
Pesticide Pollution: Detrimental Outcomes and Possible Mechanisms of Fish Exposure to Common Organophosphates and Triazines




Abstract
1. Introduction
2. Spatial Distribution of Common Pesticides in Water Bodies
3. Bioaccumulation of Organophosphate and Triazine Pesticides in Fish
4. Organophosphate and Triazine Pesticides: Adverse Effects for Fish
4.1. Oxidative Stress as the Dominant Non-Cholinesterase-Related Downstream Adverse Effect of Organophosphate and Triazine Pesticides

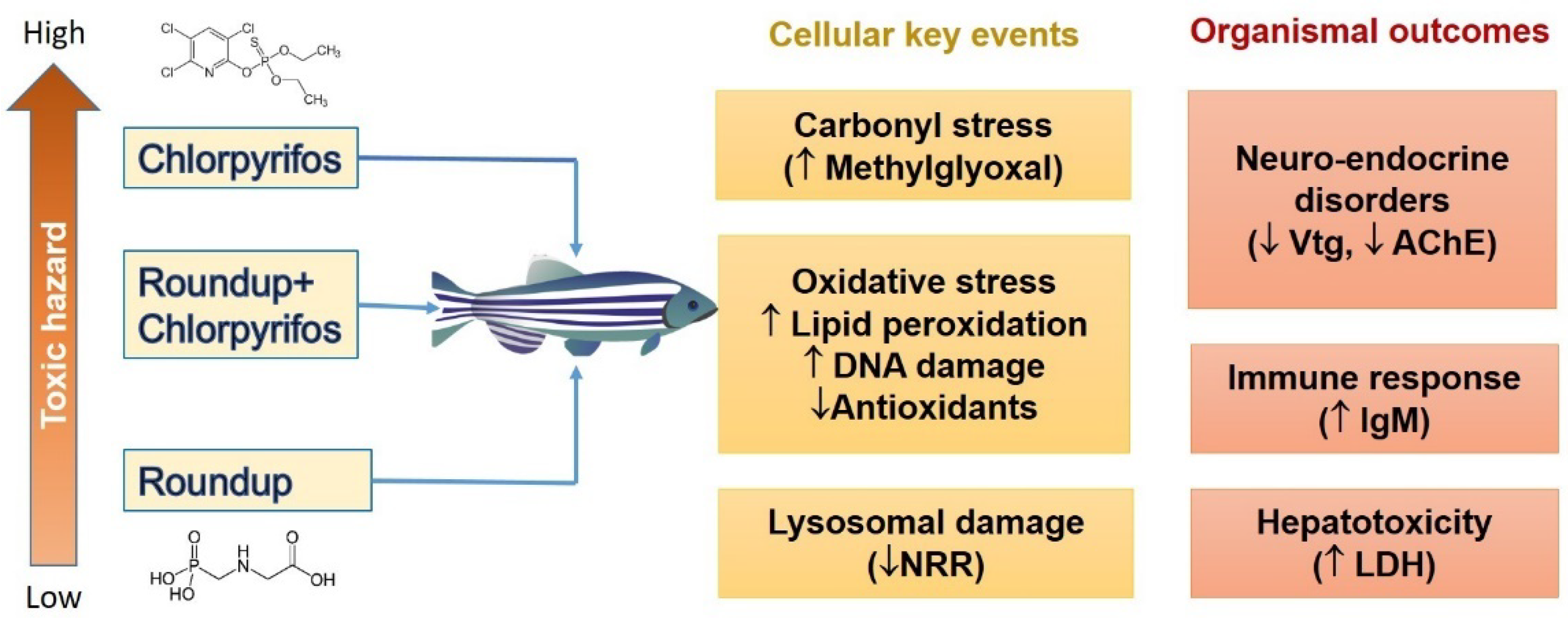{kind=link}
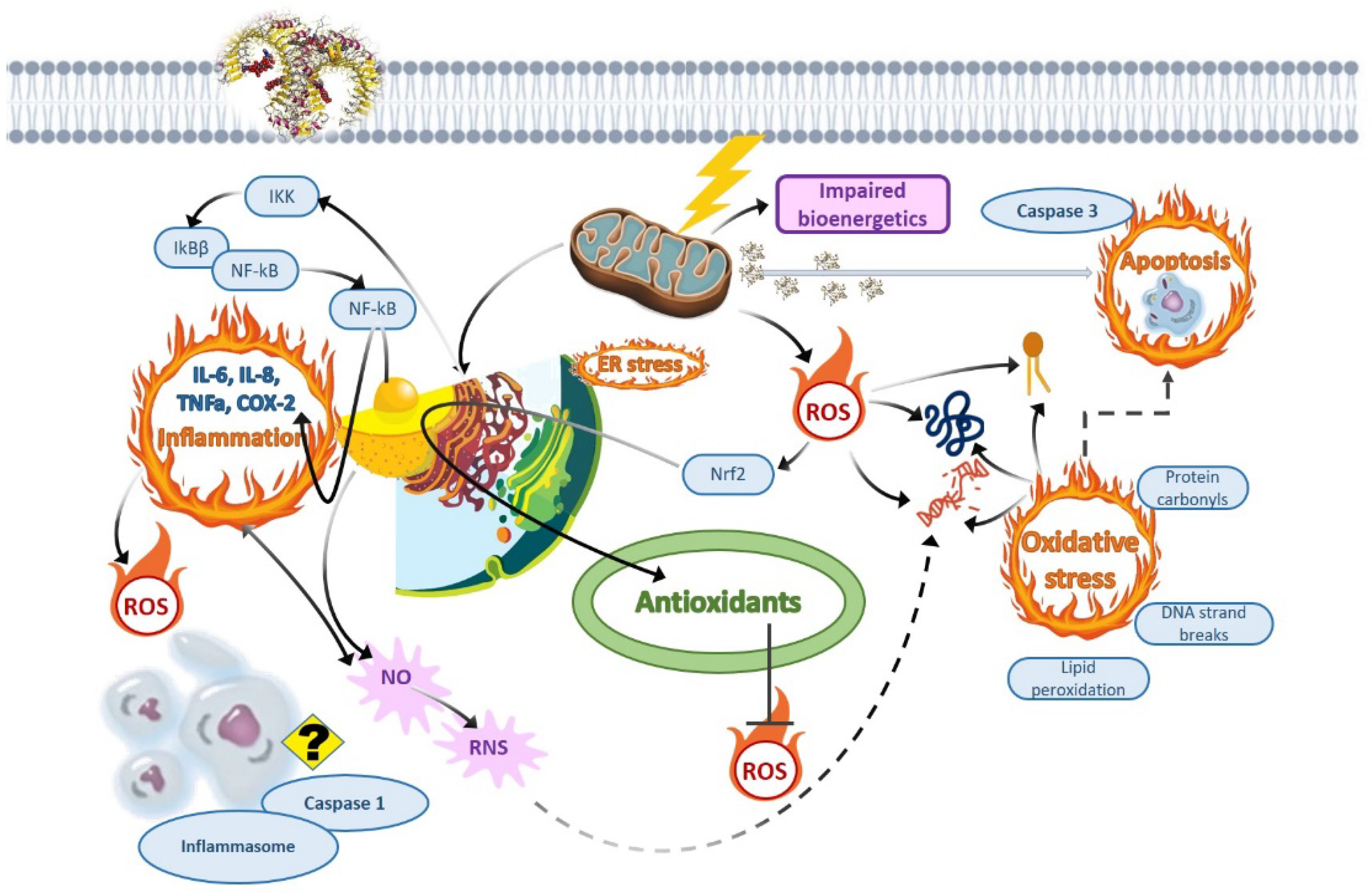{kind=link}
| Species | Pesticide, Concentration | Effects | Reference |
|---|---|---|---|
| Chlorpyrifos exposure | |||
| Common carp (Cyprinus carpio) | Colosso FC30, (0.3 μg·L−1 CYP +0.5 μg·L−1 CPF; 0.6 μg·L−1 CYP + and 1 μg·L−1 CPF | TBARS↑, NPSH↓, ASA~, GST↑, CAT↓ | [97] |
| Common carp (Cyprinus carpio) | 0.1, 0.05, 0.03 μg·L−1 CPF in combination with 0.0006, 0.0003, 0.0001 μg·L−1 CYP correspondingly, 96 h | CAT↑, GR↓, GPx↓ | [72] |
| Zebrafish (Danio rerio) | Colosso FC30, (0.3 μg·L−1 CYP +0.5 μg·L−1 CPF; 0.6 μg·L−1 CYP + and 1 μg·L−1 CPF | TBARS↑, NPSH↑, ASA↓, GST↑, CAT~ | [97] |
| Zebrafish (Danio rerio) early life stages | CPF (236 μg·L−1), CYP (5.9μg·L−1) and 236 μg·L−1 CPF + 5.9 μg·L−1 CYP, 96 h | Malformation, death in larvae, affected hatchability SOD↓, MDA↑, CAT(CPF) ↓, apoptosis ↑ | [109] |
| Zebrafish (Danio rerio) | Chlorpyrifos, 0.1 µg·L−1 and 3 µg·L−1, 14 days | Mitochondria swelling↑, Lysosomal stability↓, Cathepsin D↑, LDH↑, SDH↓, Methylglyoxal↑, ROS↑, RNS↑, TAC↓ | [115] |
| Carp, Ctenopharyngodon idellus | Chlorpyrifos, 1.4 µg·L−1 and 2.44µg·L−1 for 15, 30 and 60 days | SOD↓, MDA↑, CAT~, GSH↓, GST↓ | [98] |
| Rainbow trout, Oncorhynchus mykiss | Chlorpyrifos, 0, 2, 4 and 6 μg·L−1 7, 14, 21 days | erythrocyte count ↓, haemoglobin↓, haematocrit↓, leucocyte count↑, histological disturbances↑, AChE↓, short-term: SOD↑, CAT↑; long-term: SOD↓, CAT↓ | [116] |
| Largemouth bass (Micropterus salmoides) | 4 μg·L−1 CPF, 60 days | Apoptosis ↑, Inflammation ↑, MDA↑, SOD↑, GPx~, ACP~, CAT~, ALP~, Nrf2↑, cat~↑, sod~↑ | [117] |
| Zebrafish (Danio rerio) early life stages | 100 and 300 μg·L−1 CPF, 96 hpf | Hatchability↓, heart rate↓, morphological abnormalities↑, Immunotoxicity↑, MDA↑, CAT~, GSH↓, TNFα↑, IFN↑, IL-1β↑, IL6↑, C4↑ | [118] |
| Glyphosate/Roundup exposure | |||
| European eel (Anguilla anguilla) | 58, 116 μg·L−1 (Roundup), 1 and 3 days | DNAsb↑, erythrocytic nuclear abnormalities↑, LPO↑ CAT~, GST~, GPx~, GR~, GSHt~ | [119] |
| Spotted snakehead (Channa punctatus) | 3.25–6.51 mg·L−1 (Roundup) 1, 7, 14, 21, 28, 35 days | TBARS↑, DNA damage↑, LPO↑, ROS; CAT↓, SOD↓, GR↓ | [120] |
| Goldfish (Carassius auratus) | 2.5–20 mg·L−1 (Roundup), 2 month | CAT↑; GSH↓, GST↓, GR↓, G6PDH↓, SOD↓ | [121] |
| Goldfish (Carassius auratus) | 0.2 mmol·L−1 (Nongteshi, 30% glyphosate), 90 days | creatinine ↑, urine nitrogen↑, ALT↑, AST↑, LDH↑, MDA↑, 3- hydroxybutyrate↑; SOD↓, GPx↓, GR↓ | [122] |
| Air-breathing teleosts Anabas testudineus Heteropneustes fossilis | 17.20 mg·L−1 (Excel Mera 71) | AChE↑, LPO↑, CAT↑; GST↓, Total protein↓ | [123] |
| Zebrafih (Danio rerio) | 10, 50, 100, 200, 400 μg·L−1 (Glyphosate), 48 h | Nitric Oxide↓, Cacana1C↓, RYR2a↓, HSPb11↑ | [124] |
| Malathion exposure | |||
| Stinging catfish (Heteropneustes fossilis) | 0.44, 0.88 and 1.76 mg·L−1, 3 weeks | Erythrocytes↓, Leucocytes↓, Hemoglobin↓, Ht↓, Glucose↑, Plasma proteins↓, glutamic-oxaloacetic transaminase↑, glutamic-pyruvic transaminase↑ | [125] |
| Zebrafish (Danio rerio) | 5 and 50 µg·L−1, 14 days | Mito swelling↑, Lysosomal stability↓, Cathepsin D↑, LDH↑, SDH↓, Methylglyoxal~, ROS~, RNS~↑, TAC↓↑ | [115] |
| Spotted snakehead (Channa punctatus) | 0.4 mg·L−1, 1, 4, 8, 12 days | SOD↑, CAT↑, LPO↑, serum glucose↓, protein↑, cholesterol~, albumin~ | [126] |
