
Antisense Oligonucleotide-Capped Gold Nanoparticles as a Potential Strategy for Tackling Antimicrobial Resistance

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Bacterial Strain and Culture Conditions
2.3. Antisense Oligonucleotides
2.4. Transformation of E. coli DH5α into an Ampicillin-Resistant Strain
2.5. Plasmid DNA Extraction
2.6. Restriction Digestion and Gel Electrophoresis
2.7. Synthesis of AuNPs
2.8. ASO Thiol Activation and AuNP Conjugation
2.8.1. Thiol Activation of ASOs
2.8.2. Conjugation of ASOs to Gold Nanoparticles
2.9. TEM Characterization of AuNPs and ASO-AuNPs
2.10. Bacterial Susceptibility Testing
2.11. Statistical Analysis
3. Results and Discussion
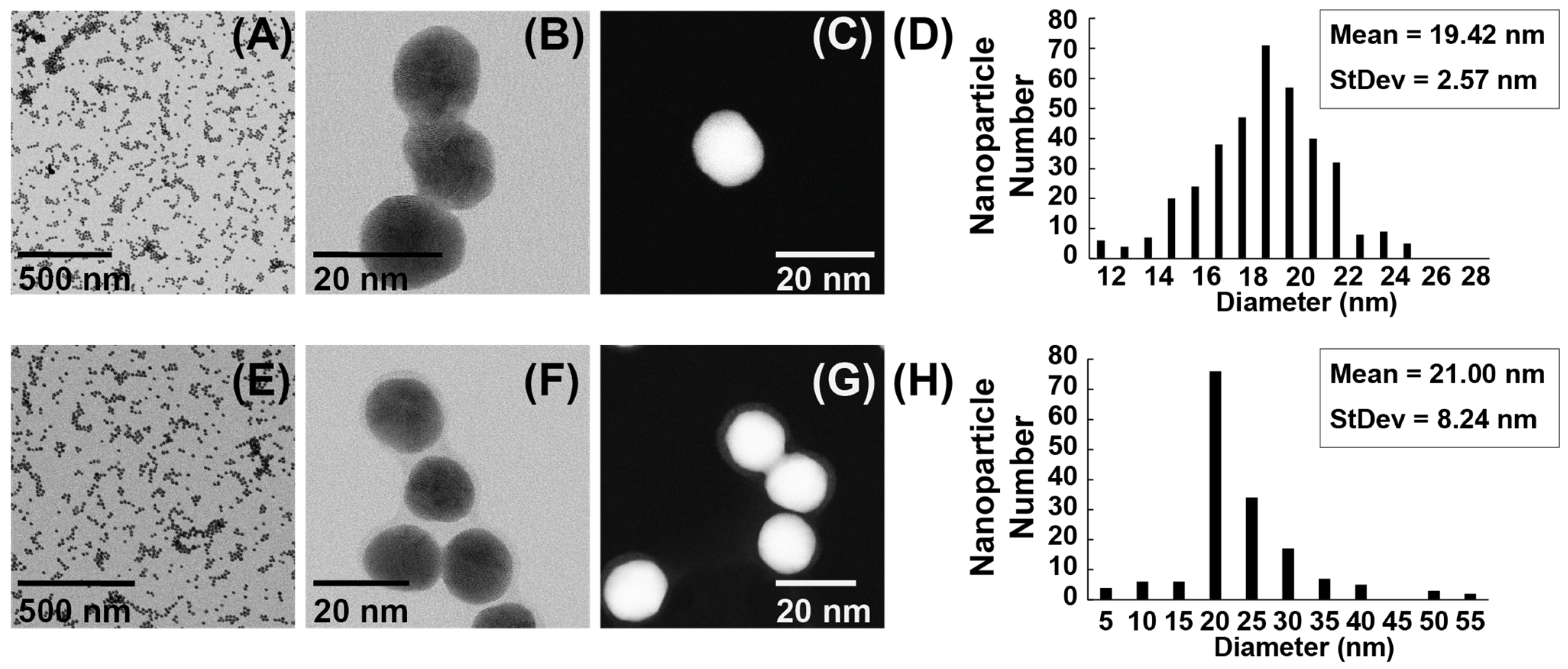
3.1. Characterization of AuNPs and ASO-AuNPs
3.2. Transmission Electron Microscopy and Size Distribution
3.3. Colloidal Stability of ASO-AuNPs
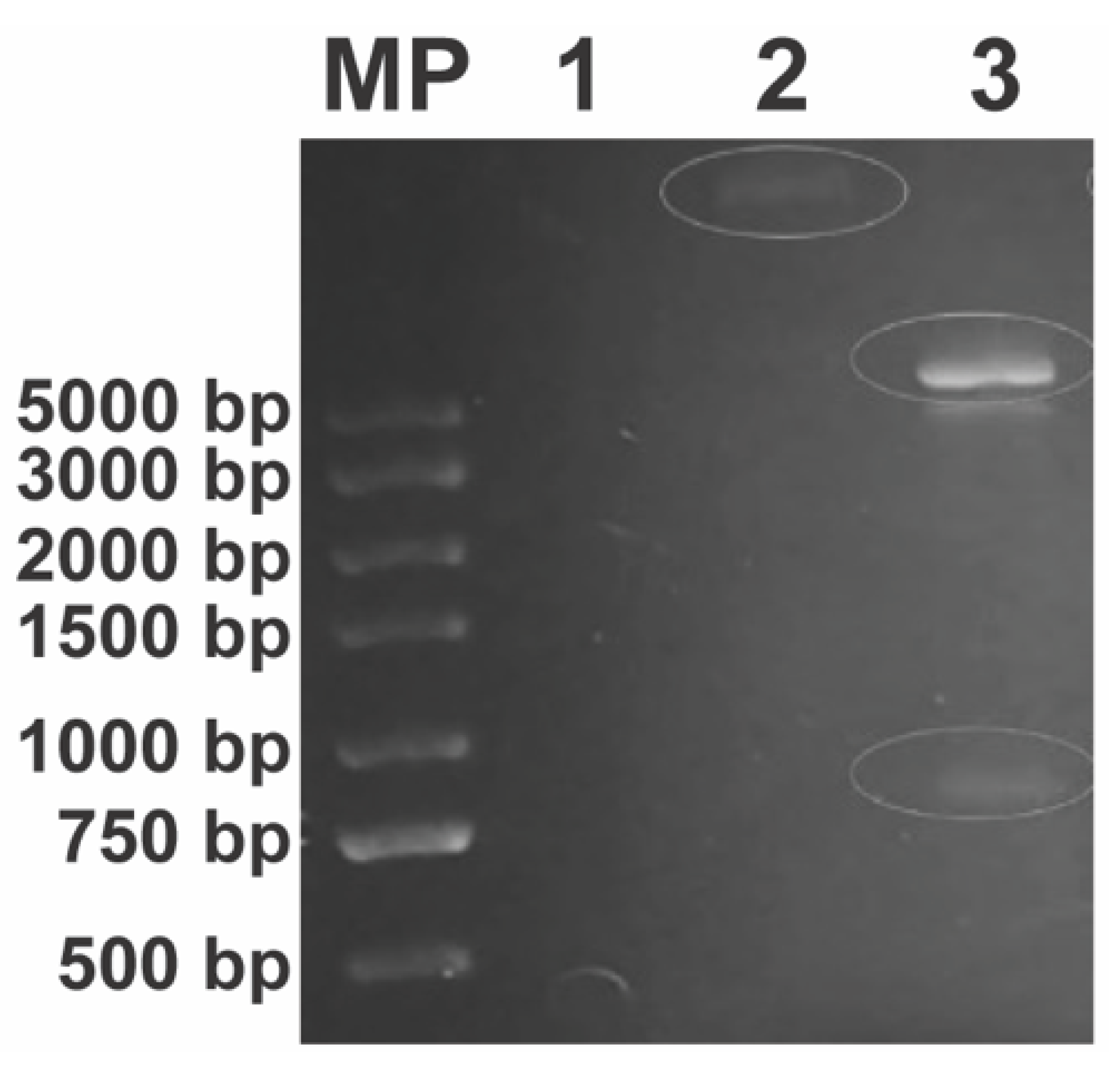
3.4. Generation and Confirmation of Ampicillin-Resistant E. coli
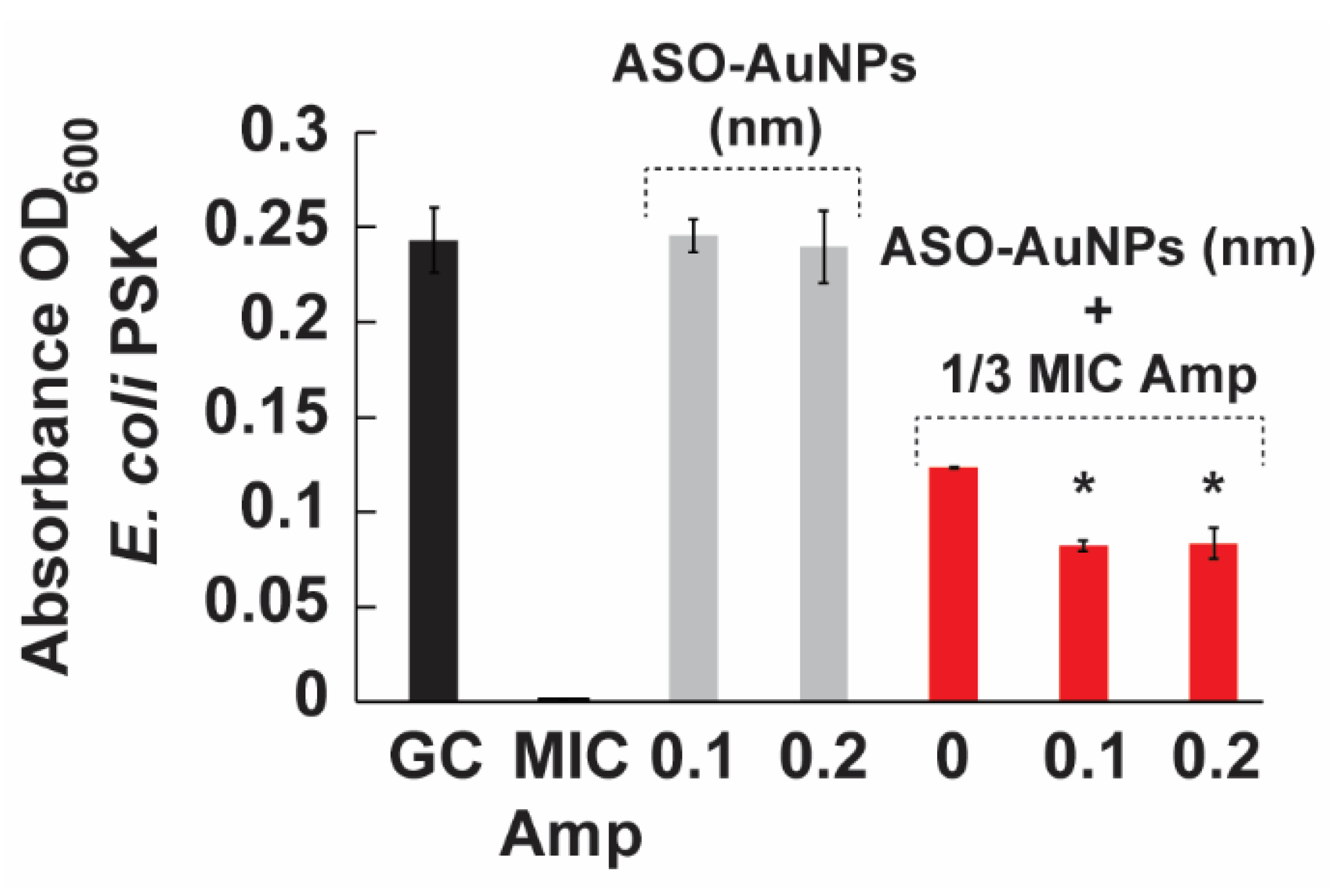
3.5. Antimicrobial Susceptibility Testing
3.5.1. Minimum Inhibitory Concentration
3.5.2. Combination Therapy for Ampicillin-Resistant Bacteria
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Medina, E.; Pieper, D.H. Tackling threats and future problems of multidrug-resistant bacteria. In How to Overcome the Antibiotic Crisis: Facts, Challenges, Technologies and Future Perspectives; Springer: Berlin/Heidelberg, Germany, 2016; pp. 3–33. [Google Scholar]
- Van Bambeke, F.; Balzi, E.; Tulkens, P.M. Antibiotic efflux pumps. Biochem. Pharmacol. 2000, 60, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Riu, F.; Ruda, A.; Ibba, R.; Sestito, S.; Lupinu, I.; Piras, S.; Widmalm, G.; Carta, A. Antibiotics and carbohydrate-containing drugs targeting bacterial cell envelopes: An overview. Pharmaceuticals 2022, 15, 942. [Google Scholar] [CrossRef]
- Tabatabaeifar, F.; Isaei, E.; Kalantar-Neyestanaki, D.; Morones-Ramírez, J.R. Antimicrobial and Antibiofilm Effects of Combinatorial Treatment Formulations of Anti-Inflammatory Drugs—Common Antibiotics against Pathogenic Bacteria. Pharmaceutics 2022, 15, 4. [Google Scholar] [CrossRef]
- Hegarty, J.P.; Stewart, D.B. Advances in therapeutic bacterial antisense biotechnology. Appl. Microbiol. Biotechnol. 2018, 102, 1055–1065. [Google Scholar] [CrossRef]
