
The Impact of Prenatal Alcohol Exposure on the Autonomic Nervous System and Cardiovascular System in Rats in a Sex-Specific Manner

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals
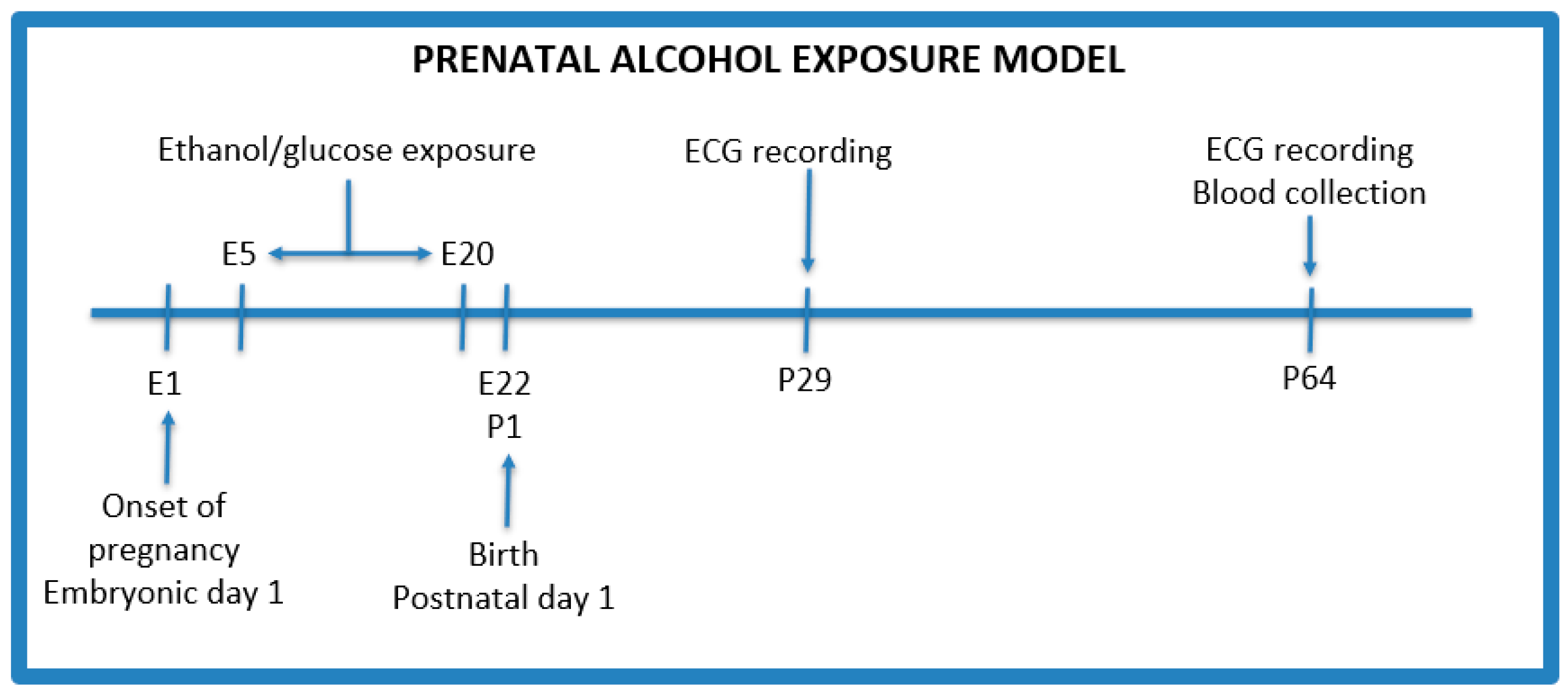
2.3. Model Protocol
- Control group (n = 2)—isocaloric glucose solution 40% (w/v) given by oral gavage administered every day between 5 and 20 gestational days;
- PAE group (n = 3)—3 g/kg b.w ethanol given by oral gavage administered every day between 5 and 20 gestational days with the following concentrations: 28.5% (v/v) ethanol from gestational day 5 to 17, and on the 18th, 19th, and 20th day, the dose of ethanol was reduced to 75%, 50%, and 25%, respectively, of the maximum dose to avoid withdrawal symptoms after the birth of the offspring.
