
Probiotic Lactobacilli Ameliorate Antibiotic-Induced Cognitive and Behavioral Impairments in Mice

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Probiotics
2.3. Experimental Design
2.4. Fecal Microbiota Analysis
2.5. Behavioral Testing
2.5.1. Open Field Test (OFT) and Integral Anxiety Score (IAS)
2.5.2. T-Maze
2.5.3. Novel Object Recognition Test (ORT)
2.6. Study of Physiological Traits of Probiotics
2.6.1. Antibiotic Susceptibility Testing
2.6.2. Determination of H2O2 Production
2.6.3. Assessment of Antioxidant Activity
2.6.4. Measurement of Bacterial Neurotransmitters
2.7. Statistical Analysis
3. Results
3.1. Modulation of Gut Microbiota by Antibiotics and Lactobacilli
3.2. Probiotics Ameliorated Antibiotics’ Toxicity and Antibiotic-Induced Mortality
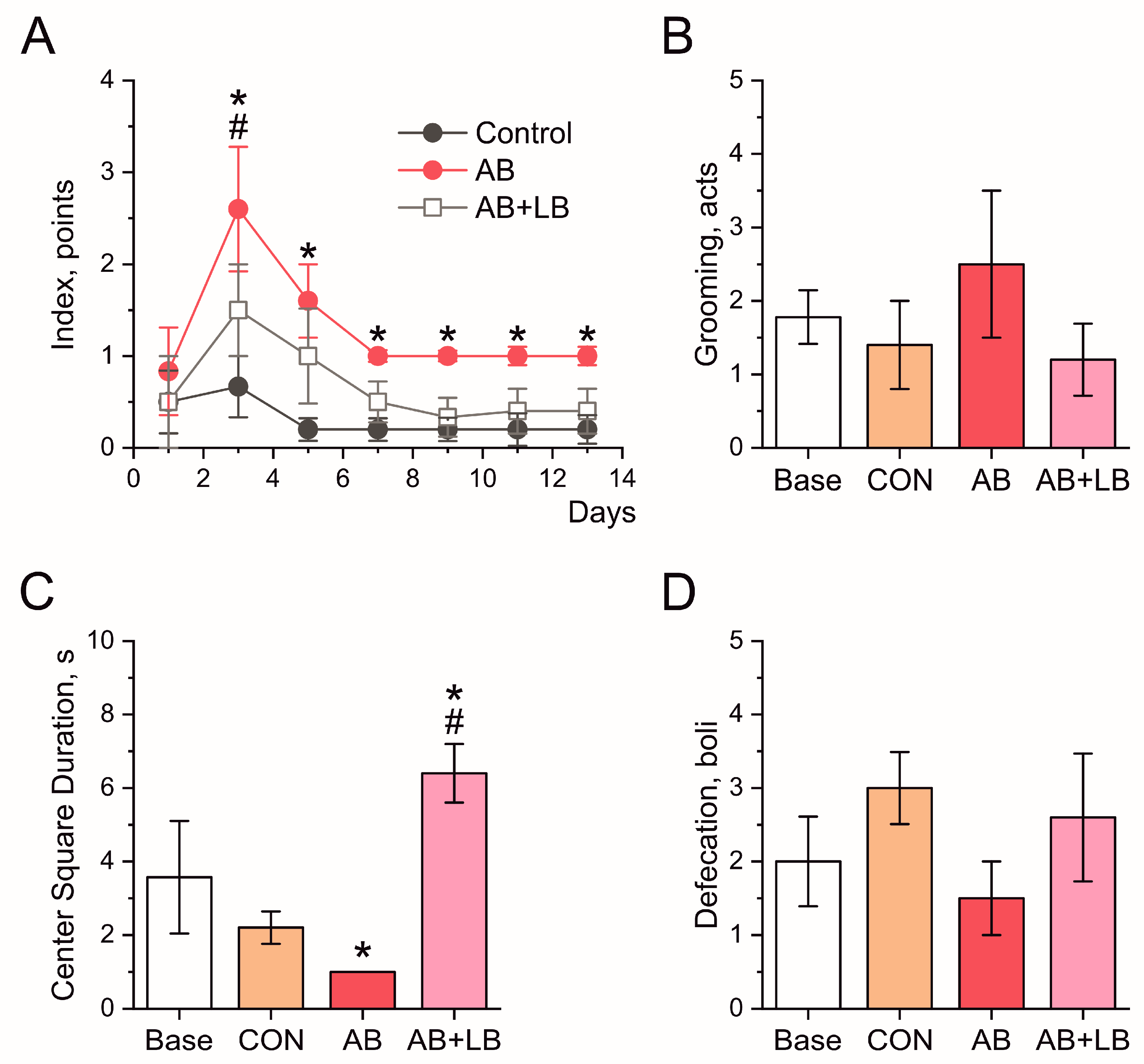
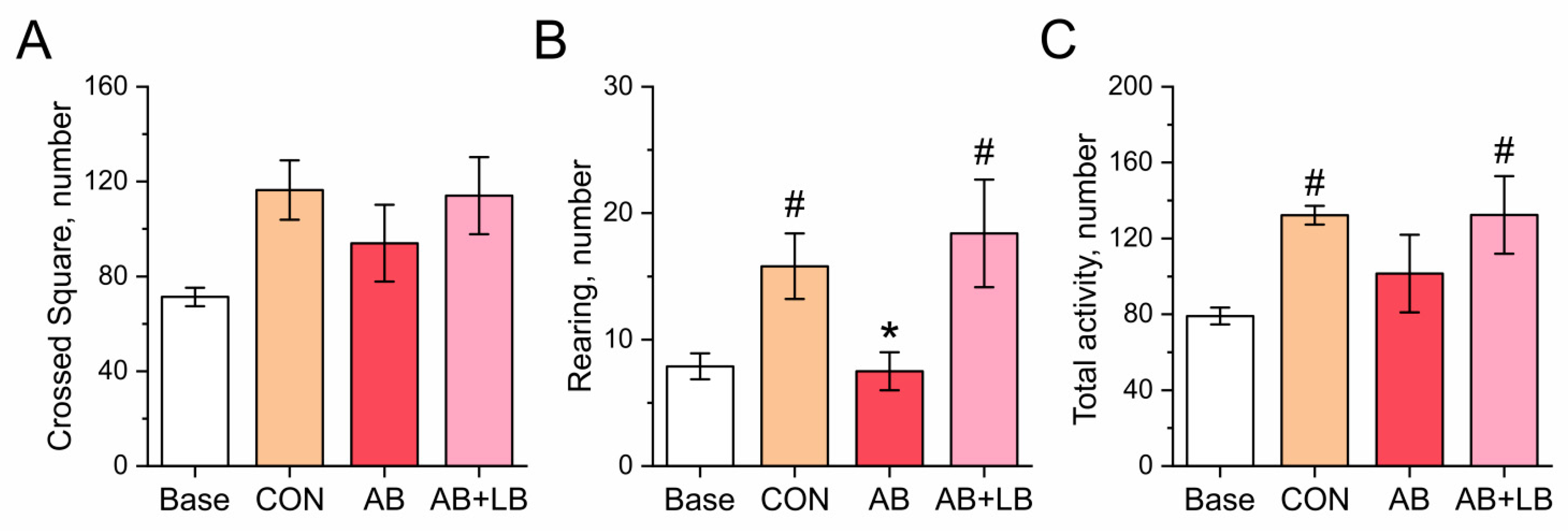
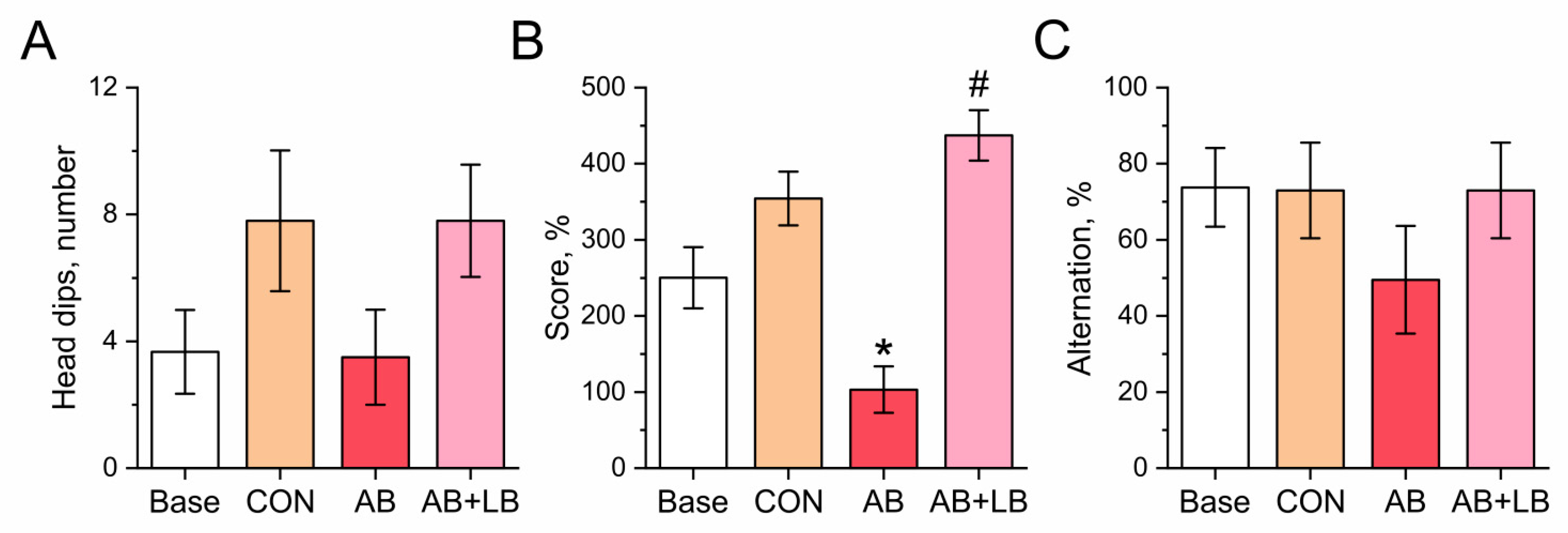
