
Calcium-Mediated Modulation of GC Switch Regulates Peroxisomal H2O2 Levels in Response to Wounding in Plants



{kind=link}
Abstract
1. Introduction
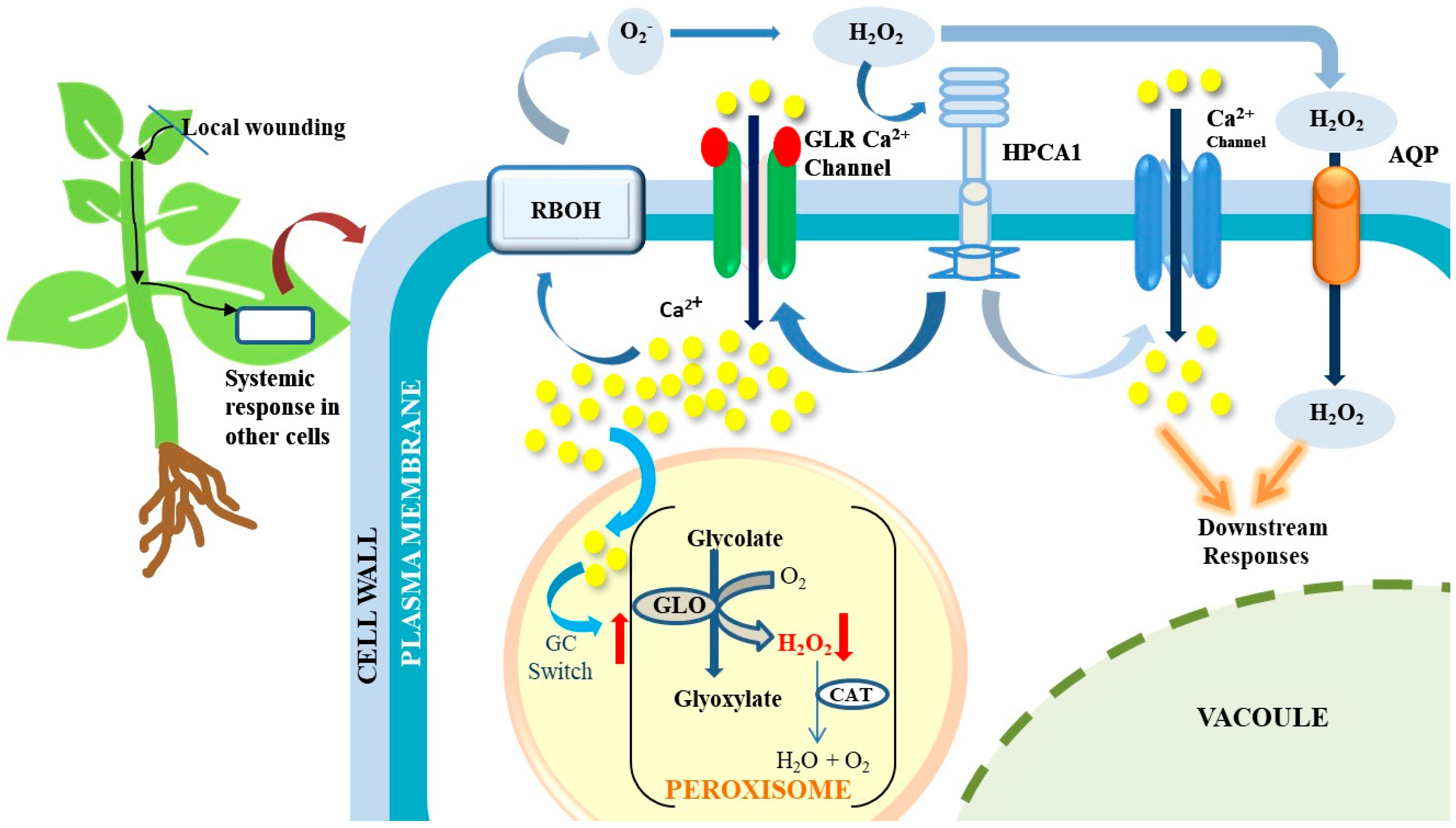
2. GC Switch Facilitates Peroxisomal H2O2 Decrease during Wounding
3. GLR-Mediated Ca2+ Influx Regulates the GC Switch
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fichman, Y.; Mittler, R. Integration of electric, calcium, reactive oxygen species and hydraulic signals during rapid systemic signaling in plants. Plant J. 2021, 107, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.J.; Fichman, Y.; Zandalinas, S.I.; Mittler, R. Jasmonic acid and salicylic acid modulate systemic reactive oxygen species signaling during stress responses. Plant Physiol. 2022, 191, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liao, M.; Huang, J.; Chen, L.; Huang, H.; Wu, K.; Pan, Q.; Zhang, Z.; Peng, X. Dynamic and fluctuating generation of hydrogen peroxide via photorespiratory metabolic channeling in plants. Plant J. 2022, 112, 1429–1446. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K. Calcium Transport Elements in Plants; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Upadhyay, S.K. Calcium Channels, OST1 and Stomatal Defence: Current Status and Beyond. Cells 2023, 12, 127. [Google Scholar] [CrossRef] [PubMed]
- Grenzi, M.; Buratti, S.; Parmagnani, A.S.; Aziz, I.A.; Bernacka-Wojcik, I.; Resentini, F.; Šimura, J.; Doccula, F.G.; Alfieri, A.; Luoni, L.; et al. Long-distance turgor pressure changes induce local activation of plant glutamate receptor-like channels. Curr. Biol. 2023, 33, 1019–1035. [Google Scholar] [CrossRef] [PubMed]
- Furch, A.C.U.; Van Bel, A.J.E.; Fricker, M.D.; Felle, H.H.; Fuchs, M.; Hafke, J.B. Sieve element Ca2+ channels as relay stations between remote stimuli and sieve tube occlusion in Vicia faba. Plant Cell 2009, 21, 2118–2132. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.R.; Mithöfer, A. Electrical long-distance signaling in plants. In Long-Distance Systemic Signaling and Communication in Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 291–308. [Google Scholar]
- Li, X.; Chen, L.; Zeng, X.; Wu, K.; Huang, J.; Liao, M.; Xi, Y.; Zhu, G.; Zeng, X.; Hou, X.; et al. Wounding induces a peroxisomal H2O2 decrease via glycolate oxidase-catalase switch dependent on glutamate receptor like channel supported Ca2+ signaling in plants. Plant J. 2023, 116, 1325–1341. [Google Scholar] [CrossRef]
- Li, X.; Liao, M.; Huang, J.; Xu, Z.; Lin, Z.; Ye, N.; Zhang, Z.; Peng, X. Glycolate oxidase-dependent H2O2 production regulates IAA biosynthesis in rice. BMC Plant Biol. 2021, 21, 326. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, Y.; Xie, Z.; Li, X.; He, Z.; Peng, X. Associationdissociation of glycolate oxidase with catalase in rice: A potential switch to modulate intracellular H2O2 levels. Mol. Plant 2016, 9, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, Z.; Zhang, K.; Zhang, X.; Zhang, Y.; Wang, X.; Han, C.; Yu, J.; Xu, K.; Li, D. Barley stripe mosaic virus cb interacts with glycolate oxidase and inhibits peroxisomal ROS production to facilitate virus infection. Mol. Plant 2018, 11, 338–341. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Alamri, S.; Khan, M.N.; Corpas, F.J.; Al-Amri, A.A.; Alsubaie, Q.D.; Ali, H.M.; Kalaji, H.M.; Ahmad, P. Melatonin and calcium function synergistically to promote the resilience through ROS metabolism under arsenic-induced stress. J. Hazard. Mater. 2020, 398, 122882. [Google Scholar] [CrossRef]