2.4. Electrocardiographic Recordings
2.5. Biochemical Analysis
2.6. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, K.; Smith, D.; Ulleland, C.; Pytkowicz-Streissguth, A. Pattern of Malformaton in Offspring of Chronic Alcoholic Mothers. Lancet 1973, 1, 1267–1271. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.; Smith, D. Recognition of the Fetal Alcohol Syndrome in Early Infancy. Lancet 1973, 3, 999–1001. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.P.; Infante, M.A.; Warren, K.R. Fetal Alcohol Spectrum Disorders: An Overview. Neuropsychol. Rev. 2011, 21, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, D.Y.; Buckley, D.D.M.; Eberhart, D.J.K. Diving into the World of Alcohol Teratogenesis: A Review of Zebrafish Models of Fetal Alcohol Spectrum Disorders. Biochem. Cell Biol. 2018, 96, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Cudd, T.A. Focus On: The Use of Animal Models for the Study of Fetal Alcohol Spectrum Disorders. Alcohol. Res. Health 2011, 34, 92–98. [Google Scholar] [PubMed]
- Petrelli, B.; Weinberg, J.; Hicks, G.G. Effects of Prenatal Alcohol Exposure (PAE): Insights into FASD Using PAE Mouse Models. Biochem. Cell Biol. 2018, 96, 131–147. [Google Scholar] [CrossRef]
- Patten, A.R.; Fontaine, C.J.; Christie, B.R. A Comparison of the Different Animal Models of Fetal Alcohol Spectrum Disorders and Their Use in Studying Complex Behaviors. Front. Pediatr. 2014, 2, 93. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S.; DeCarli, L.M. The Feeding of Alcohol in Liquid Diets: Two Decades of Applications and 1982 Update. Alcohol. Clin. Exp. Res. 1982, 6, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Allan, A.M.; Chynoweth, J.; Tyler, L.A.; Caldwell, K.K. A Mouse Model of Prenatal Ethanol Exposure Using a Voluntary Drinking Paradigm. Alcohol. Clin. Exp. Res. 2003, 27, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Dunty, W.C.; Chen, S.; Zucker, R.M.; Dehart, D.B.; Sulik, K.K. Selective Vulnerability of Embryonic Cell Populations to Ethanol-Induced Apoptosis: Implications for Alcohol-Related Birth Defects and Neurodevelopmental Disorder. Alcohol. Clin. Exp. Res. 2001, 25, 1523–1535. [Google Scholar] [CrossRef] [PubMed]
- Karanian, J.; Yergey, J.; Lister, R.; D’Souza, N.; Linnoila, M.; Salem, N. Characterization of an Automated Apparatus for Precise Control of Inhalation Chamber Ethanol Vapor and Blood Ethanol Concentrations. Alcohol. Clin. Exp. Res. 1986, 10, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Wiener, S.G.; Bloom, F.E. Long-Term Ethanol Administration Methods for Rats: Advantages of Inhalation over Intubation or Liquid Diets. Behav. Neural Biol. 1979, 27, 466–486. [Google Scholar] [CrossRef] [PubMed]