3.3. Probiotics Ameliorated Antibiotic-Induced Changes in Mouse Behavior
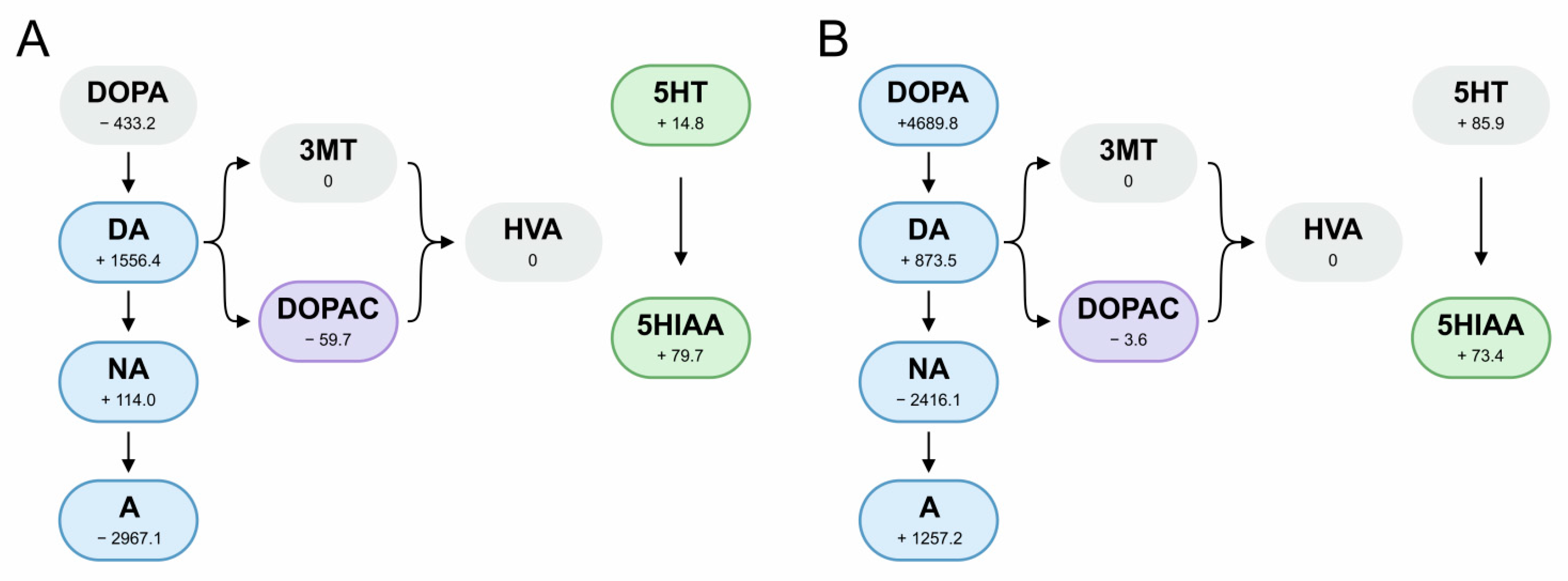
3.4. Lactobacilli’s Potential to Modulate Neurotransmitter Levels and Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Report of a Joint FAO/WHO Expert Consultation. Evaluation of Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. 2001. Available online: http://www.fao.org/tempref/docrep/fao/meeting/009/y6398e.pdf (accessed on 13 July 2024).