| Black pacu (Colossoma macropomum) | 7.30 mg·L−1, 96 h | Mitochondrial respiration~, ROS (mito)↓, GST↑, CAT↑, SOD↑, RAS↑, LPO~, p53~ | [127] |
| Senegalese sole (Solea senegalensis) early life stages | 1.56, 3.12, and 6.25 μg·L−1 | AChE↓, BChE↓, CbE↓, pyknotic nuclei↑, CYP1A~, expression AChE~ | [128] |
| Pesticide toxic effects + dietary supplement | |||
| Nile tilapia (Oreochromis niloticus) | 15 μg·L−1 CPF, 28 days CPF + 0.5% and 1% Spirulina platensis, 28 days | CPF: ALT↑, AST↑, ALP↑, SOD↓, CAT↓, MDA↑ CPF + Spirulina: ALT↓, AST↓, ALP↓, SOD↑, CAT↑, MDA↓ | [113] |
| African sharpthooth catfish (Clarias gariepinus) | 8.75 μg·L−1 CPF, 6 weeks CPF + Carica papaya (250 mg·kg−1 bw),6 weeks | CPF: ALT↓, AST↑, Glucose↑, AChE↓, TAC↑, MDA↑, GSH↓ CPF + papaya: ALT↑, AST↓, Glucose↓, AChE↑, TAC↓, MDA↓, GSH↑ | [129] |
| African sharpthooth catfish (Clarias gariepinus) | 1.5 mg·L−1 CPF, Spirulina platensis and β-glucan (0.5%) | CPF: ALT↑, AST↑, LDL↑, Glucose↑, Triglycerides↑, MDA↑, antioxidants↓, P450↑ CPF + Spirulina: Phagocytic activity↑, P450↓, MDA↓ | [130] |
| Nile tilapia (Oreochromis niloticus) | CPF (15 mg·L−1) Chlorella vulgaris (2–3%), 4/8 weeks | CPF: HSP70↑, GPx↑, GS↑, GR↓; IL-1β↑, TNF-α↑, TGFβ1↑, IL-8↑ CPF + Chlorella: HSP70↑↑, GPx↑↑, GS↑↑, GR↓↓; IL-1β↑↑, TNF-α↑↑, TGFβ1↑↑, IL-8↑↑ | [131] |
| Mixed effects | |||
| Parachromis dovii, Poecilia gillii | CPF (5 µg·L−1) + difenoconazole (325 µg·L−1) | CYP1A ↑, EROD↑, ChE ↓ | [132] |
| Neotropical fish (Rhamdia quelen) | Imidacloprid (0.11 µg·L−1) + propoxur (0.039 µg·L−1), 96 h | AChE↓, GST~↓, CAT↓↑, SOD↑, ROS~↓, ASAP~, antagonistic effect | [133] |
| Pacu (Piaractus mesopotamicus) | endosulfan (1.1 μg·L−1) + lambda-cyhalothrin (0.7 μg·L−1), 96 h | white blood cells count↑, Lymphocytes↓, Eosinophils↑↑, AST↓, ALT↓. GST↑, GR~, GPx~, CAT↑, TBARS↑, antagonistic effect | [134] |
4.2. Pesticide-Induced Carbonyl Stress in Fish
4.3. Pesticide-Induced Nitrosative Stress in Fish
4.4. Immunomodulatory and Inflammatory Effects of Organophosphate and Triazine Pesticides
4.5. Mitochondrial and Lysosome Toxicity
4.6. Pesticide-Induced Endocrine Disorders
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AChE | acetylcholinesterase |
| ALP | alkaline phosphatase |
| ALT | alanine transaminase |
| AMPA | aminomethylphosphonic acid |
| AST | aspartate transaminase |
| BAF | bioaccumulation factor |
| BCF | bioconcentration factor |
| CAT | catalase |
| GOT | glutamic oxaloacetic transaminase |
| GPT | glutamic pyruvate transaminase |
| GPx | glutathione peroxidase |
| GST | glutathione s-transferase |
| HQ | hazard quotient |
| Kow | n-octanol–water partition coefficient |
| LDH | lactate dehydrogenase |
| LOQ | limit of quantification |
| LPO | lipid peroxidation |
| PCBs | polychlorinated biphenyls |
| MDA | malondialdehyde |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| SOD | superoxide dismutase |
| TBARS | thiobarbituric acid-reactive substance |
References
- Agarwal, A.; Prajapati, R.; Singh, O.P.; Raza, S.K.; Thakur, L.K. Pesticide residue in water: A challenging task in India. Environ. Monit. Assess. 2015, 187, 54. [Google Scholar] [CrossRef] [PubMed]
- Naughton, S.X.; Terry, A.V., Jr. Neurotoxicity in acute and repeated organophosphate exposure. Toxicology 2018, 408, 101–112. [Google Scholar] [CrossRef]
- Tang, F.H.M.; Lenzen, M.; McBratney, A.; Maggi, F. Risk of pesticide pollution at the global scale. Nat. Geosci. 2021, 14, 206–210. [Google Scholar] [CrossRef]
- Worldometer. Pesticide Use by Country. Available online: https://www.worldometers.info/food-agriculture/pesticides-by-country/ (accessed on 7 July 2022).
- FAOSTAT Analytical Brief. Pesticides use Global, Regional and Country Trends, 1990–2018. 2020. Available online: https://www.fao.org/3/cb3411en/cb3411en.pdf (accessed on 7 July 2022).
