
Acaricidal Activity of Biosurfactants Produced by Serratia ureilytica on Tetranychus urticae and Their Compatibility with the Predatory Mite Amblyseius swirskii

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Production of Biosurfactants
2.3. Characterization of Biosurfactants
2.4. Colony Establishment of T. urticae
2.5. Preparation of Biosurfactants for Bioassays
2.6. Evaluation of Mortality of T. urticae and A. swirskii in the Laboratory
2.7. Evaluation of Egg Mortality of T. urticae in the Laboratory
2.8. Evaluation of Population Suppression of T. urticae in Greenhouse
2.9. Bioinformatic Analysis of Genes Related to Biosurfactant Biosynthesis
2.10. Statistical Analysis
3. Results
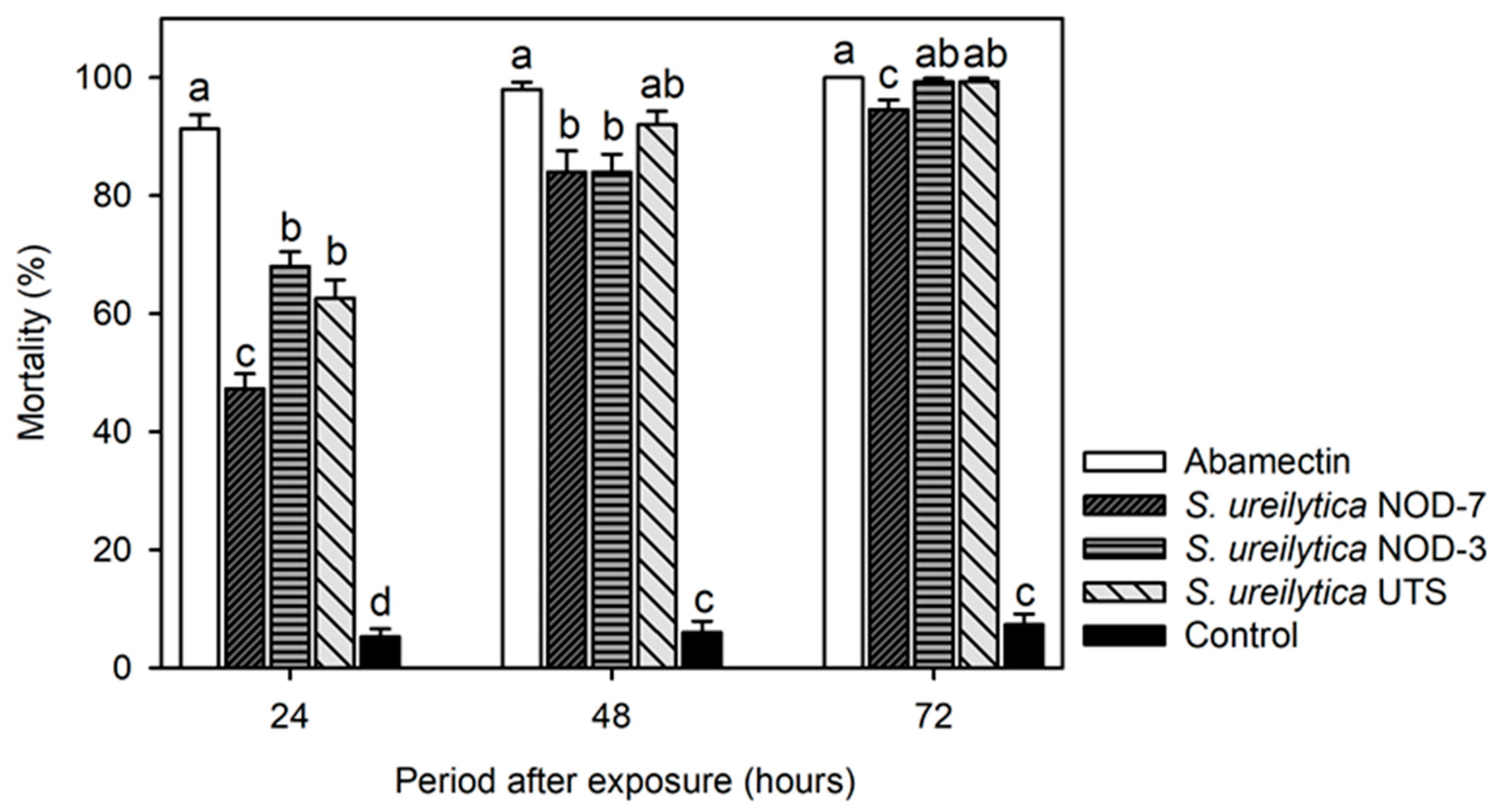
3.1. Toxicity of Biosurfactants in T. urticae Adults and Eggs in the Laboratory
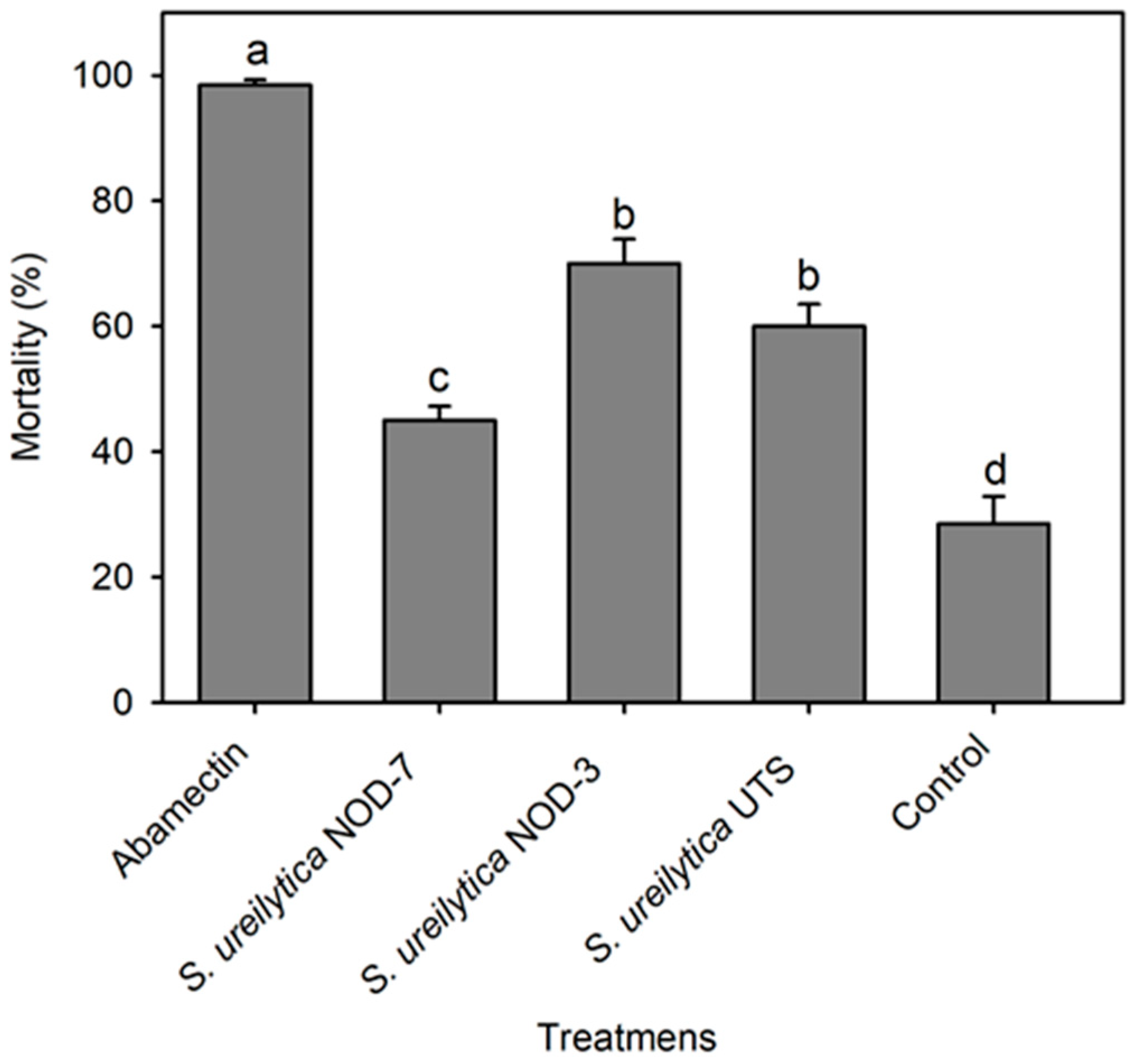
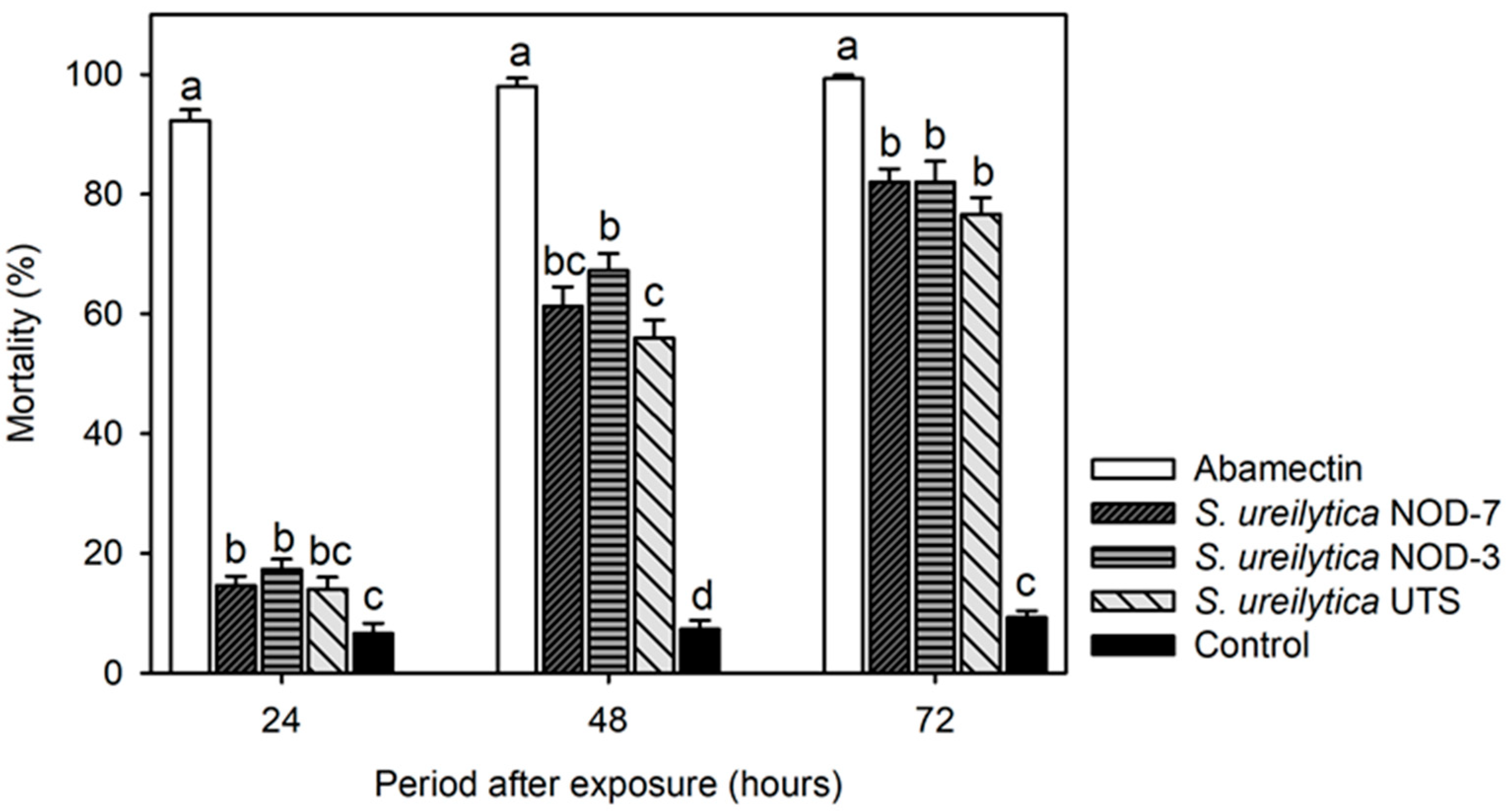
3.2. Toxicity of Biosurfactants in Predatory Mite Amblyseius swirskii in the Laboratory
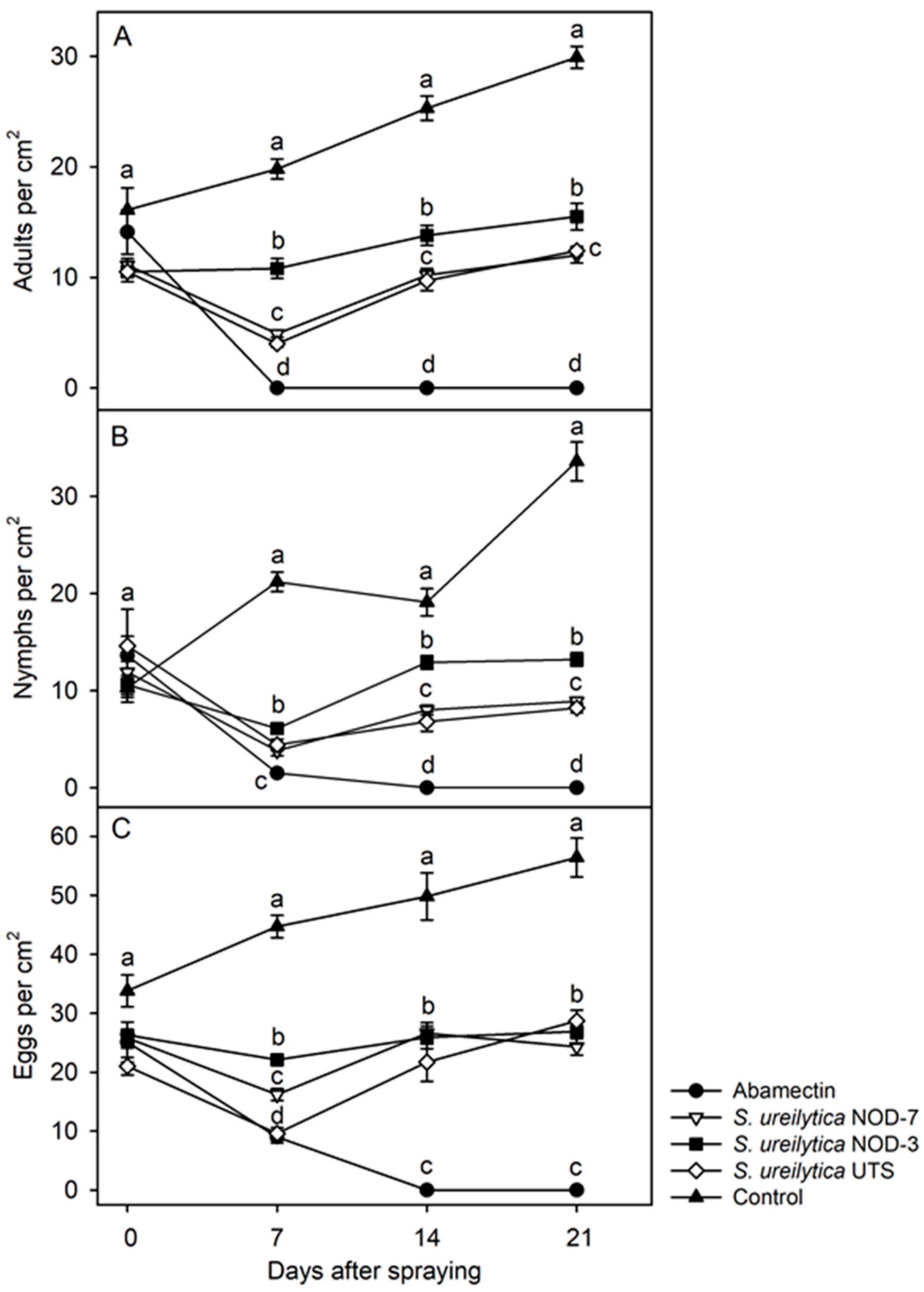
3.3. Suppression of T. urticae Population Density in Greenhouse
3.4. Analysis of Biosurfactants by GC-MS
3.5. Identification of Metabolites in S. ureilytica Genomes by Bioinformatics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kshetri, L.; Naseem, F.; Pandey, P. Role of Serratia sp. as Biocontrol Agent and Plant Growth Stimulator, with Prospects of Biotic Stress Management in Plant. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management Microorganisms for Sustainability; Sayyed, R., Ed.; Springer: Singapore, 2019; Volume 13. [Google Scholar] [CrossRef]
- Allen, J.L.; Doidge, N.P.; Cheng, C.; Lynch, M.; Crabb, H.K.; Scheerlinck, J.-P.; Bushell, R.; Browning, G.F.; Marenda, M.S.; Kuo, C.-H. Genomic characterisation of an entomopathogenic strain of Serratia ureilytica in the critically endangered phasmid Dryococelus australis. PLoS ONE 2022, 17, e0265967. [Google Scholar] [CrossRef]
- Moreno-Castillo, B.; Ortiz-Barrios, H.; Salvador-Figueroa, M.; Adriano-Anaya, M.L. Serratia marcescens (Enterobacteriaceae): Promotora de la salud vegetal. Biociencias 2022, 5, 1–9. [Google Scholar]
- Bidari, F.; Shams-Bakhsh, M.; Mehrabadi, M. Isolation and characterization of a Serratia marcescens with insecticidal activity from Polyphylla olivieri (Col.: Scarabaeidae). J. Appl. Èntomol. 2017, 142, 162–172. [Google Scholar] [CrossRef]