- Popova, S.; Dozet, D.; Shield, K.; Rehm, J.; Burd, L. Alcohol’s Impact on the Fetus. Nutrients 2021, 13, 3452. [Google Scholar] [CrossRef] [PubMed]
- Waddell, J.; Hill, E.; Tang, S.; Jiang, L.; Xu, S.; Mooney, S.M. Choline Plus Working Memory Training Improves Prenatal Alcohol-Induced Deficits in Cognitive Flexibility and Functional Connectivity in Adulthood in Rats. Nutrients 2020, 12, 3513. [Google Scholar] [CrossRef] [PubMed]
- van Ravenswaaij-Arts, C.; Kollee, L.; Hopman, J.; Stoelinga, G.; van Geijn, H. Heart Rate Variability. Ann. Intern. Med. 1993, 118, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Atum, A.L.B.; De Matos, L.P.; De Jesus, B.C.; Nasuk, G.R.; Da Silva, G.A.; Gomes, C.P.; Pesquero, J.B.; Zamuner, S.R.; Silva Júnior, J.A. Impact of Prenatal Alcohol Exposure on the Development and Myocardium of Adult Mice: Morphometric Changes, Transcriptional Modulation of Genes Related to Cardiac Dysfunction, and Antioxidant Cardioprotection. Antioxidants 2023, 12, 256. [Google Scholar] [CrossRef] [PubMed]
- Parkington, H.C.; Coleman, H.A.; Wintour, E.M.; Tare, M. Prenatal Alcohol Exposure: Implications for Cardiovascular Function in the Fetus and Beyond. Clin. Exp. Pharmacol. Physiol. 2010, 37, 91–98. [Google Scholar] [CrossRef]
- Jurczyk, M.; Dyląg, K.A.; Skowron, K.; Gil, K. Prenatal Alcohol Exposure and Autonomic Nervous System Dysfunction: A Review Article. Folia Med. Cracov. 2019, 59, 15–21. [Google Scholar] [CrossRef]
- Thomas, J.D.; Abou, E.J.; Dominguez, H.D. Prenatal Choline Supplementation Mitigates the Adverse Effects of Prenatal Alcohol Exposure on Development in Rats. Neurotoxicol. Teratol. 2009, 31, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Martin, C.D.; Lei, A.L.; Hausknecht, K.A.; Turk, M.; Micov, V.; Kwarteng, F.; Ishiwari, K.; Oubraim, S.; Wang, A.; et al. Prenatal Ethanol Exposure Impairs Sensory Processing and Habituation of Visual Stimuli, Effects Normalized by Postnatal Environmental Enrichment. Alcohol. Clin. Exp. Res. 2022, 46, 891–906. [Google Scholar] [CrossRef] [PubMed]
- Camm, A.; Malik, M.; Bigger, J.; Breithardt, G.; Cerutti, S.; Cohen, R.; Coumel, P.; Fallen, E.; Kennedy, H.; Kleiger, R.; et al. Task Force of The European Society of Cardiology and The North American Society of Pacing and Electrophysiology Heart Rate Variability Standards of Measurement, Physiological Interpretation, and Clinical Use. Eur. Heart J. 1996, 17, 354–381. [Google Scholar]