- Taylor, M.D.; Klaine, S.J.; Carvalho, F.P.; Barcelo, D. Pesticide Residues in Coastal Tropical Ecosystems: Distribution, Fate and Effects; CRC Press: Boca Raton, FL, USA, 2002; p. 576. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Matviishyn, T.M.; Husak, V.V.; Storey, J.M.; Storey, K.B. Pesticide toxicity: A mechanistic approach. EXCLI J. 2018, 17, 1101–1136. [Google Scholar] [CrossRef]
- Bodnar, O.; Horyn, O.; Soroka, O.; Nimko, K.; Falfushynska, H. Pesticide Pollution of Aquatic Ecosystems: Environmental Risks and Mechanisms of Impact on Aquatic Organisms (a Review). Hydrobiol. J. 2022, 58, 62–78. [Google Scholar] [CrossRef]
- Okwuosa, O.B.; Eyo, J.E.; Omovwohwovie, E.E. Role of Fish as Bioindicators: A Review. Iconic Res. Eng. J. 2019, 2, 354–368. [Google Scholar]
- Farag, M.R.; Alagawany, M.; Taha, H.S.A.; Ismail, T.A.; Khalil, S.R.; Abou-Zeid, S.M. Immune response and susceptibility of Nile tilapia fish to Aeromonas hydrophila infection following the exposure to Bifenthrin and/or supplementation with Petroselinum crispum essential oil. Ecotoxicol. Environ. Saf. 2021, 216, 112205. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zheng, B.; Liu, L. Biomonitoring and Bioindicators Used for River Ecosystems: Definitions, Approaches and Trends. Procedia Environ. Sci. 2010, 2, 1510–1524. [Google Scholar] [CrossRef]
- Falfushynska, H.; Gnatyshyna, L.L.; Stoliar, O.B.; Nam, Y.K. Various responses to copper and manganese exposure of Carassius auratus gibelio from two populations. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 154, 242–253. [Google Scholar] [CrossRef]
- Falfushynska, H.; Gnatyshyna, L.; Turta, O.; Stoliar, O.; Mitina, N.; Zaichenko, A.; Stoika, R. Responses of hepatic metallothioneins and apoptotic activity in Carassius auratus gibelio witness a release of cobalt and zinc from waterborne nanoscale composites. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2014, 160, 66–74. [Google Scholar] [CrossRef]
- Falfushynska, H.; Horyn, O.; Brzozowska, A.; Fedoruk, O.; Buyak, B.; Poznansky, D.; Poniedziałek, B.; Kokociński, M.; Rzymski, P. Is the presence of Central European strains of Raphidiopsis (Cylindrospermopsis) raciborskii a threat to a freshwater fish? An in vitro toxicological study in common carp cells. Aquat. Toxicol. 2019, 206, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Falfushynska, H.; Horyn, O.; Osypenko, I.; Rzymski, P.; Wejnerowski, Ł.; Dziuba, M.K.; Sokolova, I.M. Multibiomarker-based assessment of toxicity of central European strains of filamentous cyanobacteria Aphanizomenon gracile and Raphidiopsis raciborskii to zebrafish Danio rerio. Water Res. 2021, 194, 116923. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.E.; Sadler, K.C. Casting a wide net: Use of diverse model organisms to advance toxicology. Dis. Model Mech. 2020, 13, dmm.043844. [Google Scholar] [CrossRef]
- Leung, M.C.K.; Meyer, J.N. Mitochondria as a target of organophosphate and carbamate pesticides: Revisiting common mechanisms of action with new approach methodologies. Reprod. Toxicol. 2019, 89, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Santed, F.; Colomina, M.T.; Herrero Hernández, E. Organophosphate Pesticide Exposure and Neurodegeneration. Cortex 2016, 74, 417–426. [Google Scholar] [CrossRef]
- Falfushynska, H.; Khatib, I.; Kasianchuk, N.; Lushchak, O.; Horyn, O.; Sokolova, I.M. Toxic effects and mechanisms of common pesticides (Roundup and chlorpyrifos) and their mixtures in a zebrafish model (Danio rerio). Sci. Total Environ. 2022, 833, 155236. [Google Scholar] [CrossRef]
- Husak, V.; Strutynska, T.; Burdyliuk, N.; Pitukh, A.; Bubalo, V.; Falfushynska, H.; Strilbytska, O.; Lushchak, O. Low-toxic herbicides Roundup and Atrazine disturb free radical processes in Daphnia in environmentally relevant concentrations. EXCLI J. 2022, 21, 595–609. [Google Scholar] [CrossRef]
- Nabi, M.; Tabassum, N. Role of Environmental Toxicants on Neurodegenerative Disorders. Front. Toxicol. 2022, 4, 837579. [Google Scholar] [CrossRef]
- EU Pesticide Database. Active Substances, Safeners and Synergists. EU Pesticides Database (v.2.2) Search Active Substances, Safeners and Synergists. Available online: https://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/active-substances/?event=search.as (accessed on 31 August 2022).
- Ministry of Environmental Protection and Natural Resources of Ukraine. National Report about the Current State of Environment in Ukraine, 2020–2021. Available online: https://mepr.gov.ua/timeline/Nacionalni-dopovidi-pro-stan-navkolishnogo-prirodnogo-seredovishcha-v-Ukraini.html (accessed on 31 August 2022).
- Curchod, L.; Oltramare, C.; Junghans, M.; Stamm, C.; Dalvie, M.A.; Röösli, M.; Fuhrimann, S. Temporal variation of pesticide mixtures in rivers of three agricultural watersheds during a major drought in the Western Cape, South Africa. Water Res. 2019, 6, 100039. [Google Scholar] [CrossRef]
- Kruć-Fijałkowska, R.; Dragon, K.; Drożdżyński, D.; Górski, J. Seasonal variation of pesticides in surface water and drinking water wells in the annual cycle in western Poland, and potential health risk assessment. Sci. Rep. 2022, 12, 3317. [Google Scholar] [CrossRef]
- Bojakowska, I.; Gliwicz, T. Chloroorganiczne pestycydy i polichlorowane bifenyle w osadach rzek Polski. Prz. Geol. 2005, 53, 649–655. [Google Scholar]
- Bojakowska, I.; Stasiuk, M.; Gąsior, J.; Wołkowicz, W. Assessment of DDT, HCH and PCB pollution of lake sediments in Poland. Limnol. Rev. 2014, 2, 63–74. [Google Scholar] [CrossRef]
- Karlsson, A.S.; Lesch, M.; Weihermüller, L.; Thiele, B.; Disko, U.; Hofmann, D.; Vereecken, H.; Spielvogel, S. Pesticide contamination of the upper Elbe River and an adjacent floodplain area. J. Soils Sediments 2020, 20, 2067–2081. [Google Scholar] [CrossRef]
- Székács, A.; Mörtl, M.; Darvas, B. Monitoring Pesticide Residues in Surface and Ground Water in Hungary: Surveys in 1990–2015. J. Chem. 2015, 2015, 717948. [Google Scholar] [CrossRef]
- Site-Specific Sediment Quality Evaluation Guidelines. 2021. Available online: https://www.interreg-danube.eu/uploads/media/approved_project_output/0001/49/103390b9ead5d2b45ef549636361a1c066b76504.pdf (accessed on 7 July 2022).
- Rico, A.; de Oliveira, R.; Silva de Souza Nunes, G.; Rizzi, C.; Villa, S.; De Caroli Vizioli, B.; Montagner, C.C.; Waichman, A.V. Ecological risk assessment of pesticides in urban streams of the Brazilian Amazon. Chemosphere 2022, 291 Pt 1, 132821. [Google Scholar] [CrossRef]
- Panis, C.; Candiotto, L.Z.P.; Gaboardi, S.C.; Gurzenda, S.; Cruz, J.; Castro, M.; Lemos, B. Widespread pesticide contamination of drinking water and impact on cancer risk in Brazil. Environ. Int. 2022, 165, 107321. [Google Scholar] [CrossRef]
- Antić, N.; Radišić, M.; Radović, T.; Vasiljević, T.; Grujić, S.; Petković, A.; Dimkić, M.; Laušević, M. Pesticide Residues in the Danube River Basin in Serbia—A Survey during 2009–2011. Clean Soil Air Water 2015, 43, 197–204. [Google Scholar] [CrossRef]
- Agathokleous, E. European Union’s imminent ban on glyphosate: Hormesis should be considered in new chemical screening and selection. J. Res. 2022, 33, 1103–1107. [Google Scholar] [CrossRef]
- Global Glyphosate Market-Growth, Trends, Covid-19 Impact, and Forecasts (2022–2027). Available online: https://www.researchandmarkets.com/reports/4827969/global-glyphosate-market-growth-trends-covid (accessed on 7 July 2022).
- Kaszkowiak, K.; Kubacki, T.; Olejniczak, J.; Bondarenko, I. The concentration of glyphosate in the tap water in Greater Poland Region. Eur. J. Biol. Res. 2021, 11, 57–64. [Google Scholar] [CrossRef]
- Hanke, I.; Singer, H.; Hollender, J. Ultratrace-level determination of glyphosate, aminomethylphosphonic acid and glufosinate in natural waters by solid-phase extraction followed by liquid chromatography-tandem mass spectrometry: Performance tuning of derivatization, enrichment and detection. Anal. Bioanal. Chem. 2008, 391, 2265–2276. [Google Scholar] [CrossRef]
- Van Stempvoort, D.R.; Roy, J.W.; Brown, S.J.; Bickerton, G. Residues of the herbicide glyphosate in riparian groundwater in urban catchments. Chemosphere 2014, 95, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Van Stempvoort, D.R.; Spoelstra, J.; Senger, N.D.; Brown, S.J.; Post, R.; Struger, J. Glyphosate residues in rural groundwater, Nottawasaga River Watershed, Ontario, Canada. Pest. Manag. Sci. 2016, 72, 1862–1872. [Google Scholar] [CrossRef] [PubMed]
- Poiger, T.; Buerge, I.J.; Bächli, A.; Müller, M.D.; Balmer, M.E. Occurrence of the herbicide glyphosate and its metabolite AMPA in surface waters in Switzerland determined with on-line solid phase extraction LC-MS/MS. Environ. Sci. Pollut. Res. 2017, 24, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Feltracco, M.; Barbaro, E.; Morabito, E.; Zangrando, R.; Piazza, R.; Barbante, C.; Gambaro, A. Assessing glyphosate in water, marine particulate matter, and sediments in the Lagoon of Venice. Environ. Sci. Pollut. Res. Int. 2022, 29, 16383–16391. [Google Scholar] [CrossRef]
- Carles, L.; Gardon, H.; Joseph, L.; Sanchís, J.; Farré, M.; Artigas, J. Meta-analysis of glyphosate contamination in surface waters and dissipation by biofilms. Environ. Int. 2019, 124, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Medalie, L.; Baker, N.T.; Shoda, M.E.; Stone, W.W.; Meyer, M.T.; Stets, E.G.; Wilson, M. Influence of land use and region on glyphosate and aminomethylphosphonic acid in streams in the USA. Sci. Total Environ. 2020, 707, 136008. [Google Scholar] [CrossRef]
- Avila-Vazquez, M.; Difilippo, F.; Lean, B.; Maturano, E.; Etchegoyen, A. Environmental Exposure to Glyphosate and Reproductive Health Impacts in Agricultural Population of Argentina. J. Environ. Protect. 2018, 9, 241–253. [Google Scholar] [CrossRef]
- Peruzzo, P.J.; Porta, A.A.; Ronco, A.E. Levels of glyphosate in surface waters, sediments and soils associated with direct sowing soybean cultivation in north pampasic region of Argentina. Environ. Pollut. 2008, 156, 61–66. [Google Scholar] [CrossRef]
- Okada, E.; Pérez, D.; De Gerónimo, E.; Aparicio, V.; Massone, H.; Costa, J.L. Non-point source pollution of glyphosate and AMPA in a rural basin from the southeast Pampas, Argentina. Environ. Sci. Pollut. Res. Int. 2018, 25, 15120–15132. [Google Scholar] [CrossRef]
- Corrigendum to Commission Regulation (EU) 2020/1085 of 23 July 2020 Amending Annexes II and V to Regulation (EC) No 396/2005 of the European Parliament and of the Council as Regards Maximum Residue Levels for Chlorpyrifos and Chlorpyrifos-Methyl in or on Certain Products (Official Journal of the European Union L 239 of 24 July 2020). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32020R1085R%2801%29 (accessed on 7 July 2022).
