
Evaluation of Physiological and Biochemical Parameters and Some Bioindicators of Barium Tolerance in Limbarda crithmoides and Helianthus annuus


 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Plant Culture and Treatments Experimentations
2.2. Chlorophyll Fluorescence
2.3. Chlorophyll Content Determination
2.4. Minerals and Barium Accumulation
2.5. Total Inorganic Nitrogen
2.6. Polyphenols and Flavonoids
2.7. Statistical Analysis
3. Results and Discussion
3.1. Plant Growth
3.2. Chlorophyll Fluorescence
3.3. Chlorophylls Contents
3.4. Polyphenols and Flavonoids Levels
3.5. Nutrients and Total Inorganic Nitrogen Contents
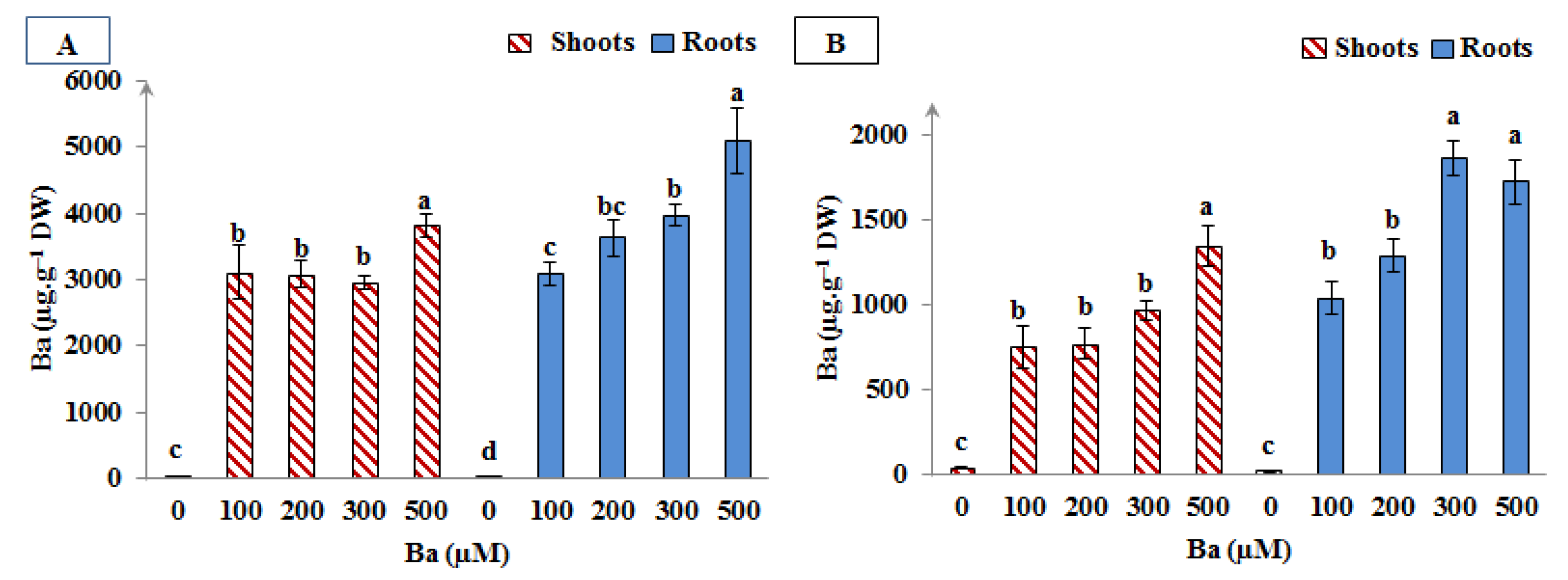
3.6. Barium Content
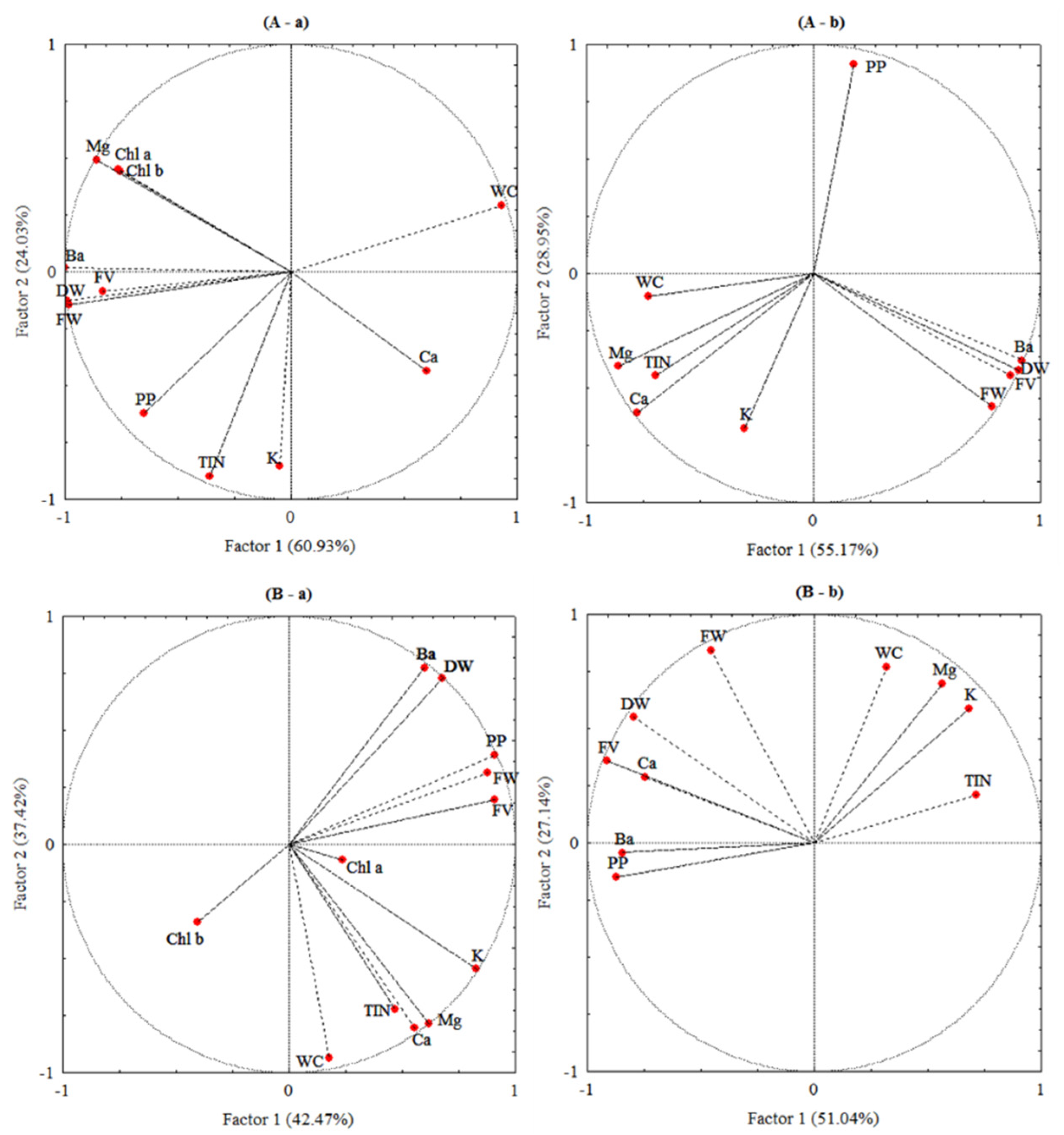
3.7. Correlation Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kresse, R.; Baudis, U.; Jager, P.; Riechers, H.H.; Wagner, H.; Winkler, J.; Wolf, H.U. Barium and Barium Compounds. Encycl. Ind. Chem. 2007, 4, 621–640. [Google Scholar] [CrossRef]
- Oskarsson, A. Barium. In Handbook on the Toxicology of Metals, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2015; Volume II. [Google Scholar] [CrossRef]
- ATSDR (Agency for Toxic Substances and Disease Registry). Toxicological Profile for Barium; Agency for Toxic Substances and Disease Registry; US Department of Health and Human Services: Atlanta, GA, USA, 2007.
- Abreu, C.A.; Cantoni, M.; Coscione, A.R.; Paz-Ferreiro, J. Organic Matter and Barium Absorption by Plant Species Grown in an Area Polluted with scrap metal residue. Appl. Environ. Soil Sci. 2012, 2012, 476821. [Google Scholar] [CrossRef]
- Madejon, P. Barium. In Heavy Metals in Soils: Trace Metals and Metalloids in Soils and their Bioavailability, Environmental Pollution; Alloway, B.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 22, pp. 507–514. [Google Scholar] [CrossRef]
- Horner, T.J.; Pryer, H.V.; Nielsen, S.G.; Crockford, P.W.; Gauglitz, J.M.; Wing, B.A.; Ricketts, R.D. Pelagic barite precipitation at micromolar ambient sulfate. Nat. Commun. 2017, 8, 1342. [Google Scholar] [CrossRef] [PubMed]
- Viana, D.G.; Filho, F.B.E.; Pires, F.R.; Soares, M.B.; Ferreira, A.D.; Bonomo, R.; Martins, L.F. In situ barium phytoremediation in flooded soil using Typha domingensis under different planting densities. Ecotoxicol. Environ. Saf. 2021, 210, 111890. [Google Scholar] [CrossRef]
- Lamb, D.T.; Matanitobua, V.P.; Palanisami, T.; Megharaj, M.; Naidu, R. Bioavailability of barium to plants and invertebrates in soils contaminated by barite. Environ. Sci. Technol. 2013, 47, 4670–4676. [Google Scholar] [CrossRef]
- Verbruggen, E.M.J.; Smit, C.E.; Van-Vlaardingen, P.L.A. Environmental quality standards for barium in surface water. Proposal for an update according to the methodology of the Water Framework Directive. In RIVM Official Reports; National Institute for Public Health and the Environment: Utrecht, The Netherlands, 2020; pp. 2–111. [Google Scholar] [CrossRef]
