
Screening of Microorganisms from Wastes and Identification of the Optimal Substrate for Biosurfactant Production

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Waste Sampling
2.2. Isolation of Microorganisms from Wastes
2.3. Assessment of Biosurfactant Production Ability
2.4. Determination of Strain Species
2.5. Cultivation of Isolates on Different Waste Types
2.6. Extraction of Biosurfactants
2.7. Statistical Analysis
3. Results and Discussion
3.1. Isolation of Biosurfactant-Producing Microorganisms from Waste
3.2. Evaluation of Isolates for Their Surface Tension Reduction Abilities
3.3. Evaluation of Emulsifying Properties of Isolates
3.4. Biosurfactants Yield
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohanty, S.S.; Koul, Y.; Varjani, S.; Pandey, A.; Ngo, H.H.; Chang, J.S.; Wong, J.W.C.; Bui, X.T. A Critical Review on Various Feedstocks as Sustainable Substrates for Biosurfactants Production: A Way towards Cleaner Production. Microb. Cell Fact. 2021, 20, 120. [Google Scholar] [CrossRef] [PubMed]
- Galitskaya, P.; Karamova, K.; Biktasheva, L.; Galieva, G.; Gordeev, A.; Selivanovskaya, S. Lipopeptides Produced by Bacillus Mojavensis P1709 as an Efficient Tool to Maintain Postharvest Cherry Tomato Quality and Quantity. Agriculture 2022, 12, 609. [Google Scholar] [CrossRef]
- Khopade, A.; Biao, R.; Liu, X.; Mahadik, K.; Zhang, L.; Kokare, C. Production and Stability Studies of the Biosurfactant Isolated from Marine Nocardiopsis sp. B4. Desalination 2012, 285, 198–204. [Google Scholar] [CrossRef]
- Carolin, C.F.; Senthil Kumar, P.; Mohanakrishna, G.; Hemavathy, R.V.; Rangasamy, G.; M Aminabhavi, T. Sustainable Production of Biosurfactants via Valorisation of Industrial Wastes as Alternate Feedstocks. Chemosphere 2023, 312, 137326. [Google Scholar] [CrossRef] [PubMed]
- Gaur, V.K.; Sharma, P.; Sirohi, R.; Varjani, S.; Taherzadeh, M.J.; Chang, J.-S.; Yong Ng, H.; Wong, J.W.C.; Kim, S.-H. Production of Biosurfactants from Agro-Industrial Waste and Waste Cooking Oil in a Circular Bioeconomy: An Overview. Bioresour. Technol. 2022, 343, 126059. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Patil, Y.; Rale, V. Biosurfactant Production: Emerging Trends and Promising Strategies. J. Appl. Microbiol. 2019, 126, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Aparna, A.; Srinikethan, G.; Smitha, H. Production and Characterization of Biosurfactant Produced by a Novel Pseudomonas sp. 2B. Colloids Surf. B Biointerfaces 2012, 95, 23–29. [Google Scholar] [CrossRef]