- Ccanccapa, A.; Masiá, A.; Navarro-Ortega, A.; Picó, Y.; Barceló, D. Pesticides in the Ebro River basin: Occurrence and risk assessment. Environ. Pollut. 2016, 211, 414–424. [Google Scholar] [CrossRef]
- Canadian Water Quality Guidelines for the Protection of Aquatic Life. Chlorpyrifos. Chlorpyrifos 1.0_e. Available online: https://ccme.ca/en/res/chlorpyrifos-en-canadian-water-quality-guidelines-for-the-protection-of-aquatic-life.pdf (accessed on 7 July 2022).
- Climent, M.J.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S.; Pedreros, P.; Urrutia, R.; Herrero-Hernández, E. Determination of pesticides in river surface waters of central Chile using spe-gc-ms multi-residue method. J. Chil. Chem. Soc. 2018, 63, 4023–4031. [Google Scholar] [CrossRef]
- Sarker, S.; Akbor, M.A.; Nahar, A.; Hasan, M.; Islam, A.R.M.T.; Siddique, M.A.B. Level of pesticides contamination in the major river systems: A review on South Asian countries perspective. Heliyon 2021, 7, e07270. [Google Scholar] [CrossRef] [PubMed]
- Olsvik, P.A.; Larsen, A.K.; Berntssen, M.H.G.; Goksøyr, A.; Karlsen, O.A.; Yadetie, F.; Sanden, M.; Kristensen, T. Effects of Agricultural Pesticides in Aquafeeds on Wild Fish Feeding on Leftover Pellets Near Fish Farms. Front. Genet. 2019, 10, 794. [Google Scholar] [CrossRef] [PubMed]
- Gunkel, G.; Streit, B. Mechanisms of bioaccumulation of a herbicide (atrazine, s-triazine) in a freshwater mollusc (Ancylus fluviatilis müll.) and a fish (Coregonus fera jurine). Water Res. 1980, 14, 1573–1584. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, H.; Tao, M.; Yang, S.; Wang, L.; Liu, Y.; Ma, D.; He, Z. Organochlorine Pesticides in Consumer Fish and Mollusks of Liaoning Province, China: Distribution and Human Exposure Implications. Arch. Environ. Contam. Toxicol. 2010, 59, 444–453. [Google Scholar] [CrossRef]
- Noble, A. Partition coefficients (n-octanol—water) for pesticides. J. Chromatogr. A 1993, 642, 3–14. [Google Scholar] [CrossRef]
- Vighi, M.; Di Guardo, A. Predictive approaches for the evaluation of pesticide exposure. In Pesticide Risk in Groundwater; Vighi, M., Funari, E., Eds.; CRC Press: Boca Raton, FL, USA, 1995; pp. 73–85. [Google Scholar]
- Finizio, A.; Vighi, M.; Sandroni, D. Determination of n-octanol/water partition coefficient (Kow) of pesticide critical review and comparison of methods. Chemosphere 1997, 34, 131–161. [Google Scholar] [CrossRef]
- Ronka, S. Removal of triazine-based herbicides on specific polymeric sorbent: Batch studies. Pure Appl. Chem. 2016, 88, 1167–1177. [Google Scholar] [CrossRef]
- Kamrin, M.A. Pesticide Profiles Toxicity, Environmental Impact, and Fate; Lewis Publishers/CRC Press: Boca Raton, FL, USA, 1997; 704p. [Google Scholar]
- Stratton, G.W. Effects of the herbicide atrazine and its degradation products, alone and in combination, on phototrophic microorganisms. Arch. Environ. Contam. Toxicol. 1984, 13, 35–42. [Google Scholar] [CrossRef]
- Katagi, T. Bioconcentration, bioaccumulation, and metabolism of pesticides in aquatic organisms. Rev. Environ. Contam. Toxicol. 2010, 204, 1–132. [Google Scholar] [CrossRef]
- Golshani, R.; Mashinchian Moradi, A.; Mosavi Nodoshan, R.; Fatemi, S.M.; Ghavam Mostafavi, P. Organophosphorus pesticides (diazinon, malathion and azinfos methyl) accumulation in three fish species, in south coasts of the Caspian Sea, Iran. Iran. J. Fish Sci. 2020, 19, 3050–3062. [Google Scholar] [CrossRef]
- Bonansea, R.I.; Marino, D.J.G.; Bertrand, L.; Wunderlin, D.A.; Amé, M.V. Tissue-specific bioconcentration and biotransformation of cypermethrin and chlorpyrifos in a native fish (Jenynsia multidentata) exposed to these insecticides singly and in mixtures. Environ. Toxicol. Chem. 2017, 36, 1764–1774. [Google Scholar] [CrossRef]
- Sun, F.; Chen, H.S. Monitoring of pesticide chlorpyrifos residue in farmed fish: Investigation of possible sources. Chemosphere 2008, 71, 1866–1869. [Google Scholar] [CrossRef]
- Zalba, S.; Ten Hagen, T.L. Cell membrane modulation as adjuvant in cancer therapy. Cancer Treat. Rev. 2017, 52, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, F. Enhancing uptake of antibiotics into Gram-negative bacteria using nonribosome-targeting aminoglycoside-based adjuvants. Future Med. Chem. 2019, 11, 1519–1522. [Google Scholar] [CrossRef] [PubMed]
- Jarvinen, A.W.; Nordling, B.R.; Henry, M.E. Chronic toxicity of Dursban (chlorpyrifos) to the fathead minnow (Pimephales promelas) and the resultant acetylcholinesterase inhibition. Ecotoxicol. Environ. Saf. 1983, 7, 423–434. [Google Scholar] [CrossRef]
- Welling, W.; de Vries, J.W. Bioconcentration kinetics of the organophosphorus insecticide chlorpyrifos in guppies (Poecilia reticulata). Ecotoxicol. Environ. Saf. 1992, 23, 64–75. [Google Scholar] [CrossRef]
- Tsuda, T.; Kojima, M.; Harada, H.; Nakajima, A.; Aoki, S. Acute toxicity, accumulation and excretion of organophosphorous insecticides and their oxidation products in killifish. Chemosphere 1997, 35, 939–949. [Google Scholar] [CrossRef]
- Varó, I.; Serrano, R.; Pitarch, E.; Amat, F.; López, F.J.; Navarro, J.C. Toxicity and bioconcentration of chlorpyrifos in aquatic organisms: Artemia parthenogenetica (Crustacea), Gambusia affinis, and Aphanius iberus (Pisces). Bull. Environ. Contam. Toxicol. 2000, 65, 623–630. [Google Scholar] [CrossRef]
- El-Amrani, S.; Pena-Abaurrea, M.; Sanz-Landaluze, J.; Ramos, L.; Guinea, J.; Cámara, C. Bioconcentration of pesticides in zebrafish eleutheroembryos (Danio rerio). Sci. Total Environ. 2012, 425, 184–190. [Google Scholar] [CrossRef]
- Georgieva, E.; Yancheva, V.; Stoyanova, S.; Velcheva, I.; Iliev, I.; Vasileva, T.; Bivolarski, V.; Petkova, E.; László, B.; Nyeste, K.; et al. Which Is More Toxic? Evaluation of the Short-Term Toxic Effects of Chlorpyrifos and Cypermethrin on Selected Biomarkers in Common Carp (Cyprinus carpio, Linnaeus 1758). Toxics 2021, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Chidya, R.; Derbalah, A.; Abdel-Dayem, S.; Kaonga, C.; Tsuji, H.; Takeda, K.; Sakugawa, H. Contamination, dynamics, and health risk assessment of pesticides in seawater and marine samples from the Seto Inland Sea, Japan. Environ. Sci. Pollut. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Baskaran, S.; Kennedy, I.R. Ecological relative risk (EcoRR): Another approach for risk assessment of pesticides in agriculture. Agric. Ecosyst. Environ. 2002, 91, 37–57. [Google Scholar] [CrossRef]
- Environmental Transport and Fate Data Analysis for Malathion. Available online: https://www3.epa.gov/pesticides/nas/final/malathion/appendix-3-1.docx (accessed on 7 July 2022).