- Nogueira, T.A.R.; de Melo, W.J.; Fonseca, I.M.; Marques, M.O.; He, Z. Barium uptake by maize plants as affected by sewage sludge in a long-term field study. J. Hazard. Mater. 2010, 181, 1148–1157. [Google Scholar] [CrossRef]
- Bouslimi, H.; Ferreira, R.; Dridi, N.; Brito, P.; Martins-Dias, S.; Caçador, I.; Sleimi, N. Effects of Barium stress in Brassica juncea and Cakile maritima: The indicator role of some antioxidant enzymes and secondary metabolites. Phyton Int. J. Exp. Bot. 2021, 90, 145–158. [Google Scholar] [CrossRef]
- Kravchenko, J.; Darrah, T.H.; Miller, R.K.; Lyerly, H.K.; Vengosh, A. A review of the health impacts of barium from natural and anthropogenic exposure. Environ. Geochem. Health 2014, 36, 797–814. [Google Scholar] [CrossRef]
- Marisamy, K.; Sevugaperumal, R.; Ramasubramanian, V. Changes in growth, biochemical and enzymatic characteristics of Cyamopsis tetragonoloba (l.) Taub under metal stress due to barium. Int. J. Biol. Pharm. Res. 2015, 6, 935–938. [Google Scholar]
- Marisamy, K.; Duraipandian, M.; Sevugaperumal, R.; Ramasubramanian, V. Estimation of Barium Toxicity Mitigating Efficacy of Amaranthus caudatus L. Univers. J. Environ. Res. Technol. 2015, 5, 295–305. [Google Scholar]
- Coscione, A.R.; Berton, R.S. Barium extraction potential by mustard, sunflower and castor bean. Sci. Agric. 2009, 66, 59–63. [Google Scholar] [CrossRef][Green Version]
- Sleimi, N.; Bankaji, I.; Dallai, M.; Kefi, O. Accumulation des éléments traces et tolérance au stress métallique chez les halophytes colonisant les bordures de la lagune de Bizerte. Rev. D’Ecologie Terre Vie 2014, 69, 49–59. [Google Scholar]
- Ghabriche, R.; Ghnaya, T.; Mnasri, M.; Zaier, H.; Baioui, R.; Vromman, D.; Abdelly, C.; Lutts, S. Polyamine and tyramine involvement in NaCl-induced improvement of Cd resistance in the halophyte Inula chrithmoides L. J. Plant Physiol. 2017, 216, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, E.J. Sand and water culture methods used in the study of plant nutrition. J. Assoc. Off. Anal. Chem. 1966, 49, 888–889. [Google Scholar]
- Kouki, R.; Ayachi, R.; Ferreira, R.; Sleimi, N. Behavior of Cucumis sativus L. in presence of aluminum stress: Germination, plant growth, and antioxidant enzymes. Food Sci. Nutr. 2021, 9, 3280–3288. [Google Scholar] [CrossRef]
- Sleimi, N.; Abdelly, C. Salt-tolerance strategy of two halophytes species: Spartina alterniflora and Suaeda fruticosa. In Tasks for Vegetation Science; Cash Crop Halophytes; Lieth, H., Mochtchenko, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherland, 2003; Volume 38, pp. 79–85. [Google Scholar] [CrossRef]
- Dridi, N.; Ferreira, R.; Bouslimi, H.; Brito, P.; Martins-Dias, S.; Caçador, I.; Sleimi, N. Assessment of tolerance to lanthanum and cerium in Helianthus annuus plant: Effect on growth, mineral nutrition and secondary metabolism. Plants 2022, 11, 988. [Google Scholar] [CrossRef]
- Labidi, O.; Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M.; Sleimi, N. Assessing of growth, antioxidant enzymes, and phytohormone regulation in Cucurbita pepo under cadmium stress. Food Sci. Nutr. 2021, 9, 2021–2031. [Google Scholar] [CrossRef]