- Cheng, Y.-S.; Williamson, P.R.; Zheng, W. Improving therapy of severe infections through drug repurposing of synergistic combinations. Curr. Opin. Pharmacol. 2019, 48, 92–98. [Google Scholar] [CrossRef]
- Meng, J.; He, G.; Wang, H.; Jia, M.; Ma, X.; Da, F.; Wang, N.; Hou, Z.; Xue, X.; Li, M.; et al. Reversion of antibiotic resistance by inhibiting mecA in clinical methicillin-resistant Staphylococci by antisense phosphorothioate oligonucleotide. J. Antibiot. 2015, 68, 158–164. [Google Scholar] [CrossRef]
- Xue, X.Y.; Mao, X.G.; Zhou, Y.; Chen, Z.; Hu, Y.; Hou, Z.; Li, M.K.; Meng, J.R.; Luo, X.X. Advances in the delivery of antisense oligonucleotides for combating bacterial infectious diseases. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 745–758. [Google Scholar] [CrossRef]
- Angrish, N.; Khare, G. Antisense oligonucleotide based therapeutics and its applications against bacterial infections. Med. Drug Discov. 2023, 20, 100166. [Google Scholar] [CrossRef]
- Good, L.; Stach, J.E. Synthetic RNA silencing in bacteria–antimicrobial discovery and resistance breaking. Front. Microbiol. 2011, 2, 185. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Meng, J.; Jia, M.; Ma, X.; He, G.; Yu, J.; Wang, R.; Bai, H.; Hou, Z.; Luo, X. oprM as a new target for reversion of multidrug resistance in Pseudomonas aeruginosa by antisense phosphorothioate oligodeoxynucleotides. FEMS Immunol. Med. Microbiol. 2010, 60, 275–282. [Google Scholar] [CrossRef] [PubMed]
- DeLong, R.K.; Reynolds, C.M.; Malcolm, Y.; Schaeffer, A.; Severs, T.; Wanekaya, A. Functionalized gold nanoparticles for the binding, stabilization, and delivery of therapeutic DNA, RNA, and other biological macromolecules. Nanotechnol. Sci. Appl. 2010, 3, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.K.; Vermerris, W. Recent advances in nanomaterials for gene delivery—A review. Nanomaterials 2017, 7, 94. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Morones, J.R.; Frey, W. Room temperature synthesis of an optically and thermally responsive hybrid PNIPAM–gold nanoparticle. J. Nanoparticle Res. 2010, 12, 1401–1414. [Google Scholar] [CrossRef]
- Morones-Ramírez, J.R. Bioinspired synthesis of optically and thermally responsive nanoporous membranes. NPG Asia Mater. 2013, 5, e52. [Google Scholar] [CrossRef]
- Rafiei, N.; Alishah Aratboni, H.; Alemzadeh, A.; Saavedra-Alonso, S.; Razi, H.; Morones-Ramírez, J.R. Nano-Regulation of Gene Expression in Chlamydomonas reinhardtii: Harnessing AuNPs for Remotely Switchable Lipid Biosynthesis via Antisense Oligonucleotides. ACS Synth. Biol. 2024, 13, 1694–1704. [Google Scholar] [CrossRef]