- Ruiu, L.; Virdis, B.; Mura, M.E.; Floris, I.; Satta, A.; Tarasco, E. Oral insecticidal activity of new bacterial isolates against insects in two orders. Biocontrol Sci. Technol. 2017, 27, 886–902. [Google Scholar] [CrossRef]
- Fu, R.; Zhou, L.; Feng, K.; Lu, X.; Luo, J.; Tang, F. Effects of Serratia marcescens (SM1) and its interaction with common biocontrol agents on the termite, Odontotermes formosanus (Shiraki). J. For. Res. 2021, 32, 1263–1267. [Google Scholar] [CrossRef]
- Niu, H.; Sun, Y.; Zhang, Z.; Zhao, D.; Wang, N.; Wang, L.; Guo, H. The endophytic bacterial entomopathogen Serratia marcescens promotes plant growth and improves resistance against Nilaparvata lugens in rice. Microbiol. Res. 2022, 256, 126956. [Google Scholar] [CrossRef]
- Mai, A.-G.M. Serratia A Novel Source of Secondary Metabolites. Adv. Biotechnol. Microbiol. 2018, 11, 555814. [Google Scholar] [CrossRef]
- Cao, X.; Ye, Y.; Li, P. Genome Sequence Resource of Serratia ureilytica HNU47: A Strain with Biocontrol Potential Against Bacterial Wilt Pathogen Ralstonia solanacearum. Plant Dis. 2023, 107, 919–921. [Google Scholar] [CrossRef]
- Zamorano-González, C.A.; Ramírez-Trujillo, J.A.; Pilotzi-Xahuentitla, H.; Yáñez-Ocampo, G.; Hernández-Nuñéz, E.; Suárez-Rodríguez, R.; Orea-Flores, M.L.A.; Gómez-Rodríguez, O.; Espinosa-Zaragoza, S.; Rangel-Zaragoza, J.L.; et al. In Vitro Evaluation of the Biosurfactant Produced by Serratia ureilytica UTS with Antifungal and Nematicidal Activity Against Nacobbus aberrans. Curr. Microbiol. 2025, 82, 63. [Google Scholar] [CrossRef]
- Espinosa-Zaragoza, S.; Domínguez-Liévano, A.; Gómez-Gutiérrez, J.A.; Wong-Villarreal, A.; Aguilar-Marcelino, L.; Cerqueda-García, D.; Rangel-Zaragoza, J.L.; Sanzón-Gómez, D.; Mireles-Arriaga, A.I.; Sachman-Ruíz, B. In vitro Acaricidal Activity of Serratia Ureilytica Against the Dust Mite Tyrophagus Putrescentiae and Identification of Genes Related to Biocontrol. Curr. Microbiol. 2024, 81, 199. [Google Scholar] [CrossRef]
- Wong-Villarreal, A.; Méndez-Santiago, E.W.; Gómez-Rodríguez, O.; Aguilar-Marcelino, L.; García, D.C.; García-Maldonado, J.Q.; Hernández-Velázquez, V.M.; Yañez-Ocampo, G.; Espinosa-Zaragoza, S.; Ramírez-González, S.I.; et al. Nematicidal Activity of the Endophyte Serratia ureilytica against Nacobbus aberrans in Chili Plants (Capsicum annuum L.) and Identification of Genes Related to Biological Control. Plants 2021, 10, 2655. [Google Scholar] [CrossRef]