- Latif, A.; Shehzad, A.; Niazi, S.; Zahid, A.; Ashraf, W.; Iqbal, M.W.; Rehman, A.; Riaz, T.; Aadil, R.M.; Khan, I.M.; et al. Probiotics: Mechanism of action, health benefits and their application in food industries. Front. Microbiol. 2023, 14, 1216674. [Google Scholar] [CrossRef]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef]
- Yong, S.J.; Tong, T.; Chew, J.; Lim, W.L. Antidepressive mechanisms of probiotics and their therapeutic potential. Front. Neurosci. 2020, 13, 1361. [Google Scholar] [CrossRef]
- Ullah, H.; Arbab, S.; Tian, Y.; Liu, C.Q.; Chen, Y.; Qijie, L.; Khan, M.I.U.; Hassan, I.U.; Li, K. The gut microbiota-brain axis in neurological disorder. Front. Neurosci. 2023, 17, 1225875. [Google Scholar] [CrossRef]
- Sanguinetti, E.; Guzzardi, M.A.; Tripodi, M.; Panetta, D.; Selma-Royo, M.; Zega, A.; Telleschi, M.; Collado, M.C.; Iozzo, P. Microbiota signatures relating to reduced memory and exploratory behaviour in the offspring of overweight mothers in a murine model. Sci. Rep. 2019, 9, 12609. [Google Scholar] [CrossRef]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef]
- Messaoudi, M.; Lalonde, R.; Violle, N.; Javelot, H.; Desor, D.; Nejdi, A.; Bisson, J.F.; Rougeot, C.; Pichelin, M.; Cazaubiel, M.; et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 2011, 105, 755–764. [Google Scholar] [CrossRef]
- Ohland, C.L.; Kish, L.; Bell, H.; Thiesen, A.; Hotte, N.; Pankiv, E.; Madsen, K.L. Effects of Lactobacillus helveticus on murine behavior are dependent on diet and genotype and correlate with alterations in the gut microbiome. Psychoneuroendocrinology 2013, 38, 1738–1747. [Google Scholar] [CrossRef]
- Emge, J.R.; Huynh, K.; Miller, E.N.; Kaur, M.; Reardon, C.; Barrett, K.E.; Gareau, M.G. Modulation of the microbiota-gut-brain axis by probiotics in a murine model of inflammatory bowel disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G989–G998. [Google Scholar] [CrossRef]
- Arslanova, A.; Tarasova, A.; Alexandrova, A.; Novoselova, V.; Shaidullov, I.; Khusnutdinova, D.; Grigoryeva, T.; Yarullina, D.; Yakovleva, O.; Sitdikova, G. Protective effects of probiotics on cognitive and motor functions, anxiety level, visceral sensitivity, oxidative stress and microbiota in mice with antibiotic-induced dysbiosis. Life 2021, 11, 764. [Google Scholar] [CrossRef]
- Akkasheh, G.; Kashani-Poor, Z.; Tajabadi-Ebrahimi, M.; Jafari, P.; Akbari, H.; Taghizadeh, M.; Memarzadeh, M.R.; Asemi, Z.; Esmaillzadeh, A. Clinical and metabolic response to probiotic administration in patients with major depressive disorder: A randomized, double-blind, placebo-controlled trial. Nutrition 2016, 32, 315–320. [Google Scholar] [CrossRef]
- Feng, P.; Zhao, S.; Zhang, Y.; Li, E. A review of probiotics in the treatment of autism spectrum disorders: Perspectives from the gut-brain axis. Front. Microbiol. 2023, 14, 1123462. [Google Scholar] [CrossRef]
- Akbari, E.; Asemi, Z.; Daneshvar Kakhaki, R.; Bahmani, F.; Kouchaki, E.; Tamtaji, O.R.; Hamidi, G.A.; Salami, M. Effect of probiotic supplementation on cognitive function and metabolic status in Alzheimer’s disease: A randomized, double-blind and controlled trial. Front. Aging Neurosci. 2016, 8, 256. [Google Scholar] [CrossRef]