- Grönbeck, H.; Curioni, A.; Andreoni, W. Thiols and disulfides on the Au (111) surface: The headgroup—Gold interaction. J. Am. Chem. Soc. 2000, 122, 3839–3842. [Google Scholar] [CrossRef]
- Ghosh, S.; Su, Y.H.; Yang, C.J.; Lai, J.Y. Design of Highly Adhesive Urchin-Like Gold Nanostructures for Effective Topical Drug Administration and Symptomatic Relief of Corneal Dryness. Small Struct. 2025, 6, 2400484. [Google Scholar] [CrossRef]
- Han, G.; Ghosh, P.; Rotello, V.M. Functionalized gold nanoparticles for drug delivery. Nanomedicine 2007, 2, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Okkeh, M.; Bloise, N.; Restivo, E.; De Vita, L.; Pallavicini, P.; Visai, L. Gold nanoparticles: Can they be the next magic bullet for multidrug-resistant bacteria? Nanomaterials 2021, 11, 312. [Google Scholar] [CrossRef] [PubMed]
- Hurst, S.J.; Lytton-Jean, A.K.; Mirkin, C.A. Maximizing DNA loading on a range of gold nanoparticle sizes. Anal. Chem. 2006, 78, 8313–8318. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Li, X.; Li, H.; Zhang, W. Quantifying thiol–gold interactions towards the efficient strength control. Nat. Commun. 2014, 5, 4348. [Google Scholar] [CrossRef]
- Hammel, M.; Fan, Y.; Sarode, A.; Byrnes, A.E.; Zang, N.; Kou, P.; Nagapudi, K.; Leung, D.; Hoogenraad, C.C.; Chen, T.; et al. Correlating the structure and gene silencing activity of oligonucleotide-loaded lipid nanoparticles using small-angle X-ray scattering. ACS Nano 2023, 17, 11454–11465. [Google Scholar] [CrossRef]
- Sarode, A.; Fan, Y.; Byrnes, A.E.; Hammel, M.; Hura, G.L.; Fu, Y.; Kou, P.; Hu, C.; Hinz, F.I.; Roberts, J.; et al. Predictive high-throughput screening of PEGylated lipids in oligonucleotide-loaded lipid nanoparticles for neuronal gene silencing. Nanoscale Adv. 2022, 4, 2107–2123. [Google Scholar] [CrossRef]
- Ferreira, D.; Fernandes, A.R.; Baptista, P.V. Mild hyperthermia via gold nanoparticles and visible light irradiation for enhanced siRNA and ASO delivery in 2D and 3D tumour spheroids. Cancer Nanotechnol. 2024, 15, 19. [Google Scholar] [CrossRef]
- García-García, P.; Ruiz, M.; Reyes, R.; Delgado, A.; Évora, C.; Riancho, J.A.; Rodríguez-Rey, J.C.; Pérez-Campo, F.M. Smurf1 silencing using a LNA-ASOs/lipid nanoparticle system to promote bone regeneration. Stem Cells Transl. Med. 2019, 8, 1306–1317. [Google Scholar] [CrossRef]