- Kumari, S.; Chauhan, U.; Kumari, A.; Nadda, G. Comparative toxicities of novel and conventional acaricides against different stages of Tetranychus urticae Koch (Acarina: Tetranychidae). J. Saudi Soc. Agric. Sci. 2017, 16, 191–196. [Google Scholar] [CrossRef]
- Adesanya, A.W.; Beauchamp, M.J.; Lavine, M.D.; Lavine, L.C.; Zhu, F.; Walsh, D.B. Physiological resistance alters behavioral response of Tetranychus urticae to acaricides. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- A Schmidt-Jeffris, R.; Coffey, J.L.; Miller, G.; A Farfan, M.; Riddick, E. Residual Activity of Acaricides for Controlling Spider Mites in Watermelon and Their Impacts on Resident Predatory Mites. J. Econ. Èntomol. 2021, 114, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Uddin, N.; Alam, Z.; Miah, R.U.; Mian, I.H.; Mustarin, K.-E. Toxicity of pesticides to Tetranychus urticae Koch (Acari:Tetranychidae) and their side effects on Neoseiulus californicus (Acari:Phytoseiidae). Int. J. Acarol. 2015, 41, 688–693. [Google Scholar] [CrossRef]
- Montoya, A.; Galano-Flores, G.; Rodríguez, H.; Franco, A.A.; Zardi, O.Z.; Yamamoto, P.T. Toxicity of acaricides on Tetranychus urticae (Koch) in the laboratory. Rev. Prot. Veg. 2017, 32, 60–67. [Google Scholar]
- Zhao, J.; Wang, Z.; Ji, M.; Cheng, J.; Zhu, G.; Yu, C. Synthesis and bioactivity evaluation of novel spiromesifen derivatives. Pest Manag. Sci. 2011, 68, 10–15. [Google Scholar] [CrossRef]
- Tu, S.; Qiu, X.; Cao, L.; Han, R.; Zhang, Y.; Liu, X. Expression and characterization of the chitinases from Serratia marcescens GEI strain for the control of Varroa destructor, a honey bee parasite. J. Invertebr. Pathol. 2010, 104, 75–82. [Google Scholar] [CrossRef]
- Pritam, C.; Sukanta, K.S. Systemic infestation of Serratia entomophila AB2 through plant tissue inferred protection against insect pest and fungal pathogens. Afr. J. Microbiol. Res. 2013, 7, 2651–2655. [Google Scholar] [CrossRef]
- Ruiu, L. Insect Pathogenic Bacteria in Integrated Pest Management. Insects 2015, 6, 352–367. [Google Scholar] [CrossRef]