- Sghaier, D.B.; Duarte, B.; Bankaj, I.; Caçador, I.; Sleimi, N. Growth, chlorophyll fluorescence and mineral nutrition in the halophyte Tamarix gallica cultivated in combined stress conditions: Arsenic and NaCl. J. Photochem. Photobiol. B Biol. 2015, 149, 204–214. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Guo, P. Chlorophyll fluorescence: A useful tool in barley plant breeding programs. Photochem. Res. Prog. 2008, 29, 448–471. [Google Scholar]
- Bankaji, I.; Caçador, I.; Sleimi, N. Assessing of tolerance to metallic and saline stresses in the halophyte Suaeda fruticosa: The indicator role of antioxidative enzymes. Ecol. Indic. 2016, 64, 297–308. [Google Scholar] [CrossRef]
- Ayvaz, M.; Koyuncu, M.; Guven, A.; Fagerstedt, K.V. Does boron affect hormone levels of barley cultivars? Eur. Asian J. Biosci. 2012, 120, 113–120. [Google Scholar] [CrossRef]
- Sleimi, N.; Bankaji, I.; Kouki, R.; Dridi, N.; Duarte, B.; Caçador, I. Assessment of extraction methods of trace metallic elements in plants: Approval of a common method. Sustainability 2022, 14, 1428. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 11th ed.; Etats-Unis; Association of Official Analytical Chemists: Washington, DC, USA, 1970. [Google Scholar]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant Activity and Total Phenolics in Selected Fruits, Vegetables, and Grain Products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Lamaison, J.L.; Carnat, A. Teneurs en acide rosmarinique, en dérivés hydroxycinnamiques totaux et activités antioxydantes chez les Apiacées, les Borraginacées et les Lamiacées médicinales. Pharm. Acta Helv. 1990, 65, 315–320. [Google Scholar]
- Sleimi, N.; Kouki, R.; Hadj-Ammar, M.; Ferreira, R.; Pérez-Clemente, R. Barium effect on germination, plant growth, and antioxidant enzymes in Cucumis sativus L. plants. Food Sci. Nutr. 2021, 9, 2086–2094. [Google Scholar] [CrossRef] [PubMed]
- Suwa, R.K.; Jayachandran, N.T.; Nguyen, A.; Boulenouar, K.; Fujita, K.; Saneoka, H. Barium toxicity effects in soybean plants. Arch. Environ. Contam. Toxicol. 2008, 55, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Kaur, L.; Gadgil, K.; Sharma, S. Comparative study of natural phytoextraction and induced phytoextraction of lead using mustard plant (Brassica juncea arawali). Int. J. Bioassay 2012, 2, 352–357. [Google Scholar]
- Caçador, I.; Duarte, B.; Marques, J.C.; Sleimi, N. Carbon mitigation: A salt marsh ecosystem service in times of change. In Halophytes for Food Security in Dry Lands; Khan, M.A., Ozturk, M., Gul, B., Ahmed, M.Z., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2016; pp. 83–110. [Google Scholar] [CrossRef]
- Monteiro, F.A.; Nogueirol, R.C.; Melo, L.C.A.; Artur, A.G.; da Rocha, F. Effect of barium on growth and macronutrient nutrition in Tanzania guinea grass grown in nutrient solution. Commun. Soil Sci. Plant Anal. 2011, 42, 1510–1521. [Google Scholar] [CrossRef]
