1. Introduction
Food loss and waste are not only an economic loss to farmers and to supply chain businesses, but also waste energy and other resources. Furthermore, food loss and waste are major contributors to anthropogenic greenhouse gas emissions, and in 2017 half the greenhouse gas emissions from food systems were attributed to food loss and waste [
1]. Fruit and vegetables have the largest proportion of production lost or wasted because of their short shelf life [
2], exacerbated by innate peaks in production, making it difficult for growers to match supply to the very volatile demand in the UK retail environment [
3]. Strawberries have a typical shelf-life of 7 days under optimum conditions [
4] and have a distinct production peak [
5]. If demand drops unexpectedly, surplus production is either lost because it is not economic to harvest, wasted after harvest because it remains unsold, or it may be sold but at a heavily discounted price, leading to economic loss for growers [
6]. Strawberry growers would value technologies such as plant growth regulators (PGRs) to temporarily delay production until demand increases again [
6], but there are no PGRs available for delaying production in strawberries.
We have proposed that polymers that physically block stomata (stomatal blockers) and restrict photosynthesis and assimilate supply for growth may have potential, in the absence of PGRs, for delaying the production of strawberries and other fresh produce [
7]. Much research in the 1960s and 1970s studied the effects of stomatal blocking hydrophobic polymers on leaf gas exchange [
8], conclusively demonstrating that a wide range of polymers sprayed on leaves reduces both transpiration and photosynthesis. This led to commercial use, mainly as antitranspirants [
9]. The ability of these polymers to act as PGRs in grapevines by restricting photosynthesis and excessive yield at maturity, and thereby improving berry quality, has previously been demonstrated [
10]. Here, we present proof-of-concept results from two experiments to test the hypothesis that stomatal blockers can delay strawberry production. The experiments were conducted using two cultivars of glasshouse-grown strawberry plants in the UK spring and summer with a commercial stomatal blocker applied to the plants during early fruit ripening. Production curves were fitted to the cumulative yield to enable the delay to be quantified.
2. Materials and Methods
The timing of all tasks conducted on the experiments is shown in
Table 1. Forty plants of a June-bearer cultivar (Malling Centenary, Experiment [Exp] 1) were planted in eight growbags and thirteen days later, forty plants of an everbearer cultivar (Favori, Exp 2) were planted in another eight growbags in an unheated glasshouse with natural daylight at Harper Adams University (52°46′ N, 2°25′ W). The growbags were the same as the growbags used by commercial strawberry growers in the UK [
6] (02 mix Safeguard plus, Cocogreen [UK] Limited, Manchester, UK). For each experiment, growbags were placed end-to-end on three tables in the glasshouse. Two tables each with three growbags were 1.6 m apart and were both oriented North–South. Space restriction in the glasshouse meant that the third table, with two growbags, had to be perpendicular to the other two and oriented East–West 0.5 m from the ends of the other two tables. Nutrients and water were provided by automated drip fertigation, with four drippers in each growbag, for 3 min every day. The initial nutrient mix for vegetative growth (Starter, Solufeed Limited, Chichester, UK) was at a concentration of 1.5 kg/15 L diluted 1:50. When the fruits started forming, this was changed to a nutrient mix for fruit production (SF-C, Solufeed Limited, Chichester, UK) at the same rate. Runners were regularly removed. At early fruit ripening, a hand-held sprayer was used to apply treatments to individual plants, separated by a guard, to provide complete coverage of leaves. Because no obvious positional differences were seen in growth between growbags or tables, the treatments were allocated to the 40 plants in each experiment in a completely randomised design. In each experiment, twenty replicate plants were sprayed with water, and twenty replicate plants were sprayed with 0.5%
v/
v Nu-Film P (96% poly-1-
p-menthene; Miller Chemical, Hanover, PA, USA).