- Sanchéz-Cruz, R.; Vázquez, I.T.; Batista-García, R.A.; Méndez-Santiago, E.W.; del Rayo, S.C.; Leija, A.; Lira-Ruan, V.; Hernández, G.; Wong-Villarreal, A.; Folch-Mallol, J.L. Isolation and characterization of endophytes from nodules of Mimosa pudica with biotechnological potential. Microbiol. Res. 2019, 218, 76–86. [Google Scholar] [CrossRef]
- Rosas-Galván, N.S.; Martínez-Morales, F.; Marquina-Bahena, S.; Tinoco-Valencia, R.; Serrano-Carreón, L.; Bertrand, B.; León-Rodríguez, R.; Guzmán-Aparicio, J.; Alvaréz-Berber, L.; Trejo-Hernández, M.d.R. Improved production, purification, and characterization of biosurfactants produced by Serratia marcescens SM3 and its isogenic SMRG-5 strain. Biotechnol. Appl. Biochem. 2018, 65, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Das, P.; Sen, R. Towards commercial production of microbial surfactants. Trends Biotechnol. 2006, 24, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Phetrong, K.; H-Kittikun, A.; Maneerat, S. Production and characterization of bioemulsifier from a marine bacterium, Acinetobacter calcoaceticus subsp. anitratus SM7. Songklanakarin J. Sci. Technol. 2008, 30, 297–305. [Google Scholar]
- García-Reyes, S.; Yáñez-Ocampo, G.; Wong-Villarreal, A.; Rajaretinam, R.K.; Thavasimuthu, C.; Patiño, R.; Ortiz-Hernández, M.L. Partial characterization of a biosurfactant extracted from Pseudomonas sp. B0406 that enhances the solubility of pesticides. Environ. Technol. 2018, 39, 2622–2631. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Soni, J.; Kaur, G.; Kaur, J. A study on biosurfactant production in Lactobacillus and Bacillus sp. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 723–733. [Google Scholar]
- García-Reyes, S.; Yañez-Ocampo, G. Microbial Biosurfactants: Methods for Their Isolation and Characterization. J. Microbiol. Biotechnol. Food Sci. 2016, 6, 641–648. [Google Scholar] [CrossRef]
- Dubey, K.V.; Charde, P.N.; Meshram, S.U.; Shendre, L.P.; Dubey, V.S.; Juwarkar, A.A. Surface-active potential of biosurfactants produced in curd whey by Pseudomonas aeruginosa strain-PP2 and Kocuria turfanesis strain-J at extreme environmental conditions. Bioresour. Technol. 2012, 126, 368–374. [Google Scholar] [CrossRef]
- Ben Ayed, H.; Jemil, N.; Maalej, H.; Bayoudh, A.; Hmidet, N.; Nasri, M. Enhancement of solubilization and biodegradation of diesel oil by biosurfactant from Bacillus amyloliquefaciens An6. Int. Biodeterior. Biodegrad. 2015, 99, 8–14. [Google Scholar] [CrossRef]