- Moussa, T.A.A.; Mohamed, M.S.; Samak, N. Production and Characterization of Di-Rhamnolipid Produced by Pseudomonas Aeruginosa TMN. Braz. J. Chem. Eng. 2014, 31, 867–880. [Google Scholar] [CrossRef]
- Bonmatin, J.-M.; Laprevote, O.; Peypoux, F. Diversity Among Microbial Cyclic Lipopeptides: Iturins and Surfactins. Activity-Structure Relationships to Design New Bioactive Agents. Comb. Chem. High Throughput Screen. 2012, 6, 541–556. [Google Scholar] [CrossRef]
- Das, P.; Mukherjee, S.; Sen, R. Substrate Dependent Production of Extracellular Biosurfactant by a Marine Bacterium. Bioresour. Technol. 2009, 100, 1015–1019. [Google Scholar] [CrossRef]
- Singh, A.K.; Rautela, R.; Cameotra, S.S. Substrate Dependent in Vitro Antifungal Activity of Bacillus Sp Strain AR2. Microb. Cell Fact. 2014, 13, 67. [Google Scholar] [CrossRef] [PubMed]
- Mouafo, T.H.; Mbawala, A.; Ndjouenkeu, R. Effect of Different Carbon Sources on Biosurfactants’ Production by Three Strains of Lactobacillus spp. Biomed Res. Int. 2018, 2018, 5034783. [Google Scholar] [CrossRef] [PubMed]
- Ndlovu, T.; Rautenbach, M.; Khan, S.; Khan, W. Variants of Lipopeptides and Glycolipids Produced by Bacillus Amyloliquefaciens and Pseudomonas Aeruginosa Cultured in Different Carbon Substrates. AMB Express 2017, 7, 109. [Google Scholar] [CrossRef]
- Santos, D.; Rufino, R.; Luna, J.; Santos, V.; Sarubbo, L. Biosurfactants: Multifunctional Biomolecules of the 21st Century. Int. J. Mol. Sci. 2016, 17, 401. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh-Sani, M.; Hamishehkar, H.; Khezerlou, A.; Azizi-Lalabadi, M.; Azadi, Y.; Nattagh-Eshtivani, E.; Fasihi, M.; Ghavami, A.; Aynehchi, A.; Ehsani, A. Bioemulsifiers Derived from Microorganisms: Applications in the Drug and Food Industry. Adv. Pharm. Bull. 2018, 8, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Uzoigwe, C.; Burgess, J.G.; Ennis, C.J.; Rahman, P.K.S.M. Bioemulsifiers Are Not Biosurfactants and Require Different Screening Approaches. Front. Microbiol. 2015, 6, 245. [Google Scholar] [CrossRef] [PubMed]
- Stoimenova, E.; Vasileva-Tonkova, E.; Sotirova, A.; Galabova, D.; Lalchev, Z. Evaluation of Different Carbon Sources for Growth and Biosurfactant Production by Pseudomonas Fluorescens Isolated from Wastewaters. Z. Fur Naturforschung Sect. C J. Biosci. 2009, 64, 96–102. [Google Scholar] [CrossRef]