Thermal images were taken 5 days after application with a FLIR T420bx thermal camera (Model: FLIR-T62101, Teledyne FLIR, West Malling, UK). The camera was positioned at approximately 0.5 m from the leaf canopy of each plant and at a 40~45° angle. All images were taken during the period of 11:00 a.m. to 13:00 p.m. and were analysed with FLIR Tools software (Teledyne FLIR, West Malling, UK). The temperature of three leaves per plant was recorded and the mean leaf temperature for each plant was calculated for analysis. Completely ripe berries were harvested at irregular intervals over a period of 23 days after spraying. Every berry was weighed, and the maximum diameter was measured. The sugar content in °Brix was determined separately for every berry with an analogue refractometer (Model: Bellingham + Stanley Eclipse, Xylem Analytics, Oberbayern, Germany). The number of berries harvested and measured varied between about 100 and 200 berries for each treatment in each experiment. The mean diameter and mean °Brix were calculated for each plant for data analysis.
Data analysis was conducted either in Excel or Genstat (23rd edition, VSNi, Hemel Hempstead, UK). The cumulative mean yield from all replicate plants for each treatment at each harvest date was calculated and a single analysis was conducted over the entire harvest period to test whether the pattern of production of berry yield differed between treatments. The analysis was conducted by fitting regression lines to the curves for each treatment in each experiment using a polynomial regression in groups procedure, with significance tested in the regression ANOVA. Once the significant lines to fit were determined from the regression ANOVA, the lines were drawn on the cumulative yield graph and the patterns could be described with confidence and the delay in production quantified from the curves. Because the harvest dates were irregular, the curve was first smoothed by aggregating some harvest dates and calculating the cumulative yield in every 3-day period to give one cumulative yield data point every 3 days for Exp 1. Because very few berries ripened initially in Exp 2, the first cumulative yield data point was calculated after 7 days, thereafter in every 3-day period.
The mean leaf temperature per plant, berry diameter, and °Brix were analysed with a completely randomised ANOVA comparing the two treatments. For all ANOVAs, p = 0.05 was used to determine significance, and the validity of all analyses was checked by inspecting the residual graphs. For a few analyses, outliers were clearly visible in the graphs of residuals against fitted values and these outliers were omitted and the analyses re-run, providing fewer degrees of freedom for these analyses.
3. Results
The mean leaf temperature was not significantly affected by the blocker either in Exp 1 (water 22.8 °C and blocker 22.8 °C, p = 0.865, SED 0.273, 36 DF) or in Exp 2 (water 22.6 °C and blocker 22.8 °C, p = 0.293, SED 0.222, 37 DF).