- Cua-Basulto, M.E.; Ruiz-Sánchez, E.; Chan-Cupul, W.; Reyes-Ramírez, A.; Ballina-Gómez, H.; Núñez, E.H.; Martin-Mex, R.; Gorocica, Á.M.H.; Ruiz-Jiménez, A.L. Effects of botanical acaricides on Tetranychus urticae and compatibility with the predatory mite Amblyseius swirskii. Arch. Phytopathol. Plant Prot. 2023, 57, 1359–1371. [Google Scholar] [CrossRef]
- Nexticapan-Garcéz, Á.; Cua-Basulto, M.; Martín-Mex, R.; Pérez-Brito, D.; Larqué-Saavedra, A.; Villanueva-Couoh, E.; Pérez-Gutiérrez, A.; Ruiz-Sánchez, E. Effects of botanical acaricides on Raoiella indica and Oligonychus sp. and their toxicity on two species of phytoseiid predatory mites. Arch. Phytopathol. Plant Prot. 2021, 54, 2221–2232. [Google Scholar] [CrossRef]
- Cua-Basulto, M.; Ruiz-Sánchez, E.; Chan-Cupul, W.; Reyes-Ramírez, A.; Ballina-Gómez, H.; Hernández-Nuñez, E. Effects of chemical acaricides on the mortality of the two spotted spider mite Tetranychus urticae KOCH (Acari: Tetranychidae). Trop. Subtrop. Agroecosyst. 2022, 25, 1–9. [Google Scholar] [CrossRef]
- Buchoux, S.; Lai-Kee-Him, J.; Garnier, M.; Tsan, P.; Besson, F.; Brisson, A.; Dufourc, E.J. Surfactin-Triggered Small Vesicle Formation of Negatively Charged Membranes: A Novel Membrane-Lysis Mechanism. Biophys. J. 2008, 95, 3840–3849. [Google Scholar] [CrossRef] [PubMed]
- Yun, D.C.; Yang, S.Y.; Kim, Y.C.; Kim, I.S.; Kim, Y.H. Identification of surfactin as an aphicidal metabolite produced by Bacillus amyloliquefaciens G1. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 751–753. [Google Scholar] [CrossRef]
- Mnif, I.; Elleuch, M.; Chaabouni, S.E.; Ghribi, D. Bacillus subtilis SPB1 biosurfactant: Production optimization and insecticidal activity against the carob moth Ectomyelois ceratoniae. Crop Prot. 2013, 50, 66–72. [Google Scholar] [CrossRef]
- Ghribi, D.; Abdelkefi-Mesrati, L.; Boukedi, H.; Elleuch, M.; Ellouze-Chaabouni, S.; Tounsi, S. The impact of the Bacillus subtilis SPB1 biosurfactant on the midgut histology of Spodoptera littoralis (Lepidoptera: Noctuidae) and determination of its putative receptor. J. Invertebr. Pathol. 2012, 109, 183–186. [Google Scholar] [CrossRef]