- Kime, L.; Randall, C.P.; Banda, F.I.; Coll, F.; Wright, J.; Richardson, J.; Empel, J.; Parkhill, J.; O’Neill, A.J. Transient silencing of antibiotic resistance by mutation represents a significant potential source of unanticipated therapeutic failure. MBio 2019, 10, 10–1128. [Google Scholar] [CrossRef]
- Hoynes-O’Connor, A.; Moon, T.S. Development of design rules for reliable antisense RNA behavior in E. coli. ACS Synth. Biol. 2016, 5, 1441–1454. [Google Scholar] [CrossRef]
- Michel, S.; Schirduan, K.; Shen, Y.; Klar, R.; Tost, J.; Jaschinski, F. Using RNA-seq to assess off-target effects of antisense oligonucleotides in human cell lines. Mol. Diagn. Ther. 2021, 25, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, H.; Yoshida, T.; Sasaki, K.; Obika, S.; Inoue, T. Reduction of off-target effects of gapmer antisense oligonucleotides by oligonucleotide extension. Mol. Diagn. Ther. 2022, 26, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Terada, C.; Oh, K.; Tsubaki, R.; Chan, B.; Aibara, N.; Ohyama, K.; Shibata, M.A.; Wada, T.; Harada-Shiba, M.; Yamayoshi, A.; et al. Dynamic and static control of the off-target interactions of antisense oligonucleotides using toehold chemistry. Nat. Commun. 2023, 14, 7972. [Google Scholar] [CrossRef]
- Tu, Q.; Yin, J.; Fu, J.; Herrmann, J.; Li, Y.; Yin, Y.; Stewart, A.F.; Müller, R.; Zhang, Y. Room temperature electrocompetent bacterial cells improve DNA transformation and recombineering efficiency. Sci. Rep. 2016, 6, 24648. [Google Scholar] [CrossRef]
- Ghaffari, E.; Rezatofighi, S.E.; Ardakani, M.R.; Rastegarzadeh, S. Delivery of antisense peptide nucleic acid by gold nanoparticles for the inhibition of virus replication. Nanomedicine 2019, 14, 1827–1840. [Google Scholar] [CrossRef]
- Haiss, W.; Thanh, N.T.; Aveyard, J.; Fernig, D.G. Determination of size and concentration of gold nanoparticles from UV− Vis spectra. Anal. Chem. 2007, 79, 4215–4221. [Google Scholar] [CrossRef]
- Garza-Cervantes, J.A.; Chávez-Reyes, A.; Castillo, E.C.; García-Rivas, G.; Antonio Ortega-Rivera, O.; Salinas, E.; Ortiz-Martínez, M.; Gómez-Flores, S.L.; Peña-Martínez, J.A.; Pepi-Molina, A.; et al. Synergistic antimicrobial effects of silver/transition-metal combinatorial treatments. Sci. Rep. 2017, 7, 903. [Google Scholar] [CrossRef]