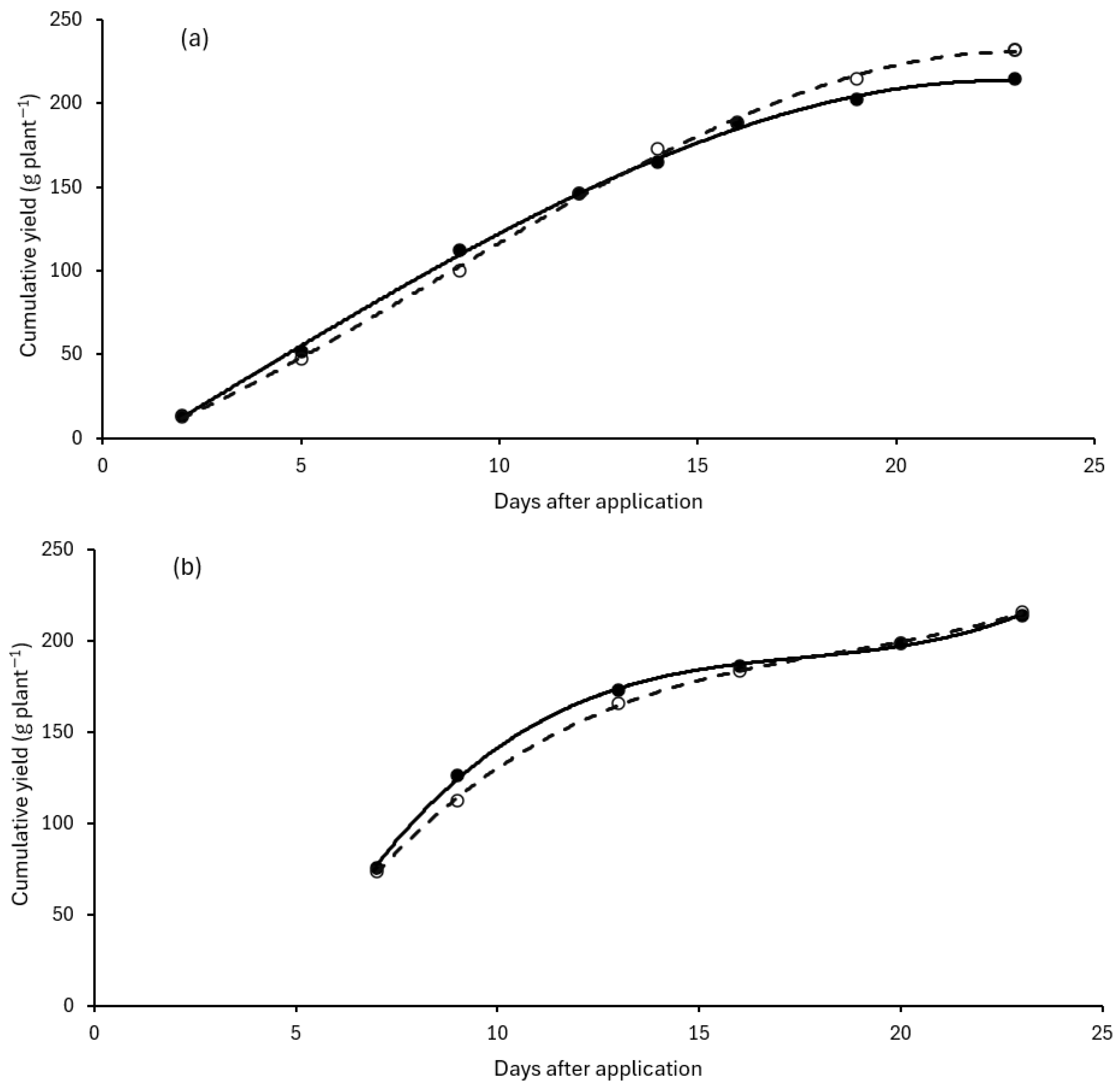
The polynomial regression in groups of cumulative yield curves over time showed that a cubic relationship was significant and that the interaction between days after application and treatment for the June-bearer (Exp 1) was very highly significant (
p < 0.001), and for the everbearer (Exp 2) it was borderline significant (
p = 0.052), indicating that separate cubic curves for each treatment are appropriate for both cultivars (
Table 2,
Figure 1). By using statistical significance to guide the choice of line to fit, there is statistical confidence in drawing conclusions directly from the fitted lines.
The significant interaction between the cubic polynomial and the treatment, together with visual inspection of the fitted curves, shows that the blocker delayed production in both cultivars. The blocker started to impact production and induce a delay a few days after spraying and the delay continued for most of the period when production was increasing rapidly. The magnitude of the delay was small: it took about 1 to 2 days in both cultivars for the blocker-treated plants to gain the same cumulative yield as the water-treated plants. The delay diminished after about 9 days after spraying in the June-bearer and after about 13 days in the everbearer. Equal yield occurred 13 days after application in the June-bearer, but took longer in the blocker-treated everbearer which did not return to the same yield as the water-treated plants until 18 days after application. Subsequently yield of the June-bearer rose above the yield of the water-treated plants to provide 8% greater cumulative yield of berries at 23 days after application. In contrast, there was negligible difference at the end of the harvest period in the everbearer with only 1% more cumulative yield at 23 days after application.
There appeared to be no change in visual defects on the berries from applying the blocker, and the two quantitative measures of berry quality (berry diameter and sugar content) showed no significant change in response to blocker (
Table 3).
4. Discussion
It is not clear why no evidence was found for the expected greater leaf temperature from blocked stomata leading to less transpirational cooling of plants treated with poly-1-
p-menthene. The leaf temperature measurements were, however, taken 5 days after application when the blocker was probably already degrading, and this would have led to a smaller temperature rise than if measured earlier. This conclusion is supported by results of previous research on poly-1-
p-menthene applied to well-watered rapeseed, which showed that the initial reduction in stomatal conductance was already diminishing by 3 days after application [
11]. In addition, a lack of pollinators in the enclosed glasshouse may have increased variability of the plants, making it harder to detect small differences in leaf temperature.