- Ribeiro, P.R.C.; Viana, D.G.; Pires, F.R.; Filho, F.B.E.; Bonomo, R.; Filho, A.C.; Martins, L.F.; Cruz, L.B.S.; Nascimento, M.C.P. Selection of plants for phytoremediation of barium-polluted flooded soils. Chemosphere 2018, 206, 522–530. [Google Scholar] [CrossRef]
- Zhen, Y.; Qi, J.L.; Wang, S.S.; Su, J.; Xu, G.H.; Zhang, M.S.; Miao, L.V.; Peng, X.X.; Tian, D.; Yang, Y.H. Comparative proteome analysis of differentially expressed proteins induced by Al toxicity in soybean. Physiol. Plant. 2007, 131, 542–554. [Google Scholar] [CrossRef]
- Li, H.; Yang, L.T.; Qi, Y.P.; Guo, P.; Lu, Y.B.; Chen, L.S. Aluminum toxicity-induced alterations of leaf proteome in two citrus species differing in aluminum tolerance. Int. J. Mol. Sci. 2016, 17, 1180. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Combined effect of 24-epibrassinolide and salicylic acid mitigates lead (Pb) toxicity by modulating various metabolites in Brassica juncea L. seedlings. Protoplasma 2017, 255, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Bankaji, I.; Pérez-Clemente, R.M.; Caçador, I.; Sleimi, N. Accumulation potential of Atriplex halimus to zinc and lead combined with NaCl: Effects on physiological parameters and antioxidant enzymes activities. S. Afr. J. Bot. 2019, 123, 51–61. [Google Scholar] [CrossRef]
- Srivastava, S.; Mishra, S.; Dwivedi, S.; Baghel, V.S.; Verma, S.; Tandon, P.K. Barium Phytoremediation Potential of Broad Bean, ViciafabaL., and Its Biochemical Responses. Bull. Environ. Contam. Toxicol. 1974, 74, 715–724. [Google Scholar] [CrossRef]
- Ikram, K.; Abdelhakim, R.Y.H.; Topcuoglu, B. Physiological and biochemical response of Atriplex canescens (pursh) nutt under metallic stress. Plant Arch. 2019, 19, 2747–2752. [Google Scholar]
- Sosnowski, W.; Drozłowska, E. Effect of barium chloride on growth and oxidative stress of saltwater algae. World Sci. News 2018, 111, 121–130. [Google Scholar]
- Bangajavalli, S.; Selvaraj, K.; Esakkiammal, B. Retrieval of barium affected Phaseolus mungo L. by Ulva lactuca. Int. J. Bot. Stud. 2021, 6, 59–63. [Google Scholar]
- Kumar, A.; Narasimha, M.P.V. Plant-lead interactions: Transport, toxicity, tolerance, and detoxification mechanisms. Ecotoxicol. Environ. Saf. 2018, 166, 401–418. [Google Scholar] [CrossRef]
- Bücker-Neto, L.; Paiva, L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between plant hormones and heavy metals responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef]
- Nomura, T.; Itouga, M.; Kojima, M.; Kato, Y.; Hasezawa, S. Copper mediates auxin signalling to control cell differentiation in the copper moss Scopelophila cataractae. J. Exp. Bot. 2015, 66, 1205–1213. [Google Scholar] [CrossRef]
- Kalaji, M.H.; Carpentier, R.; Allakhverdiev, S.I.; Bosa, K. Fluorescence parameters as early indicators of light stress in barley. J. Photochem. Photobiol. B. 2012, 112, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Reboredo, F.H.; Pelica, J.; Lidon, F.C.; Pessoa, M.F.; Silva, M.M.; Guerra, M.; Leitão, R.; Ramalho, J.C. The Tolerance of Eucalyptus globulus to Soil Contamination with Arsenic. Plants 2021, 10, 627. [Google Scholar] [CrossRef]