- Tyagi, H.; Kushwaha, A.; Kumar, A.; Aslam, M. A facile pH controlled citrate-based reduction method for gold nanoparticle synthesis at room temperature. Nanoscale Res. Lett. 2016, 11, 362. [Google Scholar] [CrossRef]
- Amendola, V.; Pilot, R.; Frasconi, M.; Maragò, O.M.; Iatì, M.A. Surface plasmon resonance in gold nanoparticles: A review. J. Phys. Condens. Matter 2017, 29, 203002. [Google Scholar] [CrossRef]
- Storhoff, J.J.; Elghanian, R.; Mucic, R.C.; Mirkin, C.A.; Letsinger, R.L. One-pot colorimetric differentiation of polynucleotides with single base imperfections using gold nanoparticle probes. J. Am. Chem. Soc. 1998, 120, 1959–1964. [Google Scholar] [CrossRef]
- Alishah Aratboni, H.; Rafiei, N.; Uscanga-Palomeque, A.C.; Luna Cruz, I.E.; Parra-Saldivar, R.; Morones-Ramirez, J.R. Design of a nanobiosystem with remote photothermal gene silencing in Chlamydomonas reinhardtii to increase lipid accumulation and production. Microb. Cell Factories 2023, 22, 61. [Google Scholar] [CrossRef]
- Allen, N.C.; Chauhan, R.; Bates, P.J.; O’Toole, M.G. Optimization of tumor targeting gold nanoparticles for glioblastoma applications. Nanomaterials 2022, 12, 3869. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.; Ghalami, M.; Ghaemi, A.; Mosavari, N.; Abdul-Tehrani, H.; Sadeghizadeh, M. Nanodiagnostic method for colorimetric detection of Mycobacterium tuberculosis 16S rRNA. Nanobiotechnology 2008, 4, 28–35. [Google Scholar] [CrossRef]
- Giljohann, D.A.; Seferos, D.S.; Daniel, W.L.; Massich, M.D.; Patel, P.C.; Mirkin, C.A. Gold nanoparticles for biology and medicine. In Spherical Nucleic Acids; Jenny Stanford Publishing: Singapore, 2020; pp. 55–90. [Google Scholar]
- Giljohann, D.A.; Seferos, D.S.; Patel, P.C.; Millstone, J.E.; Rosi, N.L.; Mirkin, C.A. Oligonucleotide loading determines cellular uptake of DNA-modified gold nanoparticles. Nano Lett. 2007, 7, 3818–3821. [Google Scholar] [CrossRef] [PubMed]
- Javed, R.; Zia, M.; Naz, S.; Aisida, S.O.; Ain Nu Ao, Q. Role of capping agents in the application of nanoparticles in biomedicine and environmental remediation: Recent trends and future prospects. J. Nanobiotechnol. 2020, 18, 172. [Google Scholar] [CrossRef]
- Escárcega-González, C.E.; Garza-Cervantes, J.A.; Vázquez-Rodríguez, A.; Morones-Ramírez, J.R. Bacterial exopolysaccharides as reducing and/or stabilizing agents during synthesis of metal nanoparticles with biomedical applications. Int. J. Polym. Sci. 2018, 2018, 7045852. [Google Scholar] [CrossRef]