- Kouchaki, E.; Tamtaji, O.R.; Salami, M.; Bahmani, F.; Daneshvar Kakhaki, R.; Akbari, E.; Tajabadi-Ebrahimi, M.; Jafari, P.; Asemi, Z. Clinical and metabolic response to probiotic supplementation in patients with multiple sclerosis: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2017, 36, 1245–1249. [Google Scholar] [CrossRef]
- Oleskin, A.V.; El’-Registan, G.I.; Shenderov, B.A. Role of neuromediators in the functioning of the human microbiota: “Business talks” among microorganisms and the microbiota-host dialogue. Mikrobiologiia 2016, 85, 1–22. [Google Scholar] [CrossRef]
- Shandilya, S.; Kumar, S.; Kumar Jha, N.; Kumar Kesari, K.; Ruokolainen, J. Interplay of gut microbiota and oxidative stress: Perspective on neurodegeneration and neuroprotection. J. Adv. Res. 2021, 38, 223–244. [Google Scholar] [CrossRef]
- Tsapieva, A.; Duplik, N.; Suvorov, A. Structure of plantaricin locus of Lactobacillus plantarum 8P-A3. Benef. Microbes 2011, 2, 255–261. [Google Scholar] [CrossRef]
- De Man, J.C.; Rogosa, M.; Sharpe, M.T. A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Desbonnet, L.; Clarke, G.; Traplin, A.; O’Sullivan, O.; Crispie, F.; Moloney, R.D.; Cotter, P.D.; Dinan, T.G.; Cryan, J.F. Gut microbiota depletion from early adolescence in mice: Implications for brain and behaviour. Brain Behav. Immun. 2015, 48, 165–173. [Google Scholar] [CrossRef]
- Karl, T.; Pabst, R.; von Hörsten, S. Behavioral phenotyping of mice in pharmacological and toxicological research. Exp. Toxicol. Pathol. 2003, 55, 69–83. [Google Scholar] [CrossRef]
- Rodina, V.I.; Krupina, N.A.; Kryzhanovskiĭ, G.N.; Oknina, N.B. A multiparameter method for the complex evaluation of anxiety-phobic states in rats. Zhurnal Vyss. Nervn. Deiatelnosti Im. IP Pavlova 1993, 43, 1006–1017. (In Russian) [Google Scholar]
- Deacon, R.M.; Rawlins, J.N. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef]
- Wu, C.Y.C.; Lerner, F.M.; Couto E Silva, A.; Possoit, H.E.; Hsieh, T.H.; Neumann, J.T.; Minagar, A.; Lin, H.W.; Lee, R.H.C. Utilizing the modified T-maze to assess functional memory outcomes after cardiac arrest. J. Vis. Exp. 2018, 131, 56694. [Google Scholar] [CrossRef]
- Kumar, M.; Sandhir, R. Hydrogen sulfide suppresses homocysteine-induced glial activation and inflammatory response. Nitric Oxide 2019, 90, 15–28. [Google Scholar] [CrossRef]
- Anisimova, E.A.; Yarullina, D.R. Antibiotic resistance of LACTOBACILLUS strains. Curr. Microbiol. 2019, 76, 1407. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Woollard, A.C.; Wolff, S.P. Hydrogen peroxide production during experimental protein glycation. FEBS Lett. 1990, 268, 69–71. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Langille, M.G.; Meehan, C.J.; Koenig, J.E.; Dhanani, A.S.; Rose, R.A.; Howlett, S.E.; Beiko, R.G. Microbial shifts in the aging mouse gut. Microbiome 2014, 2, 50. [Google Scholar] [CrossRef]
- van der Lugt, B.; Rusli, F.; Lute, C.; Lamprakis, A.; Salazar, E.; Boekschoten, M.V.; Hooiveld, G.J.; Müller, M.; Vervoort, J.; Kersten, S.; et al. Integrative analysis of gut microbiota composition, host colonic gene expression and intraluminal metabolites in aging C57BL/6J mice. Aging 2018, 10, 930–950. [Google Scholar] [CrossRef]