- Rane, A.N.; Baikar, V.V.; Ravi Kumar, D.V.; Deopurkar, R.L. Agro-Industrial Wastes for Production of Biosurfactant by Bacillus Subtilis ANR 88 and Its Application in Synthesis of Silver and Gold Nanoparticles. Front. Microbiol. 2017, 8, 492. [Google Scholar] [CrossRef]
- Cooper, D.G.; Goldenberg, B.G. Surface-Active Agents from Two Bacillus Species. Appl. Environ. Microbiol. 1987, 53, 224–229. [Google Scholar] [CrossRef]
- Yin, H.; Qiang, J.; Jia, Y.; Ye, J.; Peng, H.; Qin, H.; Zhang, N.; He, B. Characteristics of Biosurfactant Produced by Pseudomonas Aeruginosa S6 Isolated from Oil-Containing Wastewater. Process Biochem. 2009, 44, 302–308. [Google Scholar] [CrossRef]
- Ludwig, W. Nucleic Acid Techniques in Bacterial Systematics and Identification. Int. J. Food Microbiol. 2007, 120, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Waghmare, P.R.; Patil, S.M.; Jadhav, S.L.; Jeon, B.H.; Govindwar, S.P. Utilization of Agricultural Waste Biomass by Cellulolytic Isolate Enterobacter sp. SUK-Bio. Agric. Nat. Resour. 2018, 52, 399–406. [Google Scholar] [CrossRef]
- Pei, X.H.; Zhan, X.H.; Wang, S.M.; Lin, Y.S.; Zhou, L.X. Effects of a Biosurfactant and a Synthetic Surfactant on Phenanthrene Degradation by a Sphingomonas Strain. Pedosphere 2010, 20, 771–779. [Google Scholar] [CrossRef]
- Berg, G. Rhizobacteria of Oilseed Rape Antagonistic to Verticillium Dahliae Var. Longisporum STARK. Z Pflanzenkr Pflanzenschutz 1996, 103, 20–30. [Google Scholar]
- Kalvandi, S.; Garousin, H.; Pourbabaee, A.A.; Farahbakhsh, M. The Release of Petroleum Hydrocarbons from a Saline-Sodic Soil by the New Biosurfactant-Producing Strain of Bacillus sp. Sci. Rep. 2022, 12, 19770. [Google Scholar] [CrossRef] [PubMed]
- Montecillo, J.A.V.; Bae, H. Reclassification of Brevibacterium Frigoritolerans as Peribacillus Frigoritolerans Comb. Nov. Based on Phylogenomics and Multiple Molecular Synapomorphies. Int. J. Syst. Evol. Microbiol. 2022, 72, 005389. [Google Scholar] [CrossRef]
- Khondee, N.; Ruamyat, N.; Luepromchai, E.; Sikhao, K.; Hawangchu, Y. Bioconversion of Lignocellulosic Wastes to Zwitterionic Biosurfactants by an Alkaliphilic Bacterium: Process Development and Product Characterization. Biomass Bioenergy 2022, 165, 106568. [Google Scholar] [CrossRef]