- Li, Z.; Jin, R.; Mirkin, C.A.; Letsinger, R.L. Multiple thiol-anchor capped DNA–gold nanoparticle conjugates. Nucleic Acids Res. 2002, 30, 1558–1562. [Google Scholar] [CrossRef]
- Narendrakumar, L.; Chakraborty, M.; Kumari, S.; Paul, D.; Das, B. β-Lactam potentiators to re-sensitize resistant pathogens: Discovery, development, clinical use and the way forward. Front. Microbiol. 2023, 13, 1092556. [Google Scholar] [CrossRef]
- Rojas, M.; Del Valle, D. Betalactamasas tipo AmpC: Generalidades y métodos para detección fenotípica. Rev. De La Soc. Venez. De Microbiol. 2009, 29, 78–83. [Google Scholar]
- Then, R. Beta-Lactamase. In xPharm: The Comprehensive Pharmacology Reference; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–9. [Google Scholar] [CrossRef]
- Brandt, C.; Braun, S.D.; Stein, C.; Slickers, P.; Ehricht, R.; Pletz, M.W.; Makarewicz, O. In silico serine β-lactamases analysis reveals a huge potential resistome in environmental and pathogenic species. Sci. Rep. 2017, 7, 43232. [Google Scholar] [CrossRef]
- Brinas, L.; Zarazaga, M.; Sáenz, Y.; Ruiz-Larrea, F.; Torres, C. β-Lactamases in ampicillin-resistant Escherichia coli isolates from foods, humans, and healthy animals. Antimicrob. Agents Chemother. 2002, 46, 3156–3163. [Google Scholar] [CrossRef] [PubMed]
- de Boer, M.; Heuer, C.; Hussein, H.; McDougall, S. Minimum inhibitory concentrations of selected antimicrobials against Escherichia coli and Trueperella pyogenes of bovine uterine origin. J. Dairy Sci. 2015, 98, 4427–4438. [Google Scholar] [CrossRef]
- Garza-Cervantes, J.A.; Meza-Bustillos, J.F.; Resendiz-Hernández, H.; Suárez-Cantú, I.A.; Ortega-Rivera, O.A.; Salinas, E.; Escárcega-González, C.E.; Morones-Ramírez, J.R. Re-sensitizing ampicillin and kanamycin-resistant E. coli and S. aureus using synergistic metal micronutrients-antibiotic combinations. Front. Bioeng. Biotechnol. 2020, 8, 612. [Google Scholar]
- Olesen, I.; Hasman, H.; Møller Aarestrup, F. Prevalence of β-lactamases among ampicillin-resistant Escherichia coli and Salmonella isolated from food animals in Denmark. Microb. Drug Resist. 2004, 10, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Jani, S.; Ramirez, M.S.; Tolmasky, M.E. Silencing antibiotic resistance with antisense oligonucleotides. Biomedicines 2021, 9, 416. [Google Scholar] [CrossRef]