- Jia, Z.Q.; Zhang, Y.C.; Huang, Q.T.; Jones, A.K.; Han, Z.J.; Zhao, C.Q. Acute toxicity, bioconcentration, elimination, action mode and detoxification metabolism of broflanilide in zebrafish, Danio rerio. J. Hazard. Mater. 2020, 394, 122521. [Google Scholar] [CrossRef]
- Bailey, H.C.; Miller, J.L.; Miller, M.J.; Wiborg, L.C.; Deanovic, L.; Shed, T. Joint acute toxicity of diazinon and chlorpyrifos to Ceriodaphnia dubia. Environ. Toxicol. Chem. 1997, 16, 2304–2308. [Google Scholar] [CrossRef]
- Chen, L.; Cai, X.; Cao, M.; Liu, H.; Liang, Y.; Hu, L.; Yin, Y.; Li, Y.; Shi, J. Long-term investigation of heavy metal variations in mollusks along the Chinese Bohai Sea. Ecotoxicol. Environ. Saf. 2022, 236, 113443. [Google Scholar] [CrossRef]
- Serrano, R.; Hernández, F.; López, F.; Peña, J.B. Bioconcentration and Depuration of Chlorpyrifos in the Marine Mollusc Mytilus edulis. Arch. Environ. Contam. Toxicol. 1997, 33, 47–52. [Google Scholar] [CrossRef]
- Wu, C.F.; Chen, C.H.; Wu, C.Y.; Lin, C.S.; Su, Y.C.; Wu, C.F.; Tsai, H.P.; Fan, P.S.; Yeh, C.H.; Yang, W.C.; et al. Quinolone and Organophosphorus Insecticide Residues in Bivalves and Their Associated Risks in Taiwan. Molecules 2020, 25, 3636. [Google Scholar] [CrossRef]
- Tong, N.X.; Huong, T.T.T.; Huong, M.; Thuy, D.T. Determination of the bioaccumulation factors of organochlorine pesticides (ocps) at some species of bivalve mollusks in Soai Rap estuary—Ho Chi Minh city. Vietnam J. Mar. Sci. Technol. 2019, 18, 433–442. [Google Scholar] [CrossRef]
- Falfushynska, H.; Gnatyshyna, L.L.; Stoliar, O. In situ exposure history modulates the molecular responses to carbamate fungicide Tattoo in bivalve mollusk. Ecotoxicology 2013, 22, 433–445. [Google Scholar] [CrossRef]
- Gnatyshyna, L.; Falfushynska, H.; Stoliar, O.; Dallinger, R. Preliminary Study of Multiple Stress Response Reactions in the Pond Snail Lymnaea stagnalis Exposed to Trace Metals and a Thiocarbamate Fungicide at Environmentally Relevant Concentrations. Arch. Environ. Contam. Toxicol. 2020, 79, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Kalyabina, V.P.; Esimbekova, E.N.; Kopylova, K.V.; Kratasyuk, V.A. Pesticides: Formulants, distribution pathways and effects on human health—A review. Toxicol. Rep. 2021, 8, 1179–1192. [Google Scholar] [CrossRef] [PubMed]
- Eddleston, M. Novel Clinical Toxicology and Pharmacology of Organophosphorus Insecticide Self-Poisoning. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 341–360. [Google Scholar] [CrossRef]
- Marsillach, J.; Costa, L.G.; Furlong, C.E. Protein adducts as biomarkers of exposure to organophosphorus compounds. Toxicology 2013, 307, 46–54. [Google Scholar] [CrossRef]
- Mangas, I.; Estevez, J.; Vilanova, E.; França, T.C. New insights on molecular interactions of organophosphorus pesticides with esterases. Toxicology 2017, 376, 30–43. [Google Scholar] [CrossRef]
- Strilbyska, O.M.; Tsiumpala, S.A.; Kozachyshyn, I.I.; Strutynska, T.; Burdyliuk, N.; Lushchak, V.I.; Lushchak, O. The effects of low-toxic herbicide Roundup and glyphosate on mitochondria. EXCLI J. 2022, 21, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Rocha, T.L.; Santos, A.P.; Yamada, Á.T.; Soares, C.M.; Borges, C.L.; Bailão, A.M.; Sabóia-Morais, S.M. Proteomic and histopathological response in the gills of Poecilia reticulata exposed to glyphosate-based herbicide. Environ. Toxicol. Pharmacol. 2015, 40, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Nur, G.; Deveci, H.A. Histopathological and biochemical responses to the oxidative stress induced by glyphosate-based herbicides in the rainbow trout (Oncorhynchus mykiss). J. Cell Neurosci. Oxid. Stress. 2018, 10, 656–665. [Google Scholar] [CrossRef]
- Želježić, D.; Žunec, S.; Bjeliš, M.; Benković, V.; Mladinić, M.; Lovaković Tariba, B.; Pavičić, I.; Marjanović Čermak, A.M.; Kašuba, V.; Milić, M.; et al. Effects of the chloro-s-triazine herbicide terbuthylazine on DNA integrity in human and mouse cells. Environ. Sci. Pollut. Res. Int. 2018, 25, 19065–19081. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annual Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Abdollahi, M.; Ranjbar, A.; Shadnia, S.; Nikfar, S.; Rezaie, A. Pesticides and oxidative stress: A review. Med. Sci. Monit. 2004, 10, 141–147. [Google Scholar]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G. Oxidative stress and lipid peroxidation products in cancer progression and therapy. ISRN Oncol. 2012, 2012, 137289. [Google Scholar] [CrossRef] [PubMed]
- Yan, E.B.; Unthank, J.K.; Castillo-Melendez, M.; Miller, S.L.; Langford, S.J.; Walker, D.W. Novel method for in vivo hydroxyl radical measurement by microdialysis in fetal sheep brain in utero. J. Appl. Physiol. 2005, 98, 2304–2310. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nunes, M.E.M.; Müller, T.E.; Murussi, C.; do Amaral, A.M.B.; Gomes, J.L.C.; Marins, A.T.; Leitemperger, J.; Rodrigues, C.C.R.; Fiuza, T.L.; Costa, M.D.; et al. Oxidative effects of the acute exposure to a pesticide mixture of cypermethrin and chlorpyrifos on carp and zebrafish—A comparative study. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 206–207, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Jindal, R. Oxidative stress response in liver, kidney and gills of ctenopharyngodon idellus(cuvier & valenciennes) exposed to chlorpyrifos. MOJ Biol. Med. 2017, 1, 103–112. [Google Scholar] [CrossRef]
- Destro, A.L.F.; Silva, S.B.; Gregório, K.P.; de Oliveira, J.M.; Lozi, A.A.; Zuanon, J.A.S.; Salaro, A.L.; da Matta, S.L.P.; Gonçalves, R.V.; Freitas, M.B. Effects of subchronic exposure to environmentally relevant concentrations of the herbicide atrazine in the Neotropical fish Astyanax altiparanae. Ecotoxicol. Environ. Saf. 2021, 208, 111601. [Google Scholar] [CrossRef]
- Modesto, K.A.; Martinez, C.B. Roundup causes oxidative stress in liver and inhibits acetylcholinesterase in muscle and brain of the fish Prochilodus lineatus. Chemosphere 2010, 78, 294–299. [Google Scholar] [CrossRef]
- Nwani, C.D.; Lakra, W.S.; Nagpure, N.S.; Kumar, R.; Kushwaha, B.; Srivastava, S.K. Toxicity of the herbicide atrazine: Effects on lipid peroxidation and activities of antioxidant enzymes in the freshwater fish Channa punctatus (Bloch). Int. J. Environ. Res. Public Health. 2010, 7, 3298–3312. [Google Scholar] [CrossRef]
- Yang, C.; Lim, W.; Song, G. Mediation of oxidative stress toxicity induced by pyrethroid pesticides in fish. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 234, 108758. [Google Scholar] [CrossRef]
- Abhijith, B.D.; Ramesh, M.; Poopal, R.K. Responses of metabolic and antioxidant enzymatic activities in gill, liver and plasma of Catla catla during methyl parathion exposure. J. Basic Appl. Zool. 2016, 77, 31–40. [Google Scholar] [CrossRef]
- Sandoval-Herrera, N.; Mena, F.; Espinoza, M.; Romero, A. Neurotoxicity of organophosphate pesticides could reduce the ability of fish to escape predation under low doses of exposure. Sci. Rep. 2019, 9, 10530. [Google Scholar] [CrossRef] [PubMed]
- Velisek, J.; Stara, A.; Zuskova, E. Effect of single and combination of three triazine metabolites at environmental concentrations on early life stages of common carp (Cyprinus carpio L.). Environ. Sci. Pollut. Res. Int. 2016, 23, 24289–24297. [Google Scholar] [CrossRef]
- Guilherme, S.; Gaivão, I.; Santos, M.A.; Pacheco, M. DNA damage in fish (Anguilla anguilla) exposed to a glyphosate-based herbicide—Elucidation of organ-specificity and the role of oxidative stress. Mutat. Res. 2012, 743, 1–9. [Google Scholar] [CrossRef]
- D’Costa, A.H.; Shyama, S.K.; Praveen Kumar, M.K.; Fernandes, T.M. Induction of DNA damage in the peripheral blood of zebrafish (Danio rerio) by an agricultural organophosphate pesticide, monocrotophos. Int. Aquat. Res. 2018, 10, 243–251. [Google Scholar] [CrossRef]
- Ledda, C.; Cannizzaro, E.; Cinà, D.; Filetti, V.; Vitale, E.; Paravizzini, G.; Di Naso, C.; Iavicoli, I.; Rapisarda, V. Oxidative stress and DNA damage in agricultural workers after exposure to pesticides. J. Occup. Med. Toxicol. 2021, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, L.; Ren, L.; Feng, W.; Lv, P.; Wu, W.; Yan, Y. The single and joint toxicity effects of chlorpyrifos and beta-cypermethrin in zebrafish (Danio rerio) early life stages. J. Hazard. Mater. 2017, 334, 121–131. [Google Scholar] [CrossRef]
- Maheshwari, N.; Mahmood, R. Protective effect of catechin on pentachlorophenol-induced cytotoxicity and genotoxicity in isolated human blood cells. Environ. Sci. Pollut. Res. Int. 2020, 27, 13826–13843. [Google Scholar] [CrossRef]
- Ali, M.F.; Soliman, A.A.; Gewaily, M.S.; Abdel-Kader, T.Y.; Amer, A.A.; Zaineldin, A.I.; Al-Asgah, N.A.; Younis, E.M.; Abdel-Warith, A.A.; Sewilam, H.; et al. Isatis phytogenic relieved atrazine induced growth retardation, hepato-renal dysfunction, and oxidative stress in Nile tilapia. Saudi J. Biol. Sci. 2022, 29, 190–196. [Google Scholar] [CrossRef]
