It exerts its antitumor effects through various cytotoxic mechanisms, including DNA damage and dysfunction of cytoplasmic organelles, particularly the endoplasmic reticulum and mitochondria [65]. | 59 |
| Ginsenoside Rh2 | A compound extracted from ginseng is monomeric and possesses anticancer activities both in vitro and in vivo [66]. | 52 |
| Formaldehyde | Xenobiotic air pollutants and their universal distribution lead to widespread human exposure [67]. | 43 |
| Diethylstilbestrol | It is a synthetic estrogen compound with an estimated potency approximately five times greater than endogenous estradiol. It shares structural analogies with other xenoestrogens and functions as an estrogen receptor-alpha (ERa) agonist with an affinity comparable to estradiol [68]. | 41 |
| Enoxacin | Second-generation synthetic quinolone is recognized for its antibacterial activity. However, recent studies have focused on its potential as an anticancer agent [69]. | 35 |
| Atorvastatin | A drug commonly used to lower cholesterol by inhibiting HMG-CoA reductase [70]. | 34 |
| Arsenic trioxide | First-line treatment for acute promyelocytic leukemia is also effective for other types of leukemia [71]. | 32 |
| Vorinostat (SAHA) | This histone deacetylase inhibitor treats advanced and refractory cutaneous T-cell lymphoma [72]. | 30 |
| 17beta-estradiol (E2) | The principal circulating estrogen hormone plays a crucial role in diverse physiological functions, encompassing the development and maintenance of numerous tissues and organs [73]. | 29 |
| Hydroxycamptothecin (HCPT) | An indole alkaloid derived from the Chinese tree Camptotheca acuminate has been found to inhibit the activity of topoisomerase I effectively. This compound demonstrates a wide-ranging spectrum of anticancer activity in vitro and in vivo [74]. | 29 |
| Doxorubicin | It is a chemotherapeutic agent employed in treating a diverse array of cancers. Its mechanism of action involves targeting multiple intracellular processes: generation of reactive oxygen species, formation of DNA adducts, inhibition of topoisomerase II, eviction of histones, regulation of Ca2+ and iron hemostasis, and overproduction of ceramide [75]. | 29 |
| Emodin | It is primarily used in traditional Chinese medicine to treat carbuncles, sore throats, blood stasis, sores, and damp-heat jaundice. In recent years, pharmacological research has uncovered other potential therapeutic uses, including anticancer, neuroprotective, antidiabetic, anti-oxidant, and anti-inflammatory properties [76]. | 27 |
| Bicalutamide | It is a potent non-steroidal androgen receptor blocker known for its peripheral selectivity, attributed to poor blood–brain barrier penetration. While its primary usage is in cisgender men with prostate cancer, it has also been employed in the treatment of rare cases of peripheral precocious puberty in boys [77]. | 23 |
| Trichostatin A (TSA) | It is a natural derivative of dienohydroxamic acid derived from a fungal metabolite and exhibits various biological activities. It exerts antidiabetic activity and reverses high glucose levels by downregulating brain-derived neurotrophic factor (BDNF) expression in Schwann cells. [78]. | 23 |
| Ascorbate | It is a potent anti-oxidant and a water-soluble vitamin [79]. | 21 |
| Glucocorticoid | It is a steroid hormone commonly employed in treating autoimmune disorders, inflammation, and cancer [80]. | 20 |
| All-trans-retinoic acid (ATRA) | It is an effective agent for preventing cancer and treating skin diseases and acute promyelocytic leukemia (APL). Its pharmacological effects are mainly mediated by retinoid X receptors (RXRs) and retinoic acid receptors (RARs) [81]. | 18 |
| Mistletoe lectin-I | This lectin is the most crucial galactoside-specific lectin. Comprising a cytotoxic A-chain (29 kDa) and a distinct carbohydrate-binding B-chain (34 kDa), it exhibits immunomodulatory functions [82]. | 14 |