- Binzel, D.W.; Li, X.; Burns, N.; Khan, E.; Lee, W.J.; Chen, L.C.; Ellipilli, S.; Miles, W.; Ho, Y.S.; Guo, P. Thermostability, tunability, and tenacity of RNA as rubbery anionic polymeric materials in nanotechnology and nanomedicine—Specific cancer targeting with undetectable toxicity. Chem. Rev. 2021, 121, 7398–7467. [Google Scholar] [CrossRef]
- Cela, E.M.; Urquiza, D.; Gómez, M.I.; Gonzalez, C.D. New Weapons to Fight against Staphylococcus aureus Skin Infections. Antibiotics 2023, 12, 1477. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Delivery Platform/Method | Target Gene/Organism | Key Outcome | Remarks | Study/Reference |
|---|---|---|---|---|
| Anionic liposome encapsulates anti-oprM phosphorothioate oligodeoxynucleotide (PS-ODN617) and PEI | oprM in MDR Pseudomonas aeruginosa | Significantly reduced oprM expression; lowered MICs for several commonly used antibiotics; improved susceptibility to piperacillin | Demonstrated that targeting oprM disrupts MexAB-OprM efflux pump function and resensitizes MDR-PA isolates to multiple antibiotics | [13] |
| Liposome-based antisense phosphorothioate oligonucleotide | mecA in MRSA/MRSE | Restored susceptibility to β-lactams in clinical Staphylococci; inhibited bacterial growth in vitro and in vivo | Identified a conserved target sequence in mecA (nt 854–871); anti-mecA PS-ODN04 dramatically reduced bacterial load and improved survival in mouse models | [9] |
| Lipid nanoparticles (LNPs) with varying PEG-lipids for antisense oligonucleotide (ASO) delivery | TMEM106b in mammalian cells (gene silencing model) | Core structural organization (ordered vs. disordered inverted hexagonal phases) correlated with knockdown efficacy | Integrated high-throughput screening, SAXS, and cryo-EM to link LNP structure with in vitro gene silencing; showed that lower ratio of disordered/ordered core phases enhances ASO knockdown | [26] |
| High-throughput screening of PEGylated LNP formulations for antisense oligonucleotide loading | Murine cortical neurons (gene silencing in vitro) | Up to 5-fold lower mRNA expression compared to naked ASO; PEG-lipid composition controls particle size and gene silencing performance | Automated workflow to optimize PEG-lipid parameters; scaled-up microfluidic formulation confirmed translatability of ASO-LNP properties and efficacy | [27] |