- Manetsberger, J.; Caballero Gómez, N.; Soria-Rodríguez, C.; Benomar, N.; Abriouel, H. Simply Versatile: The Use of Peribacillus Simplex in Sustainable Agriculture. Microorganisms 2023, 11, 2540. [Google Scholar] [CrossRef]
- Ibrahim, H.M.M. Characterization of Biosurfactants Produced by Novel Strains of Ochrobactrum Anthropi HM-1 and Citrobacter Freundii HM-2 from Used Engine Oil-Contaminated Soil. Egypt. J. Pet. 2018, 27, 21–29. [Google Scholar] [CrossRef]
- Renard, P.; Canet, I.; Sancelme, M.; Wirgot, N.; Deguillaume, L.; Delort, A.M. Screening of Cloud Microorganisms Isolated at the Puy de Dôme (France) Station for the Production of Biosurfactants. Atmo. Chem. Phys. 2016, 16, 12347–12358. [Google Scholar] [CrossRef]
- Darwesh, O.M.; Mahmoud, M.S.; Barakat, K.M.; Abuellil, A.; Ahmad, M.E. Improving the Bioremediation Technology of Contaminated Wastewater Using Biosurfactants Produced by Novel Bacillus Isolates. Heliyon 2021, 7, e08616. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, L.K.; Tamang, B.; Tirwa, R.K.; Lepcha, P.L. Influence of Biosurfactant Producing Bacillus Tequilensis LK5.4 Isolate of Kinema, a Fermented Soybean, on Seed Germination and Growth of Maize (Zea mays L.). 3 Biotech 2020, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; Rubinovitz, C.; Gottlieb, A.; Rosenhak, S.; Ron, E.Z. Production of Biodispersan by Acinetobacter Calcoaceticus A2. Appl. Environ. Microbiol. 1988, 54, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Mendes Lopes, E. Emulsification Properties of Bioemulsifiers Produced by Wild-Type and Mutant Bradyrhizobium Elkanii Strains. J. Bioremediat. Biodegrad. 2014, 5, 1000245. [Google Scholar] [CrossRef]
- Rabiei, A.; Sharifinik, M.; Niazi, A.; Hashemi, A.; Ayatollahi, S. Core Flooding Tests to Investigate the Effects of IFT Reduction and Wettability Alteration on Oil Recovery during MEOR Process in an Iranian Oil Reservoir. Appl. Microbiol. Biotechnol. 2013, 97, 5979–5991. [Google Scholar] [CrossRef] [PubMed]
- Essghaier, B.; Mallat, N.; Khwaldia, K.; Mottola, F.; Rocco, L.; Hannachi, H. Production and Characterization of New Biosurfactants/Bioemulsifiers from Pantoea Alhagi and Their Antioxidant, Antimicrobial and Anti-Biofilm Potentiality Evaluations. Molecules 2023, 28, 1912. [Google Scholar] [CrossRef] [PubMed]