| Sorafenib | It represents the first-line therapy for advanced hepatocellular carcinoma. Esteemed for its robust efficacy, this drug enhances overall survival rates [83]. | 14 |
| Lenalidomide | It has been used as the standard therapy for multiple myeloma and other hematological diseases [84]. | 13 |
| Ethanol | It is a natural byproduct of fermentation and a crucial chemical in the industrial sector. Recently, it has gained recognition as a valuable biofuel [85]. | 13 |
| Docetaxel | It is a cytotoxic agent employed in the treatment of patients with breast cancer [86]. | 13 |
| Gemcitabine | It is an anticancer agent used to treat a wide range of challenging tumors, such as breast, lung, pancreatic, and ovarian cancers [87]. | 12 |
| Trastuzumab | This compound is used as first-line therapy to treat metastatic breast cancer related to HER2 overexpression [88]. | 11 |
| Catechin | This agent affects the molecular mechanisms of cancer and related disorders, including angiogenesis, extracellular matrix degradation, cell death regulation, and multidrug resistance. Many studies have shown that drinking green tea improves heart health because of its several helpful effects, including anti-inflammatory, anti-oxidant, and antithrombogenic properties [89]. | 10 |
| 1alpha-25-Dihydroxyvitamin D3 | Calcitriol, which has been identified as an active form of vitamin D3, serves as an activator of PKC and VDR [90]. | 9 |
| Progesterone | This hormone is an endogenous steroid derived from cholesterol via pregnenolone. It is a primary gonadal hormone, predominantly synthesized in the corpus luteum of the ovaries and the placenta during pregnancy [91]. | 9 |
| Name | CID | QED Index | Degree | Betweenness | Molecule Class |
|---|---|---|---|---|---|
| Desoximetasone | 5311067 | 0.787 | 3 | 0.000821 | Corticosteroid |
| Curcumin | 969516 | 0.698 | 1 | 0 | Diarylheptanoid |
| Flurandrenolide | 15209 | 0.697 | 4 | 0.002794779 | Corticosteroid |
| Trifluridine | 6256 | 0.695 | 4 | 0.003463385 | Antiviral |
| Fludrocortisone | 31378 | 0.678 | 5 | 0.002754087 | Corticosteroid |
| Diflorasone | 71415 | 0.658 | 6 | 0.003861498 | Corticosteroid |
| Reversine | 210332 | 0.513 | 15 | 0.025602386 | Purine derivative |
| Tamoxifen | 2733526 | 0.471 | 1 | 0 | Estrogen receptor modulator |
| Vorinostat | 5311 | 0.387 | 3 | 8.21 × 104 | Histone deacetylase inhibitor |
| (3R,5S)-Atorvastatin | 46780495 | 0.212 | 12 | 0.013398551 | HMG-CoA reductase inhibitor |
| Atorvastatin, (3S,5R)- | 6093359 | 0.212 | 10 | 0.007799482 | HMG-CoA reductase inhibitor |
| ent-Atorvastatin | 62976 | 0.212 | 7 | 0.002740719 | HMG-CoA reductase inhibitor |
| Atorvastatin | 60823 | 0.212 | 12 | 0.013398551 | HMG-CoA reductase inhibitor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrera-Vázquez, O.S.; Escobar-Ramírez, J.L.; Magos-Guerrero, G.A. Network Pharmacology Approaches Used to Identify Therapeutic Molecules for Chronic Venous Disease Based on Potential miRNA Biomarkers. J. Xenobiot. 2024, 14, 1519-1540. https://doi.org/10.3390/jox14040083
Barrera-Vázquez OS, Escobar-Ramírez JL, Magos-Guerrero GA. Network Pharmacology Approaches Used to Identify Therapeutic Molecules for Chronic Venous Disease Based on Potential miRNA Biomarkers. Journal of Xenobiotics. 2024; 14(4):1519-1540. https://doi.org/10.3390/jox14040083
Chicago/Turabian StyleBarrera-Vázquez, Oscar Salvador, Juan Luis Escobar-Ramírez, and Gil Alfonso Magos-Guerrero. 2024. "Network Pharmacology Approaches Used to Identify Therapeutic Molecules for Chronic Venous Disease Based on Potential miRNA Biomarkers" Journal of Xenobiotics 14, no. 4: 1519-1540. https://doi.org/10.3390/jox14040083
APA StyleBarrera-Vázquez, O. S., Escobar-Ramírez, J. L., & Magos-Guerrero, G. A. (2024). Network Pharmacology Approaches Used to Identify Therapeutic Molecules for Chronic Venous Disease Based on Potential miRNA Biomarkers. Journal of Xenobiotics, 14(4), 1519-1540. https://doi.org/10.3390/jox14040083