- Li, S.; Yang, W.; Yang, T.; Chen, Y.; Ni, W. Effects of Cadmium Stress on Leaf Chlorophyll Fluorescence and Photosynthesis of Elsholtzia argyi—A Cadmium Accumulating Plant. Int. J. Phytoremed. 2015, 17, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Van Heerden, P.D.R.; Swanepoel, J.W.; Kruger, G.H.J. Modulation of photosynthesis by drought in two desert scrub species exhibiting C3-mode CO2 assimilation. Environ. Exp. Bot. 2007, 61, 124–136. [Google Scholar] [CrossRef]
- Gomes, M.T.G.; da Luz, A.C.; dos Santos, M.R.; Batitucci, M.D.C.P.; Silva, D.M.; Falqueto, A.R. Drought tolerance of passion fruit plants assessed by the OJIP chlorophyll a fluorescence transient. Sci. Hortic. 2012, 142, 49–56. [Google Scholar] [CrossRef]
- Liu, M.S.; Huang, X.H.; Wang, R.J.; Xu, H.Y.; Zhu, F. Inhibition of photosynthesis in Melia azedarach and Ligustrum lucidum induced by manganese toxicity using OJIP chlorophyll a fluorescence transient. Photosynthetica 2021, 59, 148–159. [Google Scholar] [CrossRef]
- Lazar, D.; Jablonsky, J. On the approaches applied in formulation of a kinetic model of photosystem II: Different approaches lead to different simulations of the chlorophyll a fluorescence transients. J. Theor. Biol. 2009, 257, 260–269. [Google Scholar] [CrossRef]
- Panda, D.; Rao, D.N.; Sharma, S.G.; Strasser, R.J.; Sarkar, R.K. Submergence effects on rice genotypes during seedling stage: Probing of submergence driven changes of photosystem II by chlorophyll a fluorescence induction O-J-I-P transients. Photosynthetica 2006, 44, 69–75. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant 2016, 38, 102. [Google Scholar] [CrossRef]
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Chai, T.; Zhang, Y.; Tan, J.; Ma, S. The effects of copper, manganese and zinc on plant growth and elemental accumulation in the manganese-hyperaccumulator Phytolacca americana. J. Plant Physiol. 2012, 169, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Kisa, D.; Elmastas, M.; Ozturk, L.; Kayir, O. Responses of the phenolic compounds of Zea mays under heavy metal stress. Appl. Biol. Chem. 2016, 59, 813–820. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and regulation of plants phenolics in abiotic stress tolerance: An overview. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 157–168. [Google Scholar] [CrossRef]
- Ali, M.B.; Singh, N.; Shohael, A.M.; Hahn, E.J.; Paek, K.Y. Phenolics metabolism and lignin synthesis in root suspension cultures of Panax ginseng in response to copper stress. Plant Sci. 2006, 171, 147–154. [Google Scholar] [CrossRef]
- Anjitha, K.S.; Sameena, P.P.; Puthur, J.T. Functional aspects of plant secondary metabolites in metal stress tolerance and their importance in pharmacology. Plant Stress 2021, 2, 100038. [Google Scholar] [CrossRef]