- Dobrek, Ł.; Baranowska, A.; Thor, P.J. Heart Rate Variability after BRL37344, a Beta-3 Agonist, in Experimental Bladder Outlet Obstruction. Postępy Hig. Med. Dośw. 2013, 67, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-T.; Huang, W.-C.; Cheng, C.-C.; Ke, M.-W.; Tsai, J.-S.; Hung, Y.-M.; Huang, N.-C.; Huang, M.-S.; Wann, S.-R. Effects of Epinephrine on Heart Rate Variability and Cytokines in a Rat Sepsis Model. Bosn. J. Basic Med. Sci. 2018, 20, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.S.; DeMarco, V.G.; Heesch, C.M.; Whaley-Connell, A.T.; Schneider, R.I.; Rehmer, N.T.; Tilmon, R.D.; Ferrario, C.M.; Sowers, J.R. Sex Differences in Baroreflex Sensitivity, Heart Rate Variability, and End Organ Damage in the TGR(mRen2)27 Rat. Am. J. Physiol.-Heart Circ. Physiol. 2011, 301, H1540–H1550. [Google Scholar] [CrossRef] [PubMed]
- Fares, S.A.; Habib, J.R.; Engoren, M.C.; Badr, K.F.; Habib, R.H. Effect of Salt Intake on Beat-to-Beat Blood Pressure Nonlinear Dynamics and Entropy in Salt-Sensitive versus Salt-Protected Rats. Physiol. Rep. 2016, 4, e12823. [Google Scholar] [CrossRef] [PubMed]
- Willson, K.; Francis, D.P.; Wensel, R.; Coats, A.J.S.; Parker, K.H. Relationship between Detrended Fluctuation Analysis and Spectral Analysis of Heart-Rate Variability. Physiol. Meas. 2002, 23, 385–401. [Google Scholar] [CrossRef] [PubMed]
- Beckers, F.; Verheyden, B.; Aubert, A.E. Aging and Nonlinear Heart Rate Control in a Healthy Population. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H2560–H2570. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.J.; Richards, J.E. Development of Heart Inter-Beat Interval Variability in Preweanling Rats: Effects of Exposure to Alcohol and Hypoxia. Physiol. Behav. 1997, 61, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, F.; Ginsberg, J.P. An Overview of Heart Rate Variability Metrics and Norms. Front. Public Health 2017, 5, 258. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, S.; Guo, L.; Peng, X.; Liu, Y. Prenatal Alcohol Exposure Induced Congenital Heart Diseases: From Bench to Bedside. Birth Defects Res. 2021, 113, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Zhu, J.; Lv, T.; Sun, H.; Huang, X.; Tian, J. Alcohol Consumption During Gestation Causes Histone3 Lysine9 Hyperacetylation and an Alternation of Expression of Heart Development-Related Genes in Mice. Alcohol. Clin. Exp. Res. 2014, 38, 2396–2402. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, S.; Muralidharan, P.; Marrs, J.A. Embryonic Ethanol Exposure Dysregulates BMP and Notch Signaling, Leading to Persistent Atrio-Ventricular Valve Defects in Zebrafish. PLoS ONE 2016, 11, e0161205. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.-Q.; Yasmin, N.; Collier, A.D.; Karatayev, O.; Khalizova, N.; Onoichenco, A.; Fam, M.; Albeg, A.S.; Campbell, S.; Leibowitz, S.F. Fibroblast Growth Factor 2: Role in Prenatal Alcohol-Induced Stimulation of Hypothalamic Peptide Neurons. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 116, 110536. [Google Scholar] [CrossRef] [PubMed]