- Sen, R. Response Surface Optimization of the Critical Media Components for the Production of Surfactin. J. Chem. Technol. Biotechnol. 1997, 68, 263–270. [Google Scholar] [CrossRef]
- Jiménez-Peñalver, P.; Koh, A.; Gross, R.; Gea, T.; Font, X. Biosurfactants from Waste: Structures and Interfacial Properties of Sophorolipids Produced from a Residual Oil Cake. J. Surfactants Deterg. 2020, 23, 481–486. [Google Scholar] [CrossRef]
- Ciurko, D.; Czyżnikowska, Ż.; Kancelista, A.; Łaba, W.; Janek, T. Sustainable Production of Biosurfactant from Agro-Industrial Oil Wastes by Bacillus Subtilis and Its Potential Application as Antioxidant and ACE Inhibitor. Int. J. Mol. Sci. 2022, 23, 10824. [Google Scholar] [CrossRef]
- Umar, A.A.; Saaid, I.B.M.; Sulaimon, A.A.; Pilus, R.B.M. A Review of Petroleum Emulsions and Recent Progress on Water-in-Crude Oil Emulsions Stabilized by Natural Surfactants and Solids. J. Pet. Sci. Eng. 2018, 165, 673–690. [Google Scholar] [CrossRef]
- Saimmai, A.; Rukadee, O.; Onlamool, T.; Sobhon, V.; Maneerat, S. Isolation and Functional Characterization of a Biosurfactant Produced by a New and Promising Strain of Oleomonas Sagaranensis AT18. World J. Microbiol. Biotechnol. 2012, 28, 2973–2986. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Yeh, K.L.; Lu, W.B.; Lin, C.L.; Chang, J.S. Rhamnolipid Production with Indigenous Pseudomonas Aeruginosa EM1 Isolated from Oil-Contaminated Site. Bioresour. Technol. 2008, 99, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Das, A.J.; Kumar, R. Utilization of Agro-Industrial Waste for Biosurfactant Production under Submerged Fermentation and Its Application in Oil Recovery from Sand Matrix. Bioresour. Technol. 2018, 260, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Liepins, J.; Balina, K.; Soloha, R.; Berzina, I.; Lukasa, L.K.; Dace, E. Glycolipid Biosurfactant Production from Waste Cooking Oils by Yeast: Review of Substrates, Producers and Products. Fermentation 2021, 7, 136. [Google Scholar] [CrossRef]
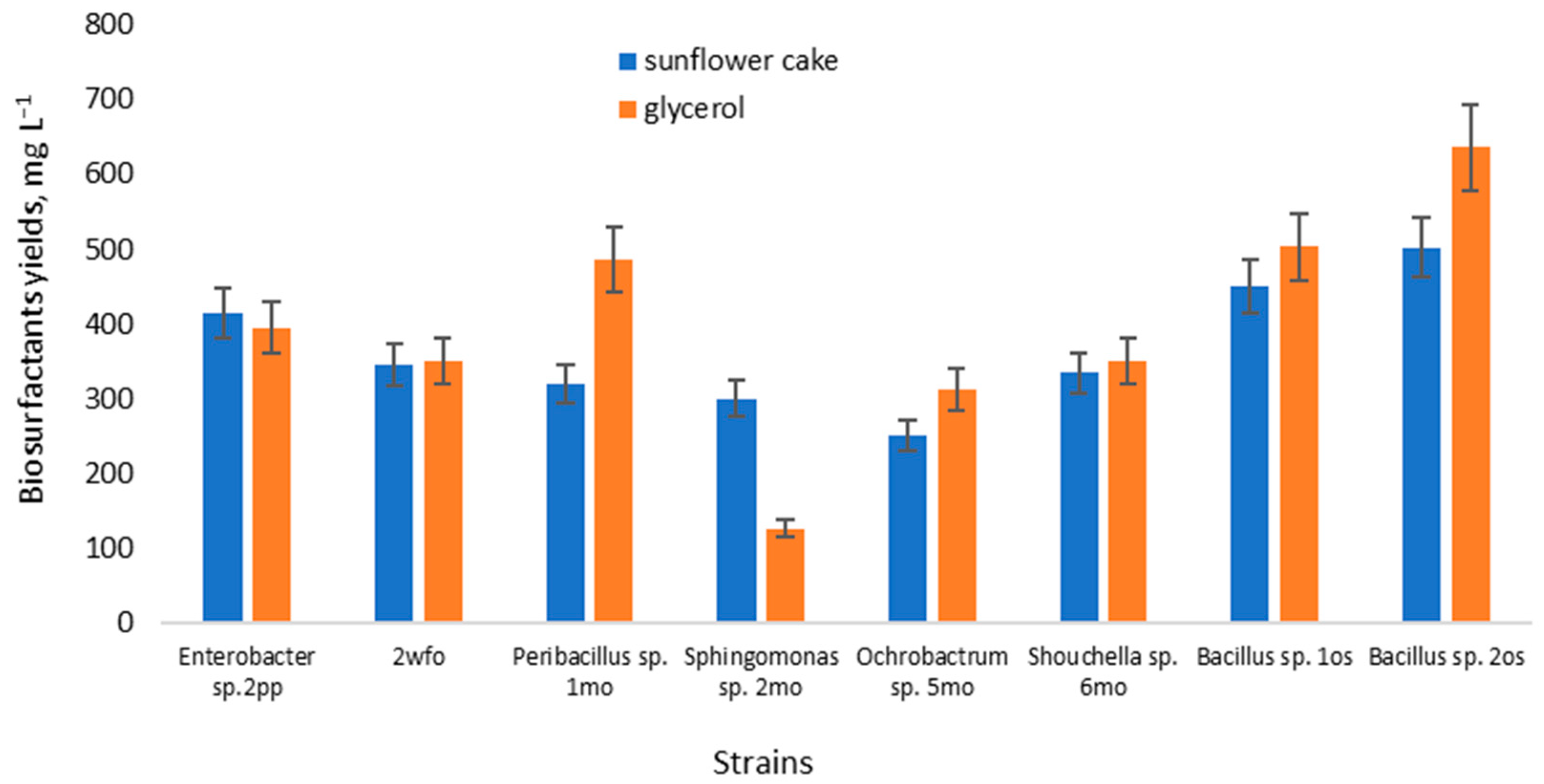

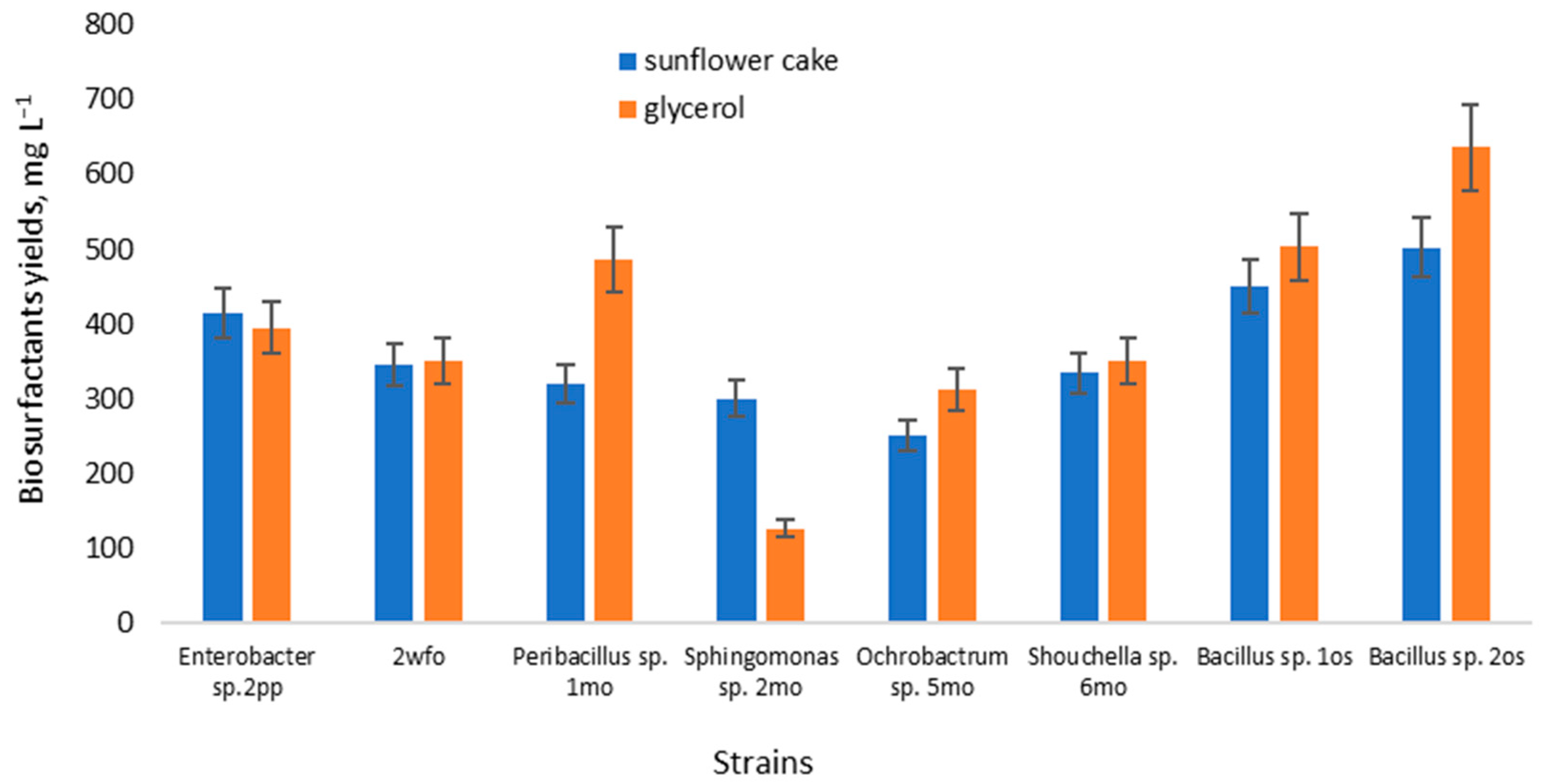{kind=link}
| Wastes | Strains | The Closest-Related Strain | Identity, % | Identified as |
|---|---|---|---|---|
| potato peelings | 1pp | Enterobacter sp. strain XN81 | 98.5 | Enterobacter sp. 1pp |
| 2pp | Enterobacter ludwigii strain E8-13 | 97.6 | Enterobacter sp. 2pp | |