The delayed yield after application in both cultivars provides evidence in support of our hypothesis that stomatal blockers can retard strawberry production. This is consistent with the well-established effects of blockers in reducing gas exchange in many species [
7]. The delay was small and may not be sufficient to be useful commercially. Increasing the concentration of the blocker may increase the length of the delay, since research in rapeseed has found greater effects on reducing gas exchange from higher concentrations of the similar blocker di-1-
p-menthene [
12]. Using di-1-
p-menthene may also prolong the period of delay of production as it is more persistent than poly-1-
p-menthene [
11,
13].
It is not clear why the blocker-treated June-bearer plants provided a greater yield at the end of the harvest period, but previous research on a petroleum-based polymer applied to
Vicia faba leaves may provide a possible reason [
14]. This previous research showed that the polymer increased leaf water potential, which in turn increased the turgidity of the guard cells and induced wider opening of stomata under the polymer film. In addition, stomata on areas of the leaf not covered by the film opened wider. If the wider stomatal opening persisted after the polymer degraded to provide increased photosynthesis in the leaf, this may have resulted in greater yield. The yield increase did not occur in the everbearer cultivar, but this cultivar took 5 days longer than the June-bearer cultivar for blocker-treated plants to compensate for the delay and reach the same yield as the water-treated plants. Thus, everbearer berries were only harvested for a further 5 days compared with a further 10 days after the time of equal yield in the June-bearer. It is possible that, given a longer harvesting period, a greater yield may also have occurred in the everbearer cultivar.
The increased yield of the blocker-treated June-bearer plants may be an example of the phenomenon of overcompensation in plant growth. This is a well-known phenomenon in wild plant species after damage occurs, often from herbivores [
15]. Overcompensation in growth has been reported to give greater crop yield in wheat after transient shading [
16], in potatoes after insect pest damage to tubers [
17], and in maize after leaf removal [
18], but no reports have been found for fruit crops. More evidence is needed to confirm whether overcompensation can occur in strawberries from application of a stomatal blocker.
Visual defects, berry diameter, and sugar content are important quality criteria for retailers [
6], and the lack of effect of the blocker on berry quality at harvest will be an important advantage if stomatal blockers are to be used commercially on the strawberry crop. Before commercial adoption of stomatal blockers can be contemplated for delaying strawberry production, further experiments will be needed to confirm the yield effects found in these proof-of-concept experiments, test higher concentrations of blocker, test more persistent blockers, study a wider range of cultivars in different growing systems, and make earlier assessments of leaf temperature. The berries in our experiments were not stored postharvest, and it will also be essential to assess quality and consumer acceptability after postharvest storage.
Assuming a commercially useful production delay can be achieved, adoption of stomatal blockers as PGRs will depend on registration. Currently, in the UK, the blocker we used (poly-1-
p-menthene) is approved for use only as an adjuvant in mixture with pesticides, not for use alone on commercial crops [
19]. For use as a PGR in the UK, the blocker would be classified as a Plant Protection Product [
19] with associated data requirements and registration costs. The very short period between application and harvest may restrict registration, but since plant oils used as food can act as stomatal blockers [
20,
21], these food oils may be more acceptable to registration authorities for application close to the time to harvest. If there are no adverse effects on appearance or flavour, plant oils may also be more acceptable to consumers. Some plant oils normally used as food ingredients have the same approval in the UK as poly-1-
p-menthene, for use as an adjuvant in mixture with pesticides. Plant oils for use as a PGR would, however, require separate registration as a Plant Protection Product under current UK legislation.
In conclusion, we have found preliminary evidence in two cultivars supporting our hypothesis that applying a stomatal blocker can delay strawberry production. In the June-bearer cultivar, the cumulative yield delay was small and cumulative yield returned to the water-treated value 13 days after application, followed by overcompensation to give greater cumulative yield. In the everbearer cultivar, the small cumulative yield delay persisted for longer (18 days) and overcompensation was negligible. Further experiments are necessary to confirm these preliminary results and determine if the potential to use stomatal blockers as PGRs in the strawberry crop can be realised.




{kind=link}