- Collier, A.D.; Yasmin, N.; Chang, G.; Karatayev, O.; Khalizova, N.; Fam, M.; Abdulai, A.R.; Yu, B.; Leibowitz, S.F. Embryonic Ethanol Exposure Induces Ectopic Hcrt and MCH Neurons Outside Hypothalamus in Rats and Zebrafish: Role in Ethanol-induced Behavioural Disturbances. Addict. Biol. 2022, 27, e13238. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, L.S.; Schwarz, J.M. Effects of Moderate Prenatal Alcohol Exposure during Early Gestation in Rats on Inflammation across the Maternal-Fetal-Immune Interface and Later-Life Immune Function in the Offspring. J. Neuroimmune Pharmacol. 2016, 11, 680–692. [Google Scholar] [CrossRef]
- Burgess, D.J.; Dorey, E.S.; Gardebjer, E.M.; Bielefeldt-Ohmann, H.; Moritz, K.M.; Cuffe, J.S.M. Periconceptional Ethanol Exposure Alters the Stress Axis in Adult Female but Not Male Rat Offspring. Stress 2019, 22, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Walton, S.L.; Tjongue, M.; Tare, M.; Kwok, E.; Probyn, M.; Parkington, H.C.; Bertram, J.F.; Moritz, K.M.; Denton, K.M. Chronic Low Alcohol Intake during Pregnancy Programs Sex-Specific Cardiovascular Deficits in Rats. Biol. Sex Differ. 2019, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kim, J.Y.; Lee, D.Y.; Park, K.J.; Kim, G.H.; Kim, J.E.; Roh, G.S.; Lim, J.Y.; Koo, S.; Lim, N.K.; et al. Alcohol Consumption before Pregnancy Causes Detrimental Fetal Development and Maternal Metabolic Disorders. Sci. Rep. 2020, 10, 10054. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Gao, F.; Liu, X.; Li, J.; Wang, Y.; Han, J.; Wang, X. Prenatal Alcohol Exposure and Offspring Liver Dysfunction: A Systematic Review and Meta-Analysis. Arch. Gynecol. Obstet. 2016, 294, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Fofana, B.; Yao, X.; Rampitsch, C.; Cloutier, S.; Wilkins, J.A.; Nyomba, B.L.G. Prenatal Alcohol Exposure Alters Phosphorylation and Glycosylation of Proteins in Rat Offspring Liver. Proteomics 2010, 10, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.-H.; Chen, L.; Nyomba, B.L.G. Adult Rats Prenatally Exposed to Ethanol Have Increased Gluconeogenesis and Impaired Insulin Response of Hepatic Gluconeogenic Genes. J. Appl. Physiol. 2006, 100, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Addolorato, G.; Gasbarrini, A.; Marcoccia, S.; Simoncini, M.; Baccarini, P.; Vagni, G.; Grieco, A.; Sbriccoli, A.; Granato, A.; Stefanini, G.F.; et al. Prenatal Exposure to Ethanol in Rats: Effects on Liver Energy Level and Antioxidant Status in Mothers, Fetuses, and Newborns. Alcohol 1997, 14, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Akison, L.K.; Probyn, M.E.; Gray, S.P.; Cullen-McEwen, L.A.; Tep, K.; Steane, S.E.; Gobe, G.C.; Wlodek, M.E.; Bertram, J.F.; Moritz, K.M. Moderate Prenatal Ethanol Exposure in the Rat Promotes Kidney Cell Apoptosis, Nephron Deficits, and Sex-specific Kidney Dysfunction in Adult Offspring. Anat. Rec. 2020, 303, 2632–2645. [Google Scholar] [CrossRef] [PubMed]