- Ben Khedher, S.; Boukedi, H.; Laarif, A.; Tounsi, S. Biosurfactant produced by Bacillus subtilis V26: A potential biological control approach for sustainable agriculture development. Org. Agric. 2020, 10, 117–124. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Cuatlayotl-Cottier, R.; la Peña, A.H.-D.; Peña-Chora, G.; Salazar-Magallón, J.A. Insecticidal activity of industrial by-products fermented by Bacillus thuringiensis strain GP139 against Mites (Prostigmata: Tetranychidae) and Aphids (Hemiptera: Aphidoidea). Biocontrol Sci. Technol. 2020, 32, 103–109. [Google Scholar] [CrossRef]
- Gómez-Gutiérrez, J.A.; Wong-Villarreal, A.; Aguilar-Marcelino, L.; Yañez-Ocampo, G.; Hernández-Nuñéz, E.; Caspeta-Mandujano, J.M.; García-Flores, A.; Cruz-Arévalo, J.; Vargas-Uriostegui, P.; Gomez-Rodríguez, O. In vitro nematicidal and acaricidal effect of biosurfactants produced by Bacillus against the root-knot nematode Nacobbus aberrans and the dust mite Tyrophagus putrescentiae. Braz. J. Microbiol. 2023, 54, 1127–1136. [Google Scholar] [CrossRef]
- Li, X.-Y.; Wang, Y.-H.; Yang, J.; Cui, W.-Y.; Munir, S.; He, P.-F.; Wu, Y.-X.; He, Y.-Q. Acaricidal Activity of Cyclodipeptides from Bacillus amyloliquefaciens W1 against Tetranychus urticae. J. Agric. Food Chem. 2018, 66, 10163–10168. [Google Scholar] [CrossRef]
- Li, X.; Munir, S.; Xu, Y.; Wang, Y.; He, Y. Combined mass spectrometry-guided genome mining and virtual screening for acaricidal activity in secondary metabolites of Bacillus velezensis W1. RSC Adv. 2021, 11, 25441–25449. [Google Scholar] [CrossRef] [PubMed]
- Fazaeli, N.; Bahador, N.; Hesami, S. A Study on Larvicidal Activity and Phylogenetic Analysis of Staphylococcus epidermidis as a Biosurfactant-Producing Bacterium. Pol. J. Environ. Stud. 2021, 30, 4511–4519. [Google Scholar] [CrossRef]
- Penha, R.O.; Vandenberghe, L.P.S.; Faulds, C.; Soccol, V.T.; Soccol, C.R. Bacillus lipopeptides as powerful pest control agents for a more sustainable and healthy agriculture: Recent studies and innovations. Planta 2020, 251, 70. [Google Scholar] [CrossRef] [PubMed]
- Long, W.; Zhao, W.; He, L.; Khan, T.A.; Lai, X.; Sun, Y.; Huang, W.; Yi, G.; Xia, L. Streptomyces enissocaesilis L-82 has broad-spectrum antibacterial activity and promotes growth for Carassius auratus. Appl. Microbiol. Biotechnol. 2024, 108, 220. [Google Scholar] [CrossRef]