- Campedelli, I.; Mathur, H.; Salvetti, E.; Clarke, S.; Rea, M.C.; Torriani, S.; Ross, R.P.; Hill, C.; O’Toole, P.W. Genus-wide assessment of antibiotic resistance in Lactobacillus spp. Appl. Environ. Microbiol. 2018, 85, e01738-18. [Google Scholar] [CrossRef]
- Anisimova, E.; Gorokhova, I.; Karimullina, G.; Yarullina, D. Alarming antibiotic resistance of lactobacilli isolated from probiotic preparations and dietary supplements. Antibiotics 2022, 11, 1557. [Google Scholar] [CrossRef]
- Weber, E.M.; Zidar, J.; Ewaldsson, B.; Askevik, K.; Udén, E.; Svensk, E.; Törnqvist, E. Aggression in group-housed male mice: A systematic review. Animals 2022, 13, 143. [Google Scholar] [CrossRef]
- Hayer, S.S.; Hwang, S.; Clayton, J.B. Antibiotic-induced gut dysbiosis and cognitive, emotional, and behavioral changes in rodents: A systematic review and meta-analysis. Front. Neurosci. 2023, 17, 1237177. [Google Scholar] [CrossRef]
- Mikkelsen, K.; Hallam, K.; Stojanovska, L.; Apostolopoulos, V. Yeast based spreads improve anxiety and stress. J. Funct. Foods 2018, 40, 471–476. [Google Scholar] [CrossRef]
- Atassi, F.; Servin, A.L. Individual and co-operative roles of lactic acid and hydrogen peroxide in the killing activity of enteric strain Lactobacillus johnsonii NCC933 and vaginal strain Lactobacillus gasseri KS120.1 against enteric, uropathogenic and vaginosis-associated pathogens. FEMS Microbiol. Lett. 2010, 304, 29–38. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, D.C. Facing a new challenge: The adverse effects of antibiotics on gut microbiota and host immunity. Chin. Med. J. 2019, 132, 1135–1138. [Google Scholar] [CrossRef]
- Éliás, A.J.; Barna, V.; Patoni, C.; Demeter, D.; Veres, D.S.; Bunduc, S.; Erőss, B.; Hegyi, P.; Földvári-Nagy, L.; Lenti, K. Probiotic supplementation during antibiotic treatment is unjustified in maintaining the gut microbiome diversity: A systematic review and meta-analysis. BMC Med. 2023, 21, 262. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Mahony, S.M. The microbiome-gut-brain axis: From bowel to behavior. Neurogastroenterol. Motil. 2011, 23, 187–192. [Google Scholar] [CrossRef]
- Huang, C.; Feng, S.; Huo, F.; Liu, H. Effects of four antibiotics on the diversity of the intestinal microbiota. Microbiol. Spectr. 2022, 10, e0190421. [Google Scholar] [CrossRef]
- Wang, P.; Tu, K.; Cao, P.; Yang, Y.; Zhang, H.; Qiu, X.T.; Zhang, M.M.; Wu, X.J.; Yang, H.; Chen, T. Antibiotics-induced intestinal dysbacteriosis caused behavioral alternations and neuronal activation in different brain regions in mice. Mol. Brain 2021, 14, 49. [Google Scholar] [CrossRef]
- Elvers, K.T.; Wilson, V.J.; Hammond, A.; Duncan, L.; Huntley, A.L.; Hay, A.D.; van der Werf, E.T. Antibiotic-induced changes in the human gut microbiota for the most commonly prescribed antibiotics in primary care in the UK: A systematic review. BMJ Open 2020, 10, e035677. [Google Scholar] [CrossRef]
- Shah, T.; Baloch, Z.; Shah, Z.; Cui, X.; Xia, X. The intestinal microbiota: Impacts of antibiotics therapy, colonization resistance, and diseases. Int. J. Mol. Sci. 2021, 22, 6597. [Google Scholar] [CrossRef] [PubMed]