- Dyląg, K.A.; Dumnicka, P.; Kowalska, K.; Migas-Majoch, A.; Przybyszewska, K.; Drożdż, D. Increased Incidence of Renal and Urinary Tract Anomalies among Individuals with Fetal Alcohol Spectrum Disorders (FASD). Birth Defects Res. 2023, 116, e2259. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.P.; Denton, K.M.; Cullen-McEwen, L.; Bertram, J.F.; Moritz, K.M. Prenatal Exposure to Alcohol Reduces Nephron Number and Raises Blood Pressure in Progeny. J. Am. Soc. Nephrol. 2010, 21, 1891–1902. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, J.; Sliwowska, J.H.; Lan, N.; Hellemans, K.G.C. Prenatal Alcohol Exposure: Foetal Programming, the Hypothalamic-Pituitary-Adrenal Axis and Sex Differences in Outcome. J. Neuroendocrinol. 2008, 20, 470–488. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zuo, N.; Li, B.; Xiong, Y.; Chen, H.; He, H.; Sun, Z.; Hu, S.; Cheng, H.; Ao, Y.; et al. The Expressional Disorder of the Renal RAS Mediates Nephrotic Syndrome of Male Rat Offspring Induced by Prenatal Ethanol Exposure. Toxicology 2018, 400–401, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Qin, J.; Zhou, J.; Magdalou, J.; Chen, L.; Xu, D.; Wang, H. Glucocorticoid Programming Mechanism for Hypercholesterolemia in Prenatal Ethanol-Exposed Adult Offspring Rats. Toxicol. Appl. Pharmacol. 2019, 375, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Hannich, M.; Wallaschofski, H.; Nauck, M.; Reincke, M.; Adolf, C.; Völzke, H.; Rettig, R.; Hannemann, A. Physiological Aldosterone Concentrations Are Associated with Alterations of Lipid Metabolism: Observations from the General Population. Int. J. Endocrinol. 2018, 2018, 4128174. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Control | PAE | p Value | |
|---|---|---|---|
| AST [IU] | 150.38 ± 6.49 | 151.87 ± 5.62 | NS |
| ALT [IU] | 46.88 ± 1.57 | 51.20 ± 1.12 * | 0.0350 |
| Na [mmol/L] | 144.00 ± 0.42 | 144.87 ± 0.34 | NS |
| K [mmol/L] | 5.6 ± 0.13 | 5.9 ± 0.11 | NS |
| Mg [mmol/L] | 1.01 ± 0.02 | 0.99 ± 0.01 | NS |
| Creat [mmol/L] | 29.99 ± 1.02 | 33.47 ± 0.67 * | 0.0079 |
| Chol [mmol/L] | 1.56 ± 0.07 | 1.70 ± 0.04 | NS |
| HDL [mmol/L] | 1.00 ± 0.05 | 1.07 ± 0.03 | NS |
| Non-HDL [mmol/L] | 0.57 ± 0.02 | 0.62 ± 0.02 | NS |
| LDL [mmol/L] | 0.18 ± 0.02 | 0.21 ± 0.1 | NS |
| TG [mmol/L] | 1.66 ± 0.10 | 1.52 ± 0.07 | NS |