- Bergmann, E. Bergmann, H.W. Comparing diagrams of plant/leaf analysis presenting by rapid inspection the mineral nutrient element status of agricultural crop plants. Potash Rev. Sub. 1985, 5, 1–10. [Google Scholar]
- Kafkafi, U. The Functions of Plant K in Overcoming Environmental Stress Situations. In Proceedings of the 22nd Colloquium of IPI 1990, Soligorsk, USSR; IPI: Bern, Switzerland, 1990; pp. 81–93. [Google Scholar]
- Alazem, M.; Lin, N.S. Antiviral roles of Abscisic acid in plants. Front. Plant Sci. 2017, 8, 1760. [Google Scholar] [CrossRef]
- Zhao, L.J.; Peralta-Videa, J.R.; Peng, B.; Bandyopadhyay, S.; Corral-Diaz, B.; Osuna-Avila, P.; Montes, M.O.; Keller, A.A.; Gardea-Torresdey, J.L. Alginate modifies the physiological impact of CeO2 nanoparticles in corn seedlings cultivated in soil. J. Environ. Sci. 2014, 26, 382–389. [Google Scholar] [CrossRef]
- Nawab, J.; Ghani, J.; Khan, S.; Khan, M.A.; Ali, A.; Rahman, Z.; Alam, M.; Hesham, A.; Lei, M. Nutrient Uptake and Plant Growth Under the Influence of Toxic Elements. In Sustainable Plant Nutrition under Contaminated Environments, Sustainable Plant Nutrition in a Changing World; Mahmood, Q., Ed.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 75–101. [Google Scholar] [CrossRef]
- Pourrut, B.; Shahid, M.; Douay, F.; Dumat, C.; Pinelli, E. Molecular mechanisms involved in lead uptake, toxicity and detoxification in higher plants. Heavy Met. Stress Plants 2013, 292, 275–281. [Google Scholar] [CrossRef]
- Rastgoo, L.; Alemzadeh, A.; Tale, A.M.; Tazangi, S.E.; Eslamzadeh, T. Effects of copper, nickel and zinc on biochemical parameters and metal accumulation in gouan Aeluropus littoralis. Plant Knowl. J. 2014, 3, 31–38. [Google Scholar]
- Kee, J.C.; Gonzales, M.J.; Ponce, O.; Ramírez, L.; León, V.; Torres, A.; Corpus, M.; Loayza-Muro, R. Accumulation of heavy metals in native Andean plants: Potential tools for soil phytoremediation in Ancash (Peru). Environ. Sci. Pollut. Res. 2018, 25, 33957–33966. [Google Scholar] [CrossRef]
- Kamachi, H.; Kitamura, N.; Sakatoku, A.; Tanaka, D.; Nakamura, S. Barium accumulation in the metalliferous fern Athyrium Yokoscense. Theor. Exp. Plant Physiol. 2015, 27, 99–107. [Google Scholar] [CrossRef]
- Jansen, S.; Broadley, M.R.; Robbrecht, E.; Smets, E. Aluminum hyperaccumulation in angiosperms: A review of Its phylogenetic significance. Bot. Rev. 2002, 68, 235–269. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, Q.; Wang, L.; Liu, W. Cadmium tolerance and accumulation characteristics of Bidens pilosa L. as a potential Cd-hyperaccumulator. J. Hazard. Mater. 2009, 161, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Baycu, G.; Tolunay, D.; Ozden, H.; Csatari, I.; Karadag, S.; Agba, T.; Rognes, S.E. An Abandoned Copper Mining Site in Cyprus and Assessment of Metal Concentrations in Plants and Soil. Int. J. Phytoremediat. 2015, 17, 622–631. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Limbarda crithmoides | |||||||
| Ba (µM) | DW (mg) | FW (mg) | WC (mL·g−1 DW) | TI (%) | |||
| Shoots | Roots | Shoots | Roots | Shoots | Roots | Entire Plant | |