- Beck, B.R.; Park, G.S.; Jeong, D.Y.; Lee, Y.H.; Im, S.; Song, W.H.; Kang, J. Multidisciplinary and comparative investigations of potential psychobiotic effects of Lactobacillus strains isolated from newborns and their impact on gut microbiota and ileal transcriptome in a healthy murine model. Front. Cell Infect. Microbiol. 2019, 9, 269. [Google Scholar] [CrossRef]
- Cai, Y.; Kumai, S.; Ogawa, M.; Benno, Y.; Nakase, T. Characterization and identification of Pediococcus species isolated from forage crops and their application for silage preparation. Appl. Environ. Microbiol. 1999, 65, 2901–2906. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Y.; Hu, A.; Shu, X.; Huang, W.; Liu, J.; Wang, B.; Zhang, R.; Yue, M.; Yang, C. Lactobacillus plantarum-derived postbiotics prevent Salmonella-induced neurological dysfunctions by modulating gut-brain axis in mice. Front. Nutr. 2022, 9, 946096. [Google Scholar] [CrossRef]
- Hijová, E. Postbiotics as metabolites and their biotherapeutic potential. Int. J. Mol. Sci. 2024, 25, 5441. [Google Scholar] [CrossRef]
- Mittal, R.; Debs, L.H.; Patel, A.P.; Nguyen, D.; Patel, K.; O’Connor, G.; Grati, M.; Mittal, J.; Yan, D.; Eshraghi, A.A.; et al. Neurotransmitters: The critical modulators regulating gut-brain axis. J. Cell Physiol. 2017, 232, 2359–2372. [Google Scholar] [CrossRef]
- Oleskin, A.; Zhilenkova, O.; Shenderov, B.; Amerhanova, A.; Kudrin, V.; Klodt, P. Lactic-acid bacteria supplement fermented dairy products with human behavior-modifying neuroactive compounds. J. Pharm. Nutr. Sci. 2014, 4, 199–206. [Google Scholar] [CrossRef]
- Dumitrescu, L.; Popescu-Olaru, I.; Cozma, L.; Tulbă, D.; Hinescu, M.E.; Ceafalan, L.C.; Gherghiceanu, M.; Popescu, B.O. Oxidative stress and the microbiota-gut-brain axis. Oxid. Med. Cell Longev. 2018, 2018, 2406594. [Google Scholar] [CrossRef]
- Patel, J.C.; Rice, M.E. Classification of H2O2 as a neuromodulator that regulates striatal dopamine release on a subsecond time scale. ACS Chem. Neurosci. 2012, 3, 991–1001. [Google Scholar] [CrossRef]
- Lee, C.R.; Patel, J.C.; O’Neill, B.; Rice, M.E. Inhibitory and excitatory neuromodulation by hydrogen peroxide: Translating energetics to information. J. Physiol. 2015, 593, 3431–3446. [Google Scholar] [CrossRef] [PubMed]
- Rio, M.; Velez-Pardo, C. The hydrogen peroxide and its importance in Alzheimer’s and Parkinson’s disease. Curr. Med. Chem. Cent. Nerv. Syst. Agents 2004, 4, 279–285. [Google Scholar] [CrossRef]
- Marin, I.A.; Goertz, J.E.; Ren, T.; Rich, S.S.; Onengut-Gumuscu, S.; Farber, E.; Wu, M.; Overall, C.C.; Kipnis, J.; Gaultier, A. Microbiota alteration is associated with the development of stress-induced despair behavior. Sci. Rep. 2017, 7, 43859. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Suárez, J.E. Biosynthesis and degradation of H2O2 by vaginal lactobacilli. Appl. Environ. Microbiol. 2010, 76, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Hertzberger, R.; Arents, J.; Dekker, H.L.; Pridmore, R.D.; Gysler, C.; Kleerebezem, M.; de Mattos, M.J. H2O2 production in species of the Lactobacillus acidophilus group: A central role for a novel NADH-dependent flavin reductase. Appl. Environ. Microbiol. 2014, 80, 2229–2239. [Google Scholar] [CrossRef] [PubMed]