- Mahar, A.N.; Munir, M.; Elawad, S.; Gowen, S.R.; Hague, N.G.M. Pathogenicity of bacterium, Xenorhabdus nematophila isolated from entomopathogenic nematode (Steinernema carpocapsae) and its secretion against Galleria mellonella larvae. J. Zhejiang Univ. B 2005, 6, 457–463. [Google Scholar] [CrossRef]
- Kegler, C.; Nollmann, F.I.; Ahrendt, T.; Fleischhacker, F.; Bode, E.; Bode, H.B. Rapid Determination of the Amino Acid Configuration of Xenotetrapeptide. ChemBioChem 2014, 15, 826–828. [Google Scholar] [CrossRef]
- Bode, H.B.; Brachmann, A.O.; Jadhav, K.B.; Seyfarth, L.; Dauth, C.; Fuchs, S.W.; Kaiser, M.; Waterfield, N.R.; Sack, H.; Heinemann, S.H.; et al. Structure Elucidation and Activity of Kolossin A, the D-/L-Pentadecapeptide Product of a Giant Nonribosomal Peptide Synthetase. Angew. Chem. Int. Ed. Engl. 2015, 54, 10352–10355. [Google Scholar] [CrossRef]
- Tobias, N.J.; Linck, A.; Bode, H.B. Natural Product Diversification Mediated by Alternative Transcriptional Starting. Angew. Chem. Int. Ed. Engl. 2018, 57, 5699–5702. [Google Scholar] [CrossRef]
- Clements, T.; Ndlovu, T.; Khan, S.; Khan, W. Biosurfactants produced by Serratia species: Classification, biosynthesis, production and application. Appl. Microbiol. Biotechnol. 2019, 103, 589–602. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Name | RT (min) | MW | Strains | ||
|---|---|---|---|---|---|
| S. ureilytica UTS | S. ureilytica NOD-3 | S. ureilytica NOD-7 | |||
| L-Proline | 7.66 | 115.13 | + | + | + |
| Hexadecanoic acid | 9.78 | 256.42 | + | + | + |
| Pentanoic acid | 10.21 | 102.13 | - | + | - |
| Octadecanoic acid | 17.42 | 284.50 | + | + | + |
| Glutaramic acid | 19.77 | 131.13 | - | + | - |
| L-Lysine | 22.64 | 182.65 | - | - | + |
| L-Valine | 22.74 | 117,151 | + | - | + |
| ç-Guanidinobutyric acid | 12.23 | 145.16 | + | - | - |
| Decanoic acid | 17.17 | 172.26 | + | - | - |
| Hexadecanamide | 19.19 | 255.44 | + | - | - |
| Octadecanamide | 20.91 | 283.50 | + | - | - |
| Tetradecanoic acid | 21.99 | 228.37 | + | - | - |
| Most Similar Known Gene Cluster | S. ureilytica UTS | S. ureilytica NOD-3 | S. ureilytica NOD-7 | |
|---|---|---|---|---|
| Product Type | Most Similar Product | |||
| NRPS | Prodigiosin | + | + | + |
| RRE-containing | Lankacidin C | + | + | + |
| T1PKS, NRPS | Viobactin | + | + | + |
| Thiopeptide | O-Antigen | + | + | + |
| Opine-like metallophore | Yersinopine | + | + | + |
| Betalactone | - | - | - | - |