- Hassan, M.A.; Hozien, S.T.; Abdel Wahab, M.M.; Hassan, A. Ameliorative effect of selenium yeast supplementation on the physio-pathological impacts ofchronic exposure to glyphosate and or malathion in Oreochromis niloticus. BMC Vet. Res. 2022, 18, 159. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Dawood, M.A.O.; Elbadawy, M.; Aleya, L.; Alkahtani, S. Spirulina platensis Reduced Oxidative Damage Induced by Chlorpyrifos Toxicity in Nile Tilapia (Oreochromis niloticus). Animals 2020, 10, 473. [Google Scholar] [CrossRef] [PubMed]
- Intayoung, U.; Wunnapuk, K.; Kohsuwan, K.; Sapbamrer, R.; Khacha-Ananda, S. Effect of Occupational Exposure to Herbicides on Oxidative Stress in Sprayers. Saf. Health Work 2021, 12, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, O.; Horyn, O.; Khatib, I.; Falfushynska, H. Multibiomarker assessment in zebrafish Danio rerio after the effects of malathion and chlorpyrifos. Toxicol. Environ. Health Sci. 2021, 13, 165–174. [Google Scholar] [CrossRef]
- Ali, M.; Majid, M.; Hussain, I.; Kali, S.; Naz, T.; Niazi, M.B.K.; Khan, M.R.A.; Zafar, M.I. Chlorpyrifos mediated oxidative damage and histopathological alterations in freshwater fish Oncorhynchus mykiss in Northern Pakistan. Aquacult. Res. 2020, 51, 4583–4594. [Google Scholar] [CrossRef]
- Zhao, L.; Tang, G.; Xiong, C.; Han, S.; Yang, C.; He, K.; Liu, Q.; Luo, J.; Luo, W.; Wang, Y.; et al. Chronic chlorpyrifos exposure induces oxidative stress, apoptosis and immune dysfunction in largemouth bass (Micropterus salmoides). Environ. Pollut. 2021, 282, 117010. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, Z.; Peng, T.; Fu, Z. The toxicity of chlorpyrifos on the early life stage of zebrafish: A survey on the endpoints at development, locomotor behavior, oxidative stress and immunotoxicity. Fish Shellfish Immunol. 2015, 43, 405–414. [Google Scholar] [CrossRef]
- Guilherme, S.; Gaivao, I.; Santos, M.A.; Pacheco, M. European eel (Anguilla anguilla) genotoxic and prooxidant responses following short-term exposure to Roundup®—A glyphosate-based herbicide. Mutagenesis 2010, 25, 523–530. [Google Scholar] [CrossRef]
- Nwani, C.D.; Nagpure, N.S.; Kumar, R.; Kushwaha, B.; Lakra, W.S. DNA damage and oxidative stress modulatory effects of glyphosate-based herbicide in freshwater fish, Channa punctatus. Environ. Toxicol. Pharmacol. 2013, 36, 539–547. [Google Scholar] [CrossRef]
- Lushchak, O.V.; Kubrak, O.I.; Storey, J.M.; Storey, K.B.; Lushchak, V.I. Low toxic herbicide Roundup induces mild oxidative stress in goldfish tissues. Chemosphere 2009, 76, 932–937. [Google Scholar] [CrossRef]
- Li, M.H.; Ruan, L.Y.; Zhou, J.W.; Fu, Y.H.; Jinag, L.; Zhao, H.; Wang, J.S. Metabolic profiling of goldfish (Carassius auratis) after long-term glyphosate-base herbicide exposure. Aquat. Toxicol. 2017, 188, 159–169. [Google Scholar] [CrossRef]
- Samanta, P.; Pal, S.; Mukherjee, A.K.; Glosh, A.R. Biochemical effects of glyphosate based herbicide, Excel Mera 71 on enzyme activities of acetylcholinesterase (AChE), lipid peroxidation (LPO), catalase (CAT), gluthation-S-transferase (GST) and protein content on teleostean fishes. Ecotoxicol. Environ. Saf. 2014, 107, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Gaur, H.; Bhargava, A. Glyphosate induces toxicity and modulates calcium and NO signaling in zebrafish embryos. Biochem. Bioph. Res. Commun. 2019, 513, 1070–1075. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.; Alkahem Al-Balawi, H.F.; Al-Ghanim, K.A.; Al-Misned, F.; Mahboob, S. Risk assessment of malathion on health indicators of catfish: Food and water security prospective research. J. King Saud. Univ. Sci. 2021, 33, 101294. [Google Scholar] [CrossRef]
- Bharti, S.; Rasool, F. Analysis of the biochemical and histopathological impact of a mild dose of commercial malathion on Channa punctatus (Bloch) fish. Toxicol. Rep. 2021, 8, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Silva de Souza, S.; Machado, R.N.; Custódio da Costa, J.; Campos, D.F.; Sebrenski da Silva, G.; Fonseca de Almeida-Val, V.M. Severe damages caused by Malathion exposure in Colossoma macropomum. Ecotoxicol. Environ. Saf. 2020, 205, 111340. [Google Scholar] [CrossRef]
- Ortiz-Delgado, J.B.; Funes, V.; Albendín, G.; Scala, E.; Sarasquete, C. Toxicity of malathion during Senegalese sole, Solea senegalensis larval development and metamorphosis: Histopathological disorders and effects on type B esterases and CYP1A enzymatic systems. Environ. Toxicol. 2021, 36, 1894–1910. [Google Scholar] [CrossRef]
- Mansour, A.T.; Hamed, H.S.; El-Beltagi, H.S.; Mohamed, W.F. Modulatory Effect of Papaya Extract against Chlorpyrifos-Induced Oxidative Stress, Immune Suppression, Endocrine Disruption, and DNA Damage in Female Clarias gariepinus. Int. J. Environ. Res. Public Health. 2022, 19, 4640. [Google Scholar] [CrossRef]
- Mokhbatly, A.A.A.; Assar, D.H.; Ghazy, E.W.; Elbialy, Z.; Rizk, S.A.; Omar, A.A.; Gaafar, A.Y.; Dawood, M.A.O. The protective role of spirulina and β-glucan in African catfish (Clarias gariepinus) against chronic toxicity of chlorpyrifos: Hemato-biochemistry, histopathology, and oxidative stress traits. Environ. Sci. Pollut. Res. 2020, 27, 31636–31651. [Google Scholar] [CrossRef]
- Zahran, E.; Elbahnaswy, S.; Risha, E.; El-Matbouli, M. Antioxidative and immunoprotective potential of Chlorella vulgaris dietary supplementation against chlorpyrifos-induced toxicity in Nile tilapia. Fish Physiol. Biochem. 2020, 46, 1549–1560. [Google Scholar] [CrossRef]
- Redondo-López, S.; León, A.C.; Jiménez, K.; Solano, K.; Blanco-Peña, K.; Mena, F. Transient exposure to sublethal concentrations of a pesticide mixture (chlorpyrifos-difenoconazole) caused different responses in fish species from different trophic levels of the same community. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 251, 109208. [Google Scholar] [CrossRef]
- Marins, A.T.; Cerezer, C.; Leitemperger, J.W.; Severo, E.S.; Costa, M.D.; Fontoura, D.O.; Nunes, M.E.M.; Ribeiro, L.C.; Zanella, R.; Loro, V.L. A mixture of pesticides at environmental concentrations induces oxidative stress and cholinergic effects in the neotropical fish Rhamdia quelen. Ecotoxicology 2021, 30, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, C.; Rossi, A.; Ale, A.; Campana, M.; Parma, M.J.; Cazenave, J. Combined toxicological effects of pesticides: A fish multi-biomarker approach. Ecol. Indic. 2014, 36, 532–538. [Google Scholar] [CrossRef]
- Dhama, K.; Latheef, S.K.; Dadar, M.; Samad, H.A.; Munjal, A.; Khandia, R.; Karthik, K.; Tiwari, R.; Yatoo, M.I.; Bhatt, P.; et al. Biomarkers in Stress Related Diseases/Disorders: Diagnostic, Prognostic, and Therapeutic Values. Front. Mol. Biosci. 2019, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Di Nica, V.; González, A.B.M.; Lencioni, V.; Villa, S. Behavioural and biochemical alterations by chlorpyrifos in aquatic insects: An emerging environmental concern for pristine Alpine habitats. Environ. Sci. Pollut. Res. Int. 2020, 27, 30918–30926. [Google Scholar] [CrossRef] [PubMed]
- Oğuz, A.R.; Kaval Oğuz, E.; Özok, N. Effects of chlorpyrifos on primary gill cell culture of Lake Van fish (Alburnus tarichi Güldenstaadt 1814). Toxicol. Res. 2020, 9, 741–745. [Google Scholar] [CrossRef]
- Parvez, S.; Raisuddin, S. Protein carbonyls: Novel biomarkers of exposure to oxidative stress-inducing pesticides in freshwater fish Channa punctata (Bloch). Environ. Toxicol. Pharmacol. 2005, 20, 112–117. [Google Scholar] [CrossRef]
- Berkoz, M.; Ozkan-Yilmaz, F.; Ozluer-Hunt, A.; Gunduz, S.G.; Yİldİrİm, M.; Yalİn, S. Influence of sublethal chlorpyrifos exposure on oxidative stress and acetylcholinesterase activity in common carp (Cyprinus carpio). Fresenius Environ. Bull. 2019, 28, 4642–4649. [Google Scholar]
- Shah, Z.U.; Parveen, S. Oxidative, biochemical and histopathological alterations in fishes from pesticide contaminated river Ganga, India. Sci. Rep. 2022, 12, 3628. [Google Scholar] [CrossRef] [PubMed]
- Bjørling-Poulsen, M.; Andersen, H.R.; Grandjean, P. Potential developmental neurotoxicity of pesticides used in Europe. Environ. Health. 2008, 7, 50. [Google Scholar] [CrossRef]
- Martínez, M.C.; Andriantsitohaina, R. Reactive nitrogen species: Molecular mechanisms and potential significance in health and disease. Antioxid. Redox Signal. 2009, 11, 669–702. [Google Scholar] [CrossRef]
- Wink, D.A.; Mitchell, J.B. Chemical biology of nitric oxide: Insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free Radic. Biol. Med. 1998, 25, 434–456. [Google Scholar] [CrossRef]
- Eddy, F.B. Role of nitric oxide in larval and juvenile fish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2005, 142, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Ward, M.; Grabowski, P.; Secombes, C.J. Molecular cloning, gene organization and expression of rainbow trout (Oncorhynchus mykiss) inducible nitric oxide synthase (iNOS) gene. Biochem. J. 2001, 358, 747–755. [Google Scholar] [CrossRef]