- Pascual, L.M.; Daniele, M.B.; Pájaro, C.; Barberis, L. Lactobacillus species isolated from the vagina: Identification, hydrogen peroxide production and nonoxynol-9 resistance. Contraception 2006, 73, 78–81. [Google Scholar] [CrossRef]
- Campana, R.; van Hemert, S.; Baffone, W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathog. 2017, 9, 12. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days | Total Microbial Growth | Lactobacillus spp. | Enterobacteria | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lactose-Positive | Lactose-Negative | |||||||||||
| CON | AB | AB + LB | CON | AB | AB + LB | CON | AB | AB + LB | CON | AB | AB + LB | |
| 1 | 13.7 ± 0.6 | 13.7 ± 0.6 | 13.7 ± 0.6 | 15.3 ± 1.2 | 15.3 ± 1.2 | 15.3 ± 1.2 | 11.5 ± 1.8 | 11.5 ± 1.8 | 11.5 ± 1.8 | 1.7 ± 0.2 | 1.7 ± 0.2 | 1.7 ± 0.2 |
| 8 | 15.5 ± 1.4 | 15.2 ± 0.2 | 15.6 ± 1.3 | 15.9 ± 1.4 | 15.9 ± 1.4 | 15.9 ± 1.4 | 6.4 ± 0.9 * | 5.8 ± 0.9 * | 3.6 ± 0.5 * | 6.8 ± 1.1 * | 7.2 ± 0.8 * | 8.7 ± 1.1 * |
| 15 | 15.4 ± 1.5 | 14.9 ± 1.4 | 15.4 ± 1.5 | 15.6 ± 1.4 | 15.6 ± 1.4 | 15.6 ± 1.5 | 5.9 ± 0.8 * | 5.2 ± 0.9 * | 6.1 ± 0.7 * | 6.9 ± 1.0 * | 7.9 ± 0.8 * | 7.1 ± 1.0 * |
| Lactobacillus Strain | Antimicrobial, µg/Disk 1 | H2O2 Production, µM 2 | Antioxidant Activity, % 2 | ||||
|---|---|---|---|---|---|---|---|
| Amphotericin-B, 40 | Ampicillin, 10 | Metronidazol, 5 | Neomycin, 30 | Vancomycin, 30 | |||
| L. plantarum 8PA3 | 0 (R) | 43 (S) | 0 (R) | 23 (S) $ | 21 (S) $ | 58.0 ± 5.0 | 20.5 ± 5.8 $ |
| L. rhamnosus 12L | 0 (R) | 22 (S) | 0 (R) | 15 (I) $ | 0 (R) $ | 60.7 ± 10.3 | 13.1 ± 4.5 $ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yarullina, D.; Novoselova, V.; Alexandrova, A.; Arslanova, A.; Yakovleva, O.; Shaidullov, I.; Nikolaev, Y.; El-Registan, G.; Kudrin, V.; Sitdikova, G. Probiotic Lactobacilli Ameliorate Antibiotic-Induced Cognitive and Behavioral Impairments in Mice. Microbiol. Res. 2024, 15, 1471-1485. https://doi.org/10.3390/microbiolres15030099
Yarullina D, Novoselova V, Alexandrova A, Arslanova A, Yakovleva O, Shaidullov I, Nikolaev Y, El-Registan G, Kudrin V, Sitdikova G. Probiotic Lactobacilli Ameliorate Antibiotic-Induced Cognitive and Behavioral Impairments in Mice. Microbiology Research. 2024; 15(3):1471-1485. https://doi.org/10.3390/microbiolres15030099
Chicago/Turabian StyleYarullina, Dina, Vera Novoselova, Anastasia Alexandrova, Alisa Arslanova, Olga Yakovleva, Ilnar Shaidullov, Yury Nikolaev, Galina El-Registan, Vladimir Kudrin, and Guzel Sitdikova. 2024. "Probiotic Lactobacilli Ameliorate Antibiotic-Induced Cognitive and Behavioral Impairments in Mice" Microbiology Research 15, no. 3: 1471-1485. https://doi.org/10.3390/microbiolres15030099
APA StyleYarullina, D., Novoselova, V., Alexandrova, A., Arslanova, A., Yakovleva, O., Shaidullov, I., Nikolaev, Y., El-Registan, G., Kudrin, V., & Sitdikova, G. (2024). Probiotic Lactobacilli Ameliorate Antibiotic-Induced Cognitive and Behavioral Impairments in Mice. Microbiology Research, 15(3), 1471-1485. https://doi.org/10.3390/microbiolres15030099