| NRPS | 5-Dimethylallylindole-3-acetonitrile | - | + | - |
| Hserlactone | - | - | - | - |
| NRPS | Xenotetrapeptide | + | + | + |
| NRPS | RhizomideA/rhizomideB/rhizomideC | + | + | + |
| NRP-metallophore, NRPS | Trichrysobactin/cyclic trichrysobactin/chrysobactin/dichrysobactina | + | + | + |
| NRPS, thiopeptide, LAP | Microcin E492 | + | + | - |
| NRPS | - | - | - | - |
| NRPS | Kolossin | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong-Villareal, A.; Ruiz-Sánchez, E.; Cua-Basulto, M.; Espinosa-Zaragoza, S.; González-Sánchez, A.A.; Ramos-Carbajal, E.; Góngora-Gamboa, C.; Garruña-Hernández, R.; Romero-Tirado, R.; Moreno-Basurto, G.; et al. Acaricidal Activity of Biosurfactants Produced by Serratia ureilytica on Tetranychus urticae and Their Compatibility with the Predatory Mite Amblyseius swirskii. Microbiol. Res. 2025, 16, 150. https://doi.org/10.3390/microbiolres16070150
Wong-Villareal A, Ruiz-Sánchez E, Cua-Basulto M, Espinosa-Zaragoza S, González-Sánchez AA, Ramos-Carbajal E, Góngora-Gamboa C, Garruña-Hernández R, Romero-Tirado R, Moreno-Basurto G, et al. Acaricidal Activity of Biosurfactants Produced by Serratia ureilytica on Tetranychus urticae and Their Compatibility with the Predatory Mite Amblyseius swirskii. Microbiology Research. 2025; 16(7):150. https://doi.org/10.3390/microbiolres16070150
Chicago/Turabian StyleWong-Villareal, Arnoldo, Esaú Ruiz-Sánchez, Marcos Cua-Basulto, Saúl Espinosa-Zaragoza, Avel A. González-Sánchez, Ernesto Ramos-Carbajal, Cristian Góngora-Gamboa, René Garruña-Hernández, Rodrigo Romero-Tirado, Guillermo Moreno-Basurto, and et al. 2025. "Acaricidal Activity of Biosurfactants Produced by Serratia ureilytica on Tetranychus urticae and Their Compatibility with the Predatory Mite Amblyseius swirskii" Microbiology Research 16, no. 7: 150. https://doi.org/10.3390/microbiolres16070150
APA StyleWong-Villareal, A., Ruiz-Sánchez, E., Cua-Basulto, M., Espinosa-Zaragoza, S., González-Sánchez, A. A., Ramos-Carbajal, E., Góngora-Gamboa, C., Garruña-Hernández, R., Romero-Tirado, R., Moreno-Basurto, G., & Pinson-Rincón, E. P. (2025). Acaricidal Activity of Biosurfactants Produced by Serratia ureilytica on Tetranychus urticae and Their Compatibility with the Predatory Mite Amblyseius swirskii. Microbiology Research, 16(7), 150. https://doi.org/10.3390/microbiolres16070150