- Ko, E.Y.; Cho, S.H.; Kwon, S.H.; Eom, C.Y.; Jeong, M.S.; Lee, W.; Kim, S.Y.; Heo, S.J.; Ahn, G.; Lee, K.P.; et al. The roles of NF-κB and ROS in regulation of pro-inflammatory mediators of inflammation induction in LPS-stimulated zebrafish embryos. Fish Shellfish Immunol. 2017, 68, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.K.; Chandra, R.K.; Tripathi, M.K. Analysis of suppressive effects of pesticide triazophos on leucocyte immune responses in a teleost, Channa Punctatus. Drug Chem. Toxicol. 2022, 45, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Liu, T.; Wang, C.; Zhao, F.Q.; Zhang, Z.W.; Yao, H.D.; Xing, H.J.; Xu, S.W. Effects of atrazine and chlorpyrifos on the production of nitric oxide and expression of inducible nitric oxide synthase in the brain of common carp (Cyprinus carpio L.). Ecotoxicol. Environ. Saf. 2013, 93, 7–12. [Google Scholar] [CrossRef]
- Jin, Y.; Chen, R.; Liu, W.; Fu, Z. Effect of endocrine disrupting chemicals on the transcription of genes related to the innate immune system in the early developmental stage of zebrafish (Danio rerio). Fish Shellfish Immunol. 2010, 28, 854–861. [Google Scholar] [CrossRef]
- Liu, J.; Gupta, R.C.; Goad, J.T.; Karanth, S.; Pope, C. Modulation of parathion toxicity by glucose feeding: Is nitric oxide involved? Toxicol. Appl. Pharmacol. 2007, 219, 106–113. [Google Scholar] [CrossRef]
- Chan, J.Y.; Wu, C.H.; Tsai, C.Y.; Cheng, H.L.; Dai, K.Y.; Chan, S.H.; Chang, A.Y. Transcriptional up-regulation of nitric oxide synthase II by nuclear factor-kappaB at rostral ventrolateral medulla in a rat mevinphos intoxication model of brain stem death. J. Physiol. 2007, 581, 1293–1307. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Mehrpour, O.; Buhrmann, C.; Pourbagher-Shahri, A.M.; Shakibaei, M.; Samarghandian, S. Organophosphorus Compounds and MAPK Signaling Pathways. Int. J. Mol. Sci. 2020, 21, 4258. [Google Scholar] [CrossRef]
- Parny, M.; Coste, A.; Aubouy, A.; Rahabi, M.; Prat, M.; Pipy, B.; Treilhou, M. Differential immunomodulatory effects of six pesticides of different chemical classes on human monocyte-derived macrophage functions. Food Chem. Toxicol. 2022, 163, 112992. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yin, D.; Hu, S.; Hou, Y. Immunotoxicity of pentachlorophenol on macrophage immunity and IgM secretion of the crucian carp (Carassius auratus). Bull. Environ. Contam. Toxicol. 2004, 73, 153–160. [Google Scholar] [CrossRef]
- Díaz-Resendiz, K.J.G.; Girón-Pérez, M.I. Effect of chlorpyrifos on the immune response of Nile tilapia (Oreochromis niloticus). Rev. Biol. Cienc. 2014, 3, 59–64. [Google Scholar]
- Mashoof, S.; Criscitiello, M.F. Fish Immunoglobulins. Biology 2016, 5, 45. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Resendiz, K.J.; Toledo-Ibarra, G.A.; Girón-Pérez, M.I. Modulation of immune response by organophosphorus pesticides: Fishes as a potential model in immunotoxicology. J. Immunol. Res. 2015, 2015, 213836. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wang, Q.; Huang, Z.; Ding, L.; Xu, Z. Immunoglobulins, Mucosal Immunity and Vaccination in Teleost Fish. Front. Immunol. 2020, 11, 567941. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, L.; Zhang, Y.; Fang, Q.; Li, Y. Toxic effects of chlorpyrifos on lysozyme activities, the contents of complement C3 and IgM, and IgM and complement C3 expressions in common carp (Cyprinus carpio L.). Chemosphere 2013, 93, 428–433. [Google Scholar] [CrossRef]
- Yang, C.; Lim, W.; Song, G. Immunotoxicological effects of insecticides in exposed fishes. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 247, 109064. [Google Scholar] [CrossRef]
- Giron-Perez, M.I.; Santerre, A.; Gonzalez-Jaime, F.; Casas-Solis, J.; Hernandez-Coronado, M.; Peregrina-Sandoval, J.; Takemura, A.; Zaitseva, G. Immunotoxicity and hepatic function evaluation in Nile tilapia (Oreochromis niloticus) exposed to diazinon. Fish Shellfish Immunol. 2007, 23, 760–769. [Google Scholar] [CrossRef]
- Moustafa, G.G.; Shaaban, F.E.; Hadeed, A.H.; Elhady, W.M. Immunotoxicological, biochemical, and histopathological studies on Roundup and Stomp herbicides in Nile catfish (Clarias gariepinus). Vet. World 2016, 9, 638–647. [Google Scholar] [CrossRef]
- Kirsten, K.S.; Canova, R.; Soveral, L.F.; Friedrich, M.T.; Frandoloso, R.; Kreutz, L.C. Reduced expression of selective immune-related genes in silver catfish (Rhamdia quelen) monocytes exposed to atrazine. Fish Shellfish Immunol. 2017, 64, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Pérez, M.R.; Covantes-Rosales, C.E.; Toledo-Ibarra, G.A.; Mercado-Salgado, U.; Ponce-Regalado, M.D.; Díaz-Resendiz, K.J.G.; Girón-Pérez, M.I. Organophosphorus Pesticides as Modulating Substances of Inflammation through the Cholinergic Pathway. Int. J. Mol. Sci. 2022, 23, 4523. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, C.; Cabon, J.; Louboutin, L.; Le Floch, S.; Morin, T.; Danion, M. Cellular, humoral and molecular responses in rainbow trout (Oncorhynchus mykiss) exposed to a herbicide and subsequently infected with infectious hematopoietic necrosis virus. Aquat. Toxicol. 2019, 215, 105282. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Rapisarda, V.; Catania, S.; Di Nola, C.; Ledda, C.; Fenga, C. Cytokine patterns in greenhouse workers occupationally exposed to α-cypermethrin: An observational study. Environ. Toxicol. Pharmacol. 2013, 36, 796–800. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, X.; Gui, B.; Xu, X.; Su, L.; Zhao, Y.H.; Martyniuk, C.J. Comparison of modes of action between fish, cell and mitochondrial toxicity based on toxicity correlation, excess toxicity and QSAR for class-based compounds. Toxicology 2022, 470, 153155. [Google Scholar] [CrossRef] [PubMed]
- Sagarkar, S.; Gandhi, D.; Devi, S.S.; Sakharkar, A.; Kapley, A. Atrazine exposure causes mitochondrial toxicity in liver and muscle cell lines. Indian J. Pharmacol. 2016, 48, 200–207. [Google Scholar] [CrossRef]
- Azevedo, R.D.; Falcão, K.V.; Assis, C.R.; Martins, R.M.; Araújo, M.C.; Yogui, G.T.; Neves, J.L.; Seabra, G.M.; Maia, M.B.; Amaral, I.P.; et al. Effects of pyriproxyfen on zebrafish brain mitochondria and acetylcholinesterase. Chemosphere 2021, 263, 128029. [Google Scholar] [CrossRef]
- Nassar, A.M.K.; Abdel-Halim, K.Y.; Abbassy, M.A. Mitochondrial biochemical and histopathological defects induced by the herbicide pendimethalin in tilapia fish (Oreochromis niloticus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 242, 108949. [Google Scholar] [CrossRef]
- Das, B.K.; Mukherjee, S.C. Chronic toxic effects of quinalphos on some biochemical parameters in Labeo rohita (Ham.). Toxicol. Lett. 2000, 114, 11–18. [Google Scholar] [CrossRef]
- Wang, L.; Espinoza, H.M.; MacDonald, J.W.; Bammler, T.K.; Williams, C.R.; Yeh, A.; Louie, K.W.; Marcinek, D.J.; Gallagher, E.P. Olfactory transcriptional analysis of Salmon exposed to mixtures of chlorpyrifos and malathion reveal novel molecular pathways of neurobehavioral injury. Toxicol. Sci. 2016, 149, 145–157. [Google Scholar] [CrossRef]
- Deus, C.M.; Yambire, K.F.; Oliveira, P.J.; Raimundo, N. Mitochondria-Lysosome Crosstalk: From Physiology to Neurodegeneration. Trends Mol. Med. 2020, 26, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.P.; Hewitt, E.W. Cellular proteostasis: Degradation of misfolded proteins by lysosomes. Essays Biochem. 2016, 60, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yang, X.; Chen, L.; Liu, Y.Y.; Venkatarangan, V.; Reist, L.; Hanson, P.; Xu, H.; Wang, Y.; Li, M. A conserved ubiquitin- and ESCRT-dependent pathway internalizes human lysosomal membrane proteins for degradation. PLoS Biol. 2021, 19, e3001361. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Zhai, W.; Lu, X.; Wang, G. The Cross-Links of Endoplasmic Reticulum Stress, Autophagy, and Neurodegeneration in Parkinson’s Disease. Front. Aging Neurosci. 2021, 13, 691881. [Google Scholar] [CrossRef]
- Venkatesan, R.; Park, Y.U.; Ji, E.; Yeo, E.J.; Kim, S.Y. Malathion increases apoptotic cell death by inducing lysosomal membrane permeabilization in N2a neuroblastoma cells: A model for neurodegeneration in Alzheimer’s disease. Cell Death Discov. 2017, 3, 17007. [Google Scholar] [CrossRef]
- Bonomo, M.M.; Fernandes, J.B.; Carlos, R.M.; Fernandes, M.N. Mitochondrial and lysosomal dysfunction induced by the novel metal-insecticide [Mg(hesp)2(phen)] in the zebrafish (Danio rerio) hepatocyte cell line (ZF-L). Chem. Biol. Interact. 2019, 307, 147–153. [Google Scholar] [CrossRef]
- Goulart, T.L.; Boyle, R.T.; Souza, M.M. Cytotoxicity of the association of pesticides Roundup Transorb® and Furadan 350 SC® on the zebrafish cell line, ZF-L. Toxicol In Vitro 2015, 29, 1377–1384. [Google Scholar] [CrossRef][Green Version]
- Leemans, M.; Couderq, S.; Demeneix, B.; Fini, J.B. Pesticides with Potential Thyroid Hormone-Disrupting Effects: A Review of Recent Data. Front. Endocrinol. 2019, 10, 743. [Google Scholar] [CrossRef]
- PAN-EU List of Chemicals that Provoke Endocrine Disruption. Available online: https://www.pan-europe.info/ (accessed on 7 July 2022).