- Lew, T.T.S.; Koman, V.B.; Silmore, K.S.; Seo, J.S.; Gordiichuk, P.; Kwak, S.Y.; Park, M.; Ang, M.C.-Y.; Khong, D.T.; Lee, M.A.; et al. Real-time detection of wound-induced H2O2 signaling waves in plants with optical nanosensors. Nat. Plants 2020, 61, 404–415. [Google Scholar] [CrossRef]
- Fichman, Y.; Miller, G.; Mittler, R. Whole-plant live imaging of reactive oxygen species. Mol. Plant 2019, 12, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Chi, Y.; Jiang, Z.; Xu, Y.; Xie, L.; Huang, F.; Wan, D.; Ni, J.; Yuan, F.; Wu, X.; et al. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis. Nature 2020, 578, 577–581. [Google Scholar] [CrossRef]
- Fichman, Y.; Zandalinas, S.I.; Peck, S.; Luan, S.; Mittler, R. HPCA1 is required for systemic reactive oxygen species and calcium cell-to-cell signaling and plant acclimation to stress. Plant Cell 2022, 34, 4453–4471. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Shabala, S. Mechanisms of cytosolic calcium elevation in plants: The role of ion channels, calcium extrusion systems and NADPH oxidase-mediated ‘ROS-Ca2+ Hub’. Funct. Plant Biol. 2018, 45, 9–27. [Google Scholar] [CrossRef]
- Choi, W.G.; Hilleary, R.; Swanson, S.J.; Kim, S.H.; Gilroy, S. Rapid, long distance electrical and calcium signaling in plants. Annu. Rev. Plant Biol. 2016, 67, 287–307. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Spencer, D.; Sawai-Toyota, S.; Jiaqi, W.; Zhang, T.; Koo, A.J.; Howe, G.A.; Gilroy, S. Glutamate triggers long-distance, calcium-based plant defense signaling. Science 2018, 361, 1112–1115. [Google Scholar] [CrossRef]
- Upadhyay, S.K. CPK12 and Ca2+-mediated hypoxia signaling. Plant Signal. Behav. 2023, 18, 2273593. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishu; Shekhawat, J.; Upadhyay, S.K. Calcium-Mediated Modulation of GC Switch Regulates Peroxisomal H2O2 Levels in Response to Wounding in Plants. Int. J. Plant Biol. 2024, 15, 198-202. https://doi.org/10.3390/ijpb15010016
Ishu, Shekhawat J, Upadhyay SK. Calcium-Mediated Modulation of GC Switch Regulates Peroxisomal H2O2 Levels in Response to Wounding in Plants. International Journal of Plant Biology. 2024; 15(1):198-202. https://doi.org/10.3390/ijpb15010016
Chicago/Turabian StyleIshu, Jyoti Shekhawat, and Santosh Kumar Upadhyay. 2024. "Calcium-Mediated Modulation of GC Switch Regulates Peroxisomal H2O2 Levels in Response to Wounding in Plants" International Journal of Plant Biology 15, no. 1: 198-202. https://doi.org/10.3390/ijpb15010016
APA StyleIshu, Shekhawat, J., & Upadhyay, S. K. (2024). Calcium-Mediated Modulation of GC Switch Regulates Peroxisomal H2O2 Levels in Response to Wounding in Plants. International Journal of Plant Biology, 15(1), 198-202. https://doi.org/10.3390/ijpb15010016