| 3pp | Bacterium strain BS0657 | 93.4 | 3pp | |
| 4pp | Uncultured bacterium clone RBL10-19 | 94.9 | 4pp | |
| 5pp | Enterobacter ludwigii strain 160-a blue | 98.7 | Enterobacter sp. 5pp | |
| rapeseed cake | 1rc | Pantoea sp. LL69 | 83.6 | Pantoea sp. 1rc |
| grease trap | 1gt | Proteus mirabilis, isolate AHI-2 | 99.5 | Proteus sp. 1gt |
| 2gt | Bacillus aerophilus strain 0125 | 97.4 | Bacillus sp. 2gt | |
| oily sludge | 1s | Citrobacter freundii strain E51 | 99.4 | Citrobacter sp. 1s |
| 2s | Sphingomonas echinoides strain B18 | 94.7 | Sphingomonas sp. 2s | |
| waste frying oil | 1wfo | Sphingomonas sp. strain FKP374 | 97.5 | Sphingomonas sp. 1wfo |
| 2wfo | Uncultured Sphingomonas sp., clone: LR564B-24 | 89.8 | 2wfo | |
| 3wfo | Pseudomonas stutzeri DSM 10701 | 99.6 | Pseudomonas sp. 3wfo | |
| 4wfo | Uncultured Sphingomonas sp., clone: LR564B-24 | 95.4 | Sphingomonas sp. 4wfo | |
| 5wfo | Sphingomonas echinoides strain KCOM 3301 (=JS364) | 91.6 | Sphingomonas sp. 5wfo | |
| sunflower cake | 1sc | Sphingomonas sp. strain FKP374 | 94.6 | Sphingomonas sp. 1sc |
| 2sc | Sphingomonas sp. Hc_01N 16S | 90.4 | Sphingomonas sp. 2sc | |
| 3sc | Bacterium strain BLEC3 | 89.5 | 3sc | |
| 4sc | Uncultured Sphingomonas sp., clone: LR564B-24 | 87.4 | Sphingomonas sp. 4sc | |
| 5sc | Sphingomonas sp. PP-2 16S | 83.9 | Sphingomonas sp. 5sc | |
| soil contaminated with motor oil | 1mo | Peribacillus frigoritolerans strain TG15 | 94.6 | Peribacillus sp. 1mo |
| 2mo | Sphingomonas echinoides strain MERYL5-24 | 94.1 | Sphingomonas sp. 2mo | |
| 3mo | Nocardiopsis sp. XLI-8 | 93.9 | Nocardiopsis sp. 3mo | |
| 4mo | Bacillus gibsonii strain S-2 | 95.0 | Bacillus sp. 4mo | |
| 5mo | Ochrobactrum sp. strain S2n90 | 81.2 | Ochrobactrum sp. 5mo | |
| 6mo | Shouchella gibsonii strain LMITABS00983 | 95.0 | Shouchella sp. 6mo | |
| 7mo | Sphingomonas sp. strain MEREH12 | 83.6 | Sphingomonas sp. 7mo | |
| oil contaminated soil | 1os | Bacillus tequilensis strain RS53 | 99.6 | Bacillus sp. 1os |
| 2os | Bacillus toyonensis strain FORT 102 | 99.1 | Bacillus sp. 2os | |
| 3os | Bacillus amyloliquefaciens strain Sihong_838_1 | 81.7 | Bacillus sp. 3os | |
| 4os | Staphylococcus sp. strain FKR3-1 | 90.8 | Staphylococcus sp. 4os | |
| 5os | Bacillus cereus strain 2-2 16S | 76.6 | Bacillus sp. 5os | |
| 6os | Sphingomonas sp. strain SA4_1 | 83.6 | Sphingomonas sp. 6os | |