- Kar, S.; Sangem, P.; Anusha, N.; Senthilkumaran, B. Endocrine disruptors in teleosts: Evaluating environmental risks and biomarkers. Aquacult. Fish. 2021, 6, 1–26. [Google Scholar] [CrossRef]
- Hara, A.; Hiramatsu, N.; Fujita, T. Vitellogenesis and choriogenesis in fishes. Fish Sci. 2016, 82, 187–202. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Li, Y.W.; Chen, Q.L.; Liu, Z.H. Long-term exposure of xenoestrogens with environmental relevant concentrations disrupted spermatogenesis of zebrafish through altering sex hormone balance, stimulating germ cell proliferation, meiosis and enhancing apoptosis. Environ. Pollut. 2019, 244, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Falfushynska, H.; Gnatyshyna, L.L.; Stoliar, O. Population-related molecular responses on the effect of pesticides in Carassius auratus gibelio. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 396–406. [Google Scholar] [CrossRef]
- Juberg, D.R.; Gehen, S.C.; Coady, K.K.; LeBaron, M.J.; Kramer, V.J.; Lu, H.; Marty, M.S. Chlorpyrifos: Weight of evidence evaluation of potential interaction with the estrogen, androgen, or thyroid pathways. Regul. Toxicol. Pharmacol. 2013, 66, 249–263. [Google Scholar] [CrossRef] [PubMed]
- De Anna, J.S.; Castro, J.M.; Darraz, L.A.; Elías, F.D.; Cárcamo, J.G.; Luquet, C.M. Exposure to hydrocarbons and chlorpyrifos alters the expression of nuclear receptors and antioxidant, detoxifying, and immune response proteins in the liver of the rainbow trout, Oncorhynchus mykiss. Ecotoxicol. Environ. Saf. 2021, 208, 111394. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Singh, A. Chlorpyrifos effect on vitellogenin, ovarian steroid in adult and nr5a1 expression in fry of the freshwater catfish, Heteropneustes fossilis (Bloch, 1794). Asian J. Biol. Life Sci. 2021, 10, 65–75. [Google Scholar] [CrossRef]
- Tian, H.; Ru, S.; Wang, Z.; Cai, W.; Wang, W. Estrogenic effects of monocrotophos evaluated by vitellogenin mRNA and protein induction in male goldfish (Carassius auratus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 150, 231–236. [Google Scholar] [CrossRef]
- Xia, S.; Zhao, Y.B.; Hu Yang, M.Q.J.Y. Induction of vitellogenin gene expression in medaka exposed to glyphosate and potential molecular mechanism. Zhongguo Huanjing Kexue 2013, 33, 1656. [Google Scholar]
- Korkmaz, C.; Dönmez, A.E. Effects of Diazinon on 17β-estradiol, Plasma Vitellogenin and Liver and Gonad Tissues of Common Carp (Cyprinus carpio, L., 1758). Turk. J. Fish. Aquat. Sci. 2017, 17, 627–638. [Google Scholar] [CrossRef]
- Darvishi, M.; Safari, R.; Hoseinifar, S.H.; Shabani, A.; Dadar, M.; Jarayedi, Z.; Paolucci, M. Sublethal doses of diazinon affected reproductive, immune, and oxidative status in female zebrafish (Danio rerio). Aquacult. Rep. 2022, 22, 100944. [Google Scholar] [CrossRef]
- Spano, L.; Tyler, C.R.; van Aerle, R.; Devos, P.; Mandiki, S.N.; Silvestre, F.; Thomé, J.P.; Kestemont, P. Effects of atrazine on sex steroid dynamics, plasma vitellogenin concentration and gonad development in adult goldfish (Carassius auratus). Aquat. Toxicol. 2004, 66, 369–379. [Google Scholar] [CrossRef]
- Suzawa, M.; Ingraham, H.A. The Herbicide Atrazine Activates Endocrine Gene Networks via Non-Steroidal NR5A Nuclear Receptors in Fish and Mammalian Cells. PLoS ONE 2008, 3, e2117. [Google Scholar] [CrossRef] [PubMed]
- Vasanth, S.; Arul, G.; Karthikeyeni, S.; Kumar, T.S.; Vignesh, V.; Manimegalai, M.; Bupesh, G.; Thirumurugan, R.; Subramanian, P. Influence of Triazine Herbicide Exposure on Guppies (Poecilia sphenops) Aromatase Activities, Altered Sex Steroid Concentration and Vitellogenin Induction. Indian J. Pharm. Sci. 2015, 77, 156–162. [Google Scholar] [CrossRef]
- Lau, E.S.; Zhang, Z.; Qin, M.; Ge, W. Knockout of Zebrafish Ovarian Aromatase Gene (cyp19a1a) by TALEN and CRISPR/Cas9 Leads to All-male Offspring Due to Failed Ovarian Differentiation. Sci. Rep. 2016, 6, 37357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, L.; Yang, K.; Tian, H.; Wang, W.; Ru, S. Monocrotophos pesticide modulates the expression of sexual differentiation genes and causes phenotypic feminization in zebrafish (Danio rerio). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2013, 157, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Schilirò, T.; Gea, M.; Bianchi, S.; Spinello, A.; Magistrato, A.; Gilardi, G.; Di Nardo, G. Molecular Basis for Endocrine Disruption by Pesticides Targeting Aromatase and Estrogen Receptor. Int. J. Environ. Res. Public Health 2020, 17, 5664. [Google Scholar] [CrossRef]
- Vinggaard, A.M.; Hnida, C.; Breinholt, V.; Larsen, J.C. Screening of selected pesticides for inhibition of CYP19 aromatase activity in vitro. Toxicol. In Vitro 2000, 14, 227–234. [Google Scholar] [CrossRef]
- Holzer, G.; Besson, M.; Lambert, A.; François, L.; Barth, P.; Gillet, B.; Hughes, S.; Piganeau, G.; Leulier, F.; Viriot, L.; et al. Fish larval recruitment to reefs is a thyroid hormone-mediated metamorphosis sensitive to the pesticide chlorpyrifos. eLife 2017, 6, e27595. [Google Scholar] [CrossRef]
- Zhang, X.; Tian, H.; Wang, W.; Ru, S. Monocrotophos pesticide decreases the plasma levels of total 3,3′,5-triiodo-l-thyronine and alters the expression of genes associated with the thyroidal axis in female goldfish (Carassius auratus). PLoS ONE 2014, 9, e108972. [Google Scholar] [CrossRef]
- Otênio, J.K.; Souza, K.D.; Alberton, O.; Alberton, L.R.; Moreno, K.G.T.; Gasparotto Junior, A.; Palozi, R.A.C.; Lourenço, E.L.B.; Jacomassi, E. Thyroid-disrupting effects of chlorpyrifos in female Wistar rats. Drug Chem. Toxicol. 2022, 45, 387–392. [Google Scholar] [CrossRef]
- Suárez, B.; Vela-Soria, F.; Castiello, F.; Olivas-Martinez, A.; Acuña-Castroviejo, D.; Gómez-Vida, J.; Olea, N.; Fernández, M.F.; Freire, C. Organophosphate pesticide exposure, hormone levels, and interaction with PON1 polymorphisms in male adolescents. Sci. Total Environ. 2021, 769, 144563. [Google Scholar] [CrossRef]
- Li, J.; Xu, Y.; Li, N.; Zuo, R.; Zhai, Y.; Chen, H. Thyroid Hormone Disruption by Organophosphate Esters Is Mediated by Nuclear/Membrane Thyroid Hormone Receptors: In Vitro, In Vivo, and In Silico Studies. Environ. Sci. Technol. 2022, 56, 4241–4250. [Google Scholar] [CrossRef] [PubMed]
- Blount, J.R.; Shields, J.N.; Wu, C.-C.; Akemann, C.; Baker, B.B.; Siddiqua, Z.; Meyer, D.N.; Hijazi, N.; Tolbert, Z.; Pitts, D.K.; et al. Phenotypic and transcriptomic effects of developmental exposure to nanomolar levels of pesticides in zebrafish. Environ. Adv. 2002, 7, 100151. [Google Scholar] [CrossRef]
- Xiao, X.; Clark, J.M.; Park, Y. Potential contribution of insecticide exposure and development of obesity and type 2 diabetes. Food Chem. Toxicol. 2017, 105, 456–474. [Google Scholar] [CrossRef] [PubMed]
- Plisetskaya, E.M. Physiology of fish endocrine pancreas. Fish Physiol. Biochem. 1989, 7, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bui-Nguyen, T.M.; Baer, C.E.; Lewis, J.A.; Yang, D.; Lein, P.J.; Jackson, D.A. Dichlorvos exposure results in large scale disruption of energy metabolism in the liver of the zebrafish, Danio rerio. BMC Genom. 2015, 16, 853. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.L.; Hou, Y.C.; Wang, I.K.; Lu, K.C.; Yen, T.H. Organophosphate pesticides and new-onset diabetes mellitus: From molecular mechanisms to a possible therapeutic perspective. World J. Diabetes 2021, 12, 1818–1831. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatib, I.; Rychter, P.; Falfushynska, H. Pesticide Pollution: Detrimental Outcomes and Possible Mechanisms of Fish Exposure to Common Organophosphates and Triazines. J. Xenobiot. 2022, 12, 236-265. https://doi.org/10.3390/jox12030018
Khatib I, Rychter P, Falfushynska H. Pesticide Pollution: Detrimental Outcomes and Possible Mechanisms of Fish Exposure to Common Organophosphates and Triazines. Journal of Xenobiotics. 2022; 12(3):236-265. https://doi.org/10.3390/jox12030018
Chicago/Turabian StyleKhatib, Ihab, Piotr Rychter, and Halina Falfushynska. 2022. "Pesticide Pollution: Detrimental Outcomes and Possible Mechanisms of Fish Exposure to Common Organophosphates and Triazines" Journal of Xenobiotics 12, no. 3: 236-265. https://doi.org/10.3390/jox12030018
APA StyleKhatib, I., Rychter, P., & Falfushynska, H. (2022). Pesticide Pollution: Detrimental Outcomes and Possible Mechanisms of Fish Exposure to Common Organophosphates and Triazines. Journal of Xenobiotics, 12(3), 236-265. https://doi.org/10.3390/jox12030018