| Control | PAE | ||||
|---|---|---|---|---|---|
| F | M | F | M | p Value | |
| AST [IU] | 151.80 ± 21.28 | 148.00 ± 16.09 | 142.75 ± 16.10 | 162.29 ± 23.80 | NS |
| ALT [IU] | 46.60 ± 5.86 | 47.33 ± 0.58 | 50.63 ± 3.58 | 51.86 ± 5.30 | NS |
| Na [mmol/L] | 144.00 ± 1.41 | 144.00 ± 1.00 | 144.50 ± 0.93 | 145.29 ± 1.60 | NS |
| K [mmol/L] | 5.30 ± 0.11 | 5.99 ± 0.22 | 5.24 ± 0.24 | 5.77 ± 0.39 | NS |
| Mg [mmol/L] | 0.98 ± 0.05 | 1.05 ± 0.03 | 0.99 ± 0.04 | 0.99 ± 0.05 | NS |
| Creat [mmol/L] | 28.92 ± 2.83 | 31.77 ± 2.37 | 34.39 ± 0.97 * | 32.41 ± 3.51 | 0.0081 |
| Chol [mmol/L] | 1.44 ± 0.11 | 1.77 ± 0.12 | 1.63 ± 0.17 | 1.79 ± 0.07 | NS |
| HDL [mmol/L] | 0.91 ± 0.10 | 1.14 ± 0.06 | 1.05 ± 0.14 | 1.10 ± 0.06 | NS |
| Non-HDL [mmol/L] | 0.52 ± 0.02 | 0.65 ± 0.03 | 0.57 ± 0.06 | 0.67 ± 0.06 | NS |
| LDL [mmol/L] | 0.13 ± 0.01 | 0.26 ± 0.00 | 0.17 ± 0.04 | 0.26 ± 0.03 | NS |
| TG [mmol/L] | 1.78 ± 0.26 | 1.46 ± 0.26 | 1.45 ± 0.27 | 1.60 ± 0.31 | NS |
| Day 29 | Day 64 | ||||
|---|---|---|---|---|---|
| Control | PAE | Control | PAE | p Value | |
| Weight [g] | 76.5 ± 1.89 | 79.06 ± 1.70 | 221.22 ± 19.92 | 234.93 ± 14.62 | NS |
| Mean HR [beats/min] | 388.57 ± 16.28 | 343.55 ± 17.52 | 308.24 ± 10.32 | 300.69 ± 14.62 | NS |
| SDNN [ms] | 1.84 ± 0.53 | 3.69 ± 0.91 | 2.12 ± 0.15 | 2.11 ± 0.33 | NS |
| RMSSD [ms] | 2.05 ± 0.27 | 5.21 ± 1.14 * | 2.87 ± 0.31 | 3.01 ± 0.40 | 0.0092 |
| LF n.u. | 10.88 ± 2.81 | 5.72 ± 1.00 | 17.32 ± 6.64 | 12.48 ± 3.32 | NS |
| HF n.u. | 89.11 ± 2.81 | 94.27 ± 1.00 | 82.68 ± 6.64 | 87.52 ± 3.32 | NS |
| LF/HF | 0.13 ± 0.04 | 0.06 ± 0.01 | 0.31 ± 0.18 | 0.17 ± 0.06 | NS |
| SD1 [ms] | 1.45 ± 0.19 | 3.69 ± 0.81 * | 2.03 ± 0.22 | 2.13 ± 0.28 | 0.0093 |
| SD2 [ms] | 2.04 ± 0.77 | 3.63 ± 1.01 | 2.11 ± 0.23 | 2.05 ± 0.39 | NS |
| SD2/SD1 | 1.30 ± 0.28 | 0.97 ± 0.07 | 1.21 ± 0.31 | 0.94 ± 0.06 | NS |
| ApEn | 1.01 ± 0.05 | 0.99 ± 0.05 | 1.13 ± 0.02 | 1.12 ± 0.02 | NS |
| SampEn | 1.31 ± 0.13 | 1.17 ± 0.08 | 1.65 ± 0.12 | 1.55 ± 0.07 | NS |
| DFA1 | 0.36 ± 0.05 | 0.24 ± 0.02 | 0.43 ± 0.11 | 0.35 ± 0.05 | NS |
| DFA2 | 0.59 ± 0.11 | 0.45 ± 0.08 | 0.71 ± 0.11 | 0.65 ± 0.04 | NS |
| Day 29 | Day 64 | |||||||
|---|---|---|---|---|---|---|---|---|
| Control | PAE | Control | PAE | |||||
| F | M | F | M | F | M | F | M | |
| Weight [g] | 71.6 ± 1.03 | 81.40 ± 1.72 | 74.25 ± 1.56 | 83.88 ± 1.82 | 171.20 ± 2.96 | 283.75 ± 4.44 | 183.29 ± 2.15 | 286.57 ± 5.76 |
| Mean HR [beats/min] | 381.37 ± 25.7 | 397.57 ± 21.0 | 301.21 ± 13.2 | 385.90 ± 21.4 | 301.57 ± 14.3 | 319.35 ± 14.6 | 296.33 ± 10.0 | 304.43 ± 7.8 |
| SDNN [ms] | 2.30 ± 0.94 | 1.26 ± 0.17 | 4.88 ± 1.70 | 2.49 ± 0.40 | 2.01 ± 0.18 | 2.30 ± 0.25 | 1.89 ± 0.33 | 2.29 ± 0.57 |