| 0 | 437.5 c ± 43.8 | 68.0 b ± 14.4 | 5887.1 b ± 541.1 | 835.3 b ± 123.5 | 0.926 a ± 0.002 | 0.909 a ± 0.008 | |
| 100 | 576.8 bc ± 76.7 | 82.6 b ± 11.4 | 7182.1 b ± 1025.8 | 812.8 b ±120.5 | 0.919 a ± 0.001 | 0.897 a ± 0.003 | 136.3 ± 20.3 |
| 200 | 669.6 b ± 87.7 | 110.9 b ± 24.0 | 7816.2 b ± 873.3 | 976.2 b ± 207.3 | 0.916 a ± 0.004 | 0.887 a ± 0.004 | 164.9 ± 21.2 |
| 300 | 761.4 ba ± 57.9 | 119.0 a ± 13.3 | 8283.4 ba ± 1651.3 | 1000.4 b ± 117.7 | 0.917 a ± 0.003 | 0.878 a ± 0.007 | 188.4 ± 16.4 |
| 500 | 875.9 a ± 72.2 | 168.1 a ± 18.8 | 9769.7 a ± 694.7 | 1500.2 a ± 134.3 | 0.911 a ± 0.002 | 0.889 a ± 0.006 | 227.0 ± 21.1 |
| Helianthus annuus | |||||||
| Ba (µM) | DW (mg) | FW (mg) | WC (mL·g−1 DW) | TI (%) | |||
| Shoots | Roots | Shoots | Roots | Shoots | Roots | Entire Plant | |
| 0 | 1807.7 a ± 105.3 | 539.3 a ± 31.7 | 12,664.5 a ± 554.6 | 8520.8 a ± 794.8 | 0.858 a ± 0.003 | 0.935 a ± 0.003 | |
| 100 | 1924.6 a ± 168.6 | 603.1 a ± 71.6 | 13,428.9 a ± 1046.5 | 9764.9 a ± 1081.9 | 0.867 a ± 0.006 | 0.938 a ± 0.002 | 102.4 ± 11.1 |
| 200 | 1931.2 a ± 156.6 | 491.5 a ± 34.8 | 14,100.7 a ± 1024.5 | 8363.1 a ± 942.0 | 0.863 a ± 0.005 | 0.939 a ± 0.004 | 103.6 ± 6.8 |
| 300 | 2097.5 a ± 134.4 | 548.5 a ± 59.2 | 14,543.0 a ± 865.6 | 7764.8 a ± 1023.8 | 0.856 a ± 0.001 | 0.927 a ± 0.005 | 114.2 ± 8.2 |
| 500 | 2169.5 a ± 85.5 | 645.3 a ± 53.6 | 14,035.1 a ± 905.6 | 10,181.4 a ± 667.4 | 0.855 a ± 0.003 | 0.937 a ± 0.002 | 116.5 ± 8.9 |
| Limbarda crithmoides | ||||||||
| Ba (µM) | Fv/Fo | Fv/Fm | Fv’/Fm’ | ETR | ABS/RC | TRo/RC | ETo/RC | DIo/RC |
| 0 | 1.67 a ± 0.15 | 0.62 a ± 0.02 | 0.70 a ± 0.04 | 62.50 b ± 4.51 | 3.02 b ± 0.08 | 1.88 a ± 0.10 | 0.26 a ± 0.02 | 1.14 b ± 0.07 |
| 100 | 1.68 a ± 0.04 | 0.63 a ± 0.01 | 0.72 a ± 0.05 | 69.51 b ± 1.50 | 3.14 ab ± 0.09 | 1.97 a ± 0.04 | 0.26 a ± 0.03 | 1.17 ab ± 0.05 |
| 200 | 1.62 a ± 0.18 | 0.58 a ± 0.01 | 0.68 a ± 0.07 | 65.21 b ± 1.73 | 3.13 ab ± 0.11 | 1.96 a ± 0.03 | 0.26 a ± 0.03 | 1.17 ab ± 0.08 |
| 300 | 1.69 a ± 0.08 | 0.63 a ± 0.01 | 0.71 a ± 0.07 | 71.03 ab ± 4.70 | 3.29 ab ± 0.30 | 1.99 a ± 0.07 | 0.29 a ± 0.04 | 1.30 ab ± 0.07 |
| 500 | 1.66 a ± 0.05 | 0.61 a ± 0.03 | 0.70 a ± 0.06 | 73.10 a ± 2.38 | 3.60 a ± 0.05 | 2.02 a ± 0.01 | 0.29 a ± 0.02 | 1.53 a ± 0.05 |
| Helianthus annuus | ||||||||
| Ba (µM) | Fv/Fo | Fv/Fm | Fv’/Fm’ | ETR | ABS/RC | TRo/RC | ETo/RC | DIo/RC |
| 0 | 2.53 a ± 0.13 | 0.72 a ± 0.01 | 0.80 a ± 0.03 | 85.00 a ± 3.79 | 2.07 b ± 0.05 | 1.48 a ± 0.02 | 0.41 a ± 0.03 | 0.59 b ± 0.03 |
| 100 | 2.45 a ± 0.20 | 0.68 a ± 0.02 | 0.78 a ± 0.05 | 86.50 a ± 2.06 | 2.13 ab ± 0.10 | 1.58 a ± 0.04 | 0.40 a ± 0.02 | 0.55 ab ± 0.02 |
| 200 | 2.50 a ± 0.08 | 0.71 a ± 0.01 | 0.77 a ± 0.06 | 72.51 a ± 6.65 | 2.05 ab ± 0.08 | 1.46 a ± 0.05 | 0.36 ab ± 0.04 | 0.59 ab ± 0.04 |
| 300 | 2.66 a ± 0.08 | 0.73 a ± 0.01 | 0.79 a ±0.07 | 81.52 a ± 2.50 | 2.19 a ± 0.02 | 1.45 a ± 0.02 | 0.36 b ± 0.00 | 0.63 a ± 0.06 |