| Gold nanoparticles (AuNPs) + visible light irradiation to induce mild hyperthermia for ASO/siRNA delivery | c-MYC in HCT116, MCF-7 (2D), and 3D tumor spheroids | 25–30% decrease in c-MYC expression under mild photothermy; improved transfection and spatial–temporal control of gene modulation | Demonstrated the potential of AuNP-mediated mild hyperthermia to enhance cellular uptake and gene silencing in both 2D and 3D tumor models | [28] |
| LNA-ASOs in lipid nanoparticle system (GapmeRs) | Smurf1 in MSCs for bone regeneration | Primed MSCs for osteogenesis, enabling lower BMP-2 doses; promoted formation of mature bone matrix in vivo | Demonstrated synergy between Smurf1 silencing and reduced BMP-2 for enhanced bone repair in osteoporotic models; clinically safe approach with no viral vectors | [29] |
| Direct antisense-based or partial gene silencing approach (mutation-induced silencing of resistance genes) | Multiple antibiotic resistance genes in Staphylococcus aureus | Many strains harbored silenced resistance determinants that revert at high frequencies, potentially causing treatment failures | Highlighted the prevalence of “silent” resistance genes that can spontaneously revert; underscored risk of underestimating antibiotic resistance if clinical microbiology only checks phenotype | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garza-Cardenas, C.R.; Leon-Buitimea, A.; Siller-Ceniceros, A.A.; Morones-Ramirez, J.R. Antisense Oligonucleotide-Capped Gold Nanoparticles as a Potential Strategy for Tackling Antimicrobial Resistance. Microbiol. Res. 2025, 16, 70. https://doi.org/10.3390/microbiolres16030070
Garza-Cardenas CR, Leon-Buitimea A, Siller-Ceniceros AA, Morones-Ramirez JR. Antisense Oligonucleotide-Capped Gold Nanoparticles as a Potential Strategy for Tackling Antimicrobial Resistance. Microbiology Research. 2025; 16(3):70. https://doi.org/10.3390/microbiolres16030070
Chicago/Turabian StyleGarza-Cardenas, Cesar Rodolfo, Angel Leon-Buitimea, A. A. Siller-Ceniceros, and Jose Ruben Morones-Ramirez. 2025. "Antisense Oligonucleotide-Capped Gold Nanoparticles as a Potential Strategy for Tackling Antimicrobial Resistance" Microbiology Research 16, no. 3: 70. https://doi.org/10.3390/microbiolres16030070
APA StyleGarza-Cardenas, C. R., Leon-Buitimea, A., Siller-Ceniceros, A. A., & Morones-Ramirez, J. R. (2025). Antisense Oligonucleotide-Capped Gold Nanoparticles as a Potential Strategy for Tackling Antimicrobial Resistance. Microbiology Research, 16(3), 70. https://doi.org/10.3390/microbiolres16030070