| RMSSD [ms] | 2.31 ± 0.41 | 1.72 ± 0.32 | 6.77 ± 2.08 | 3.64 ± 0.66 | 2.95 ± 0.34 | 2.74 ± 0.72 | 2.65 ± 0.49 | 3.33 ± 0.62 |
| LF n.u. | 9.95 ± 4.74 | 12.06 ± 3.00 | 5.04 ± 1.48 | 6.40 ± 1.41 | 12.76 ± 3.85 | 24.92 ± 17.77 | 11.59 ± 4.68 | 13.25 ± 5.02 |
| HF n.u. | 90.05 ± 4.74 | 87.94 ± 3.00 | 94.96 ± 1.48 | 93.58 ± 1.41 | 87.24 ± 3.85 | 75.08 ± 17.77 | 88.41 ± 4.68 | 86.75 ± 5.02 |
| LF/HF | 0.12 ± 0.07 | 0.14 ± 0.04 | 0.05 ± 0.02 | 0.07 ± 0.02 | 0.16 ± 0.05 | 0.56 ± 0.48 | 0.15 ± 0.08 | 0.18 ± 0.09 |
| SD1 [ms] | 1.63 ± 0.29 | 1.22 ± 0.23 | 4.80 ± 1.47 | 2.58 ± 0.46 | 2.09 ± 0.24 | 1.94 ± 0.51 | 1.88 ± 0.34 | 2.36 ± 0.44 |
| SD2 [ms] | 2.64 ± 1.38 | 1.30 ± 0.11 | 4.90 ± 1.93 | 2.35 ± 0.37 | 1.90 ± 0.18 | 2.45 ± 0.53 | 1.89 ± 0.34 | 2.18 ± 0.69 |
| SD2/ SD1 | 1.45 ± 0.52 | 1.12 ± 0.10 | 0.97 ± 0.10 | 0.97 ± 0.10 | 0.95 ± 0.12 | 1.64 ± 0.82 | 1.04 ± 0.08 | 0.85 ± 0.08 |
| ApEn | 0.96 ± 0.06 | 1.06 ± 0.08 | 1.00 ± 0.06 | 0.98 ± 0.08 | 1.16 ± 0.03 | 1.09 ± 0.02 | 1.14 ± 0.04 | 1.11 ± 0.03 |
| SampEn | 1.31 ± 0.22 | 1.31 ± 0.14 | 1.19 ± 0.10 | 1.16 ± 0.13 | 1.76 ± 0.14 | 1.45 ± 0.18 | 1.67 ± 0.09 | 1.46 ± 0.09 |
| DFA1 | 0.36 ± 0.09 | 0.35 ± 0.05 | 0.23 ± 0.04 | 0.26 ± 0.03 | 0.36 ± 0.06 | 0.54 ± 0.30 | 0.38 ± 0.07 | 0.32 ± 0.07 |
| DFA2 | 0.63 ± 0.19 | 0.54 ± 0.06 | 0.44 ± 0.10 | 0.45 ± 0.13 | 0.66 ± 0.14 | 0.78 ± 0.19 | 0.75 ± 0.07 | 0.57 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurczyk, M.; Król, M.; Midro, A.; Dyląg, K.; Kurnik-Łucka, M.; Skowron, K.; Gil, K. The Impact of Prenatal Alcohol Exposure on the Autonomic Nervous System and Cardiovascular System in Rats in a Sex-Specific Manner. Pediatr. Rep. 2024, 16, 278-287. https://doi.org/10.3390/pediatric16020024
Jurczyk M, Król M, Midro A, Dyląg K, Kurnik-Łucka M, Skowron K, Gil K. The Impact of Prenatal Alcohol Exposure on the Autonomic Nervous System and Cardiovascular System in Rats in a Sex-Specific Manner. Pediatric Reports. 2024; 16(2):278-287. https://doi.org/10.3390/pediatric16020024
Chicago/Turabian StyleJurczyk, Michał, Magdalena Król, Aleksandra Midro, Katarzyna Dyląg, Magdalena Kurnik-Łucka, Kamil Skowron, and Krzysztof Gil. 2024. "The Impact of Prenatal Alcohol Exposure on the Autonomic Nervous System and Cardiovascular System in Rats in a Sex-Specific Manner" Pediatric Reports 16, no. 2: 278-287. https://doi.org/10.3390/pediatric16020024
APA StyleJurczyk, M., Król, M., Midro, A., Dyląg, K., Kurnik-Łucka, M., Skowron, K., & Gil, K. (2024). The Impact of Prenatal Alcohol Exposure on the Autonomic Nervous System and Cardiovascular System in Rats in a Sex-Specific Manner. Pediatric Reports, 16(2), 278-287. https://doi.org/10.3390/pediatric16020024