| 500 | 2.45 a ± 0.20 | 0.71 a ± 0.02 | 0.76 a ± 0.06 | 81.60 a ± 1.50 | 2.34 a ± 0.10 | 1.50 a ± 0.04 | 0.33 b ± 0.02 | 0.76 a ± 0.07 |
| Limbarda crithmoides | ||||||||
| Ba (µM) | K (mg·g−1 DW) | Ca (mg·g−1 DW) | Mg (mg·g−1 DW) | TIN (mg·g−1 DW) | ||||
| Shoots | Roots | Shoots | Roots | Shoots | Roots | Shoots | Roots | |
| 0 | 47.1 c ± 1.6 | 31.8 a ± 2.1 | 31.6 a ± 1.7 | 24.0 a ± 0.5 | 19.5 b ± 0.5 | 8.1 a ± 0.4 | 40.1 b ± 1.3 | 40.6 a ± 1.5 |
| 100 | 60.2 a ± 1.9 | 24.2 b ± 0.2 | 32.5 a ± 2.4 | 19.4 b ± 0.6 | 17.9 b ± 0.7 | 5.7 b ± 0.4 | 50.8 a ± 2.2 | 35.0 b ± 1.0 |
| 200 | 63.1 a ± 0.9 | 20.6 c ± 1.6 | 30.8 a ± 1.4 | 20.9 b ± 0.5 | 19.0 b ± 0.7 | 5.8 b ± 0.3 | 48.6 a ± 1.4 | 37.7 a ± 1.3 |
| 300 | 52.9 b ± 2.2 | 23.6 b ± 0.5 | 31.9 a ± 1.2 | 20.4 b ± 0.4 | 21.0 b ± 0.7 | 6.8 b ± 0.2 | 51.3 a ± 1.2 | 35.5 b ± 0.9 |
| 500 | 52.8 b ± 1.9 | 25.8 b ± 1.1 | 30.5 a ± 1.0 | 20.4 b ± 0.5 | 23.4 a ± 1.4 | 5.5 b ± 0.5 | 46.4 a ± 1.1 | 31.2 c ± 1.1 |
| Helianthus annuus | ||||||||
| Ba (µM) | K (mg·g−1 DW) | Ca (mg·g−1 DW) | Mg (mg·g−1 DW) | TIN (mg·g−1 DW) | ||||
| Shoots | Roots | Shoots | Roots | Shoots | Roots | Shoots | Roots | |
| 0 | 20.5 b ± 1.0 | 10.7 b ± 0.7 | 21.1 a ± 1.1 | 13.4 a ± 0.7 | 8.9 b ± 0.2 | 7.3 a ± 0.3 | 22.7 b ± 1.1 | 14.7 b ± 1.0 |
| 100 | 32.9 a ± 1.4 | 16.5 a ± 1.0 | 24.7 a ± 1.3 | 17.3 a ± 1.9 | 11.5 a ± 0.5 | 8.4 a ± 0.3 | 30.9 a ± 1.3 | 21.4 a ± 0.9 |
| 200 | 29.9 a ±1.4 | 14.6 a ± 1.1 | 23.7 a ± 1.1 | 14.6 a ± 1.7 | 11.1 a ± 0.5 | 7.4 a ± 0.8 | 37.8 a ± 2.0 | 31.9 a ± 1.8 |
| 300 | 30.1 a ± 1.9 | 10.4 b ± 1.1 | 22.3 a ± 0.7 | 17.5 a ± 1.4 | 10.3 b ± 0.5 | 7.0 a ± 0.3 | 30.2 a ± 1.2 | 15.0 b ± 0.7 |
| 500 | 24.4 b ± 1.0 | 8.9 b ± 0.8 | 21.5 a ± 0.8 | 17.6 a ± 0.7 | 9.0 b ± 0.4 | 6.8 a ± 0.2 | 19.4 b ± 0.6 | 13.0 b ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dridi, N.; Bouslimi, H.; Duarte, B.; Caçador, I.; Sleimi, N. Evaluation of Physiological and Biochemical Parameters and Some Bioindicators of Barium Tolerance in Limbarda crithmoides and Helianthus annuus. Int. J. Plant Biol. 2022, 13, 115-131. https://doi.org/10.3390/ijpb13020012
Dridi N, Bouslimi H, Duarte B, Caçador I, Sleimi N. Evaluation of Physiological and Biochemical Parameters and Some Bioindicators of Barium Tolerance in Limbarda crithmoides and Helianthus annuus. International Journal of Plant Biology. 2022; 13(2):115-131. https://doi.org/10.3390/ijpb13020012
Chicago/Turabian StyleDridi, Nesrine, Houda Bouslimi, Bernardo Duarte, Isabel Caçador, and Noomene Sleimi. 2022. "Evaluation of Physiological and Biochemical Parameters and Some Bioindicators of Barium Tolerance in Limbarda crithmoides and Helianthus annuus" International Journal of Plant Biology 13, no. 2: 115-131. https://doi.org/10.3390/ijpb13020012
APA StyleDridi, N., Bouslimi, H., Duarte, B., Caçador, I., & Sleimi, N. (2022). Evaluation of Physiological and Biochemical Parameters and Some Bioindicators of Barium Tolerance in Limbarda crithmoides and Helianthus annuus. International Journal of Plant Biology, 13(2), 115-131. https://doi.org/10.3390/ijpb13020012