| 7os | Sphingomonas sp. strain BWLP17 | 87.6 | Sphingomonas sp. 7os | |
| 8os | Sphingomonas sp. strain MERYL1-1 | 87.3 | Sphingomonas sp. 8os | |
| 9os | Uncultured Sphingomonas sp., clone: LR564B-24 | 83.6 | Sphingomonas sp. 9os | |
| 10os | Uncultured bacterium, clone SIP12-RT-12 | 86.8 | 10os |
| Type of Wastes | Strains | ST, mN m−1 | |||
|---|---|---|---|---|---|
| Glycerol | Potato Peelings | Waste Frying Oil | Sunflower Cake | ||
| Potato peelings | Enterobacter sp. 1pp | 58.05 ± 0.681 | 54.75 ± 0.415 | 63.28 ± 0.453 | 55.66± 0.269 |
| Enterobacter sp. 2pp | 43.84 ± 0.206 | 56.73 ± 0.486 | 43.41± 0.136 | 51.96± 0.269 | |
| 3pp | 52.86 ± 0.599 | 56.42 ± 0.442 | 47.99± 0.198 | 59.03± 0.234 | |
| 4pp | 45.06 ± 0.198 | 55.99 ± 0.441 | 47.74± 0.187 | 46.53± 0.190 | |
| Enterobacter sp. 5pp | 49.58 ± 0.069 | 61.93 ± 0.305 | 53.96± 0.354 | 51.22± 0.56 | |
| rapeseed oil | Pantoea sp. 1rc | 43.41 ± 0.263 | 53.81 ± 0.328 | 52.57± 0.512 | 49.8± 0.345 |
| grease trap | Proteus sp. 1gt | 65.14 ± 0.695 | 61.58 ± 0.269 | 48.37± 0.353 | 58.61± 0.61 |
| Bacillus sp. 2gt | 65.95 ± 0.438 | 60.27 ± 0.361 | 56.25± 0.553 | 62.09± 0.655 | |
| oily sludge | Citrobacter sp. 1s | 58.59 ± 1.419 | 52.52 ± 0.225 | 48.37± 0.335 | 53.36± 0.489 |
| Sphingomonas sp. 2s | 66.53 ± 0.545 | 52.11 ± 0.315 | 56.25± 0.256 | 58.99± 0.605 | |
| waste frying oil | Sphingomonas sp. 1wfo | 53.53 ± 0.433 | 47.33 ± 0.120 | 49.82± 0.487 | 50.45± 0.180 |
| Sphingomonas sp. 2wfo | 54.27 ± 0.508 | 27.83 ± 0.033 | 47.87± 0.303 | 29.78± 0.03 | |
| Pseudomonas sp. 2wfo | 55.64 ± 0.488 | 49.70 ± 0.169 | 43.04± 0.233 | 58.81± 0.61 | |
| Sphingomonas sp. 4wfo | 60.34 ± 0.578 | 53.60 ± 0.598 | 54.13± 0.321 | 55.75± 0.614 | |
| Sphingomonas sp. 5wfo | 55.93 ± 0.596 | 57.91 ± 0.501 | 54.8± 0.567 | 58.32± 0.576 | |
| Sunflower cake | Sphingomonas sp. 1sc | 69.4 ± 0.288 | 62.02 ± 1.073 | 63.56± 0.679 | 65.34± 0.678 |
| Sphingomonas sp. 2sc | 69.29 ± 0.284 | 64.62 ± 0.761 | 66.45± 0.665 | 65.12± 0.608 | |
| 3sc | 27.83 ± 0.034 | 59.09 ± 1.147 | 60.34± 0.64 | 65.73± 0.603 | |
| Sphingomonas sp. 4sc | 68.14 ± 0.351 | 63.89 ± 0.914 | 64.87± 0.677 | 65.99± 0.555 | |
| Sphingomonas sp. 5sc | 35.78 ± 0.352 | 67.87 ± 0.604 | 65.45± 0.501 | 63.39± 0.679 | |
| soil contaminated with motor oil | Peribacillus sp. 1mo | 21.64± 0.018 | 45.76 ± 0.174 | 43.56± 0.344 | 57.57± 0.456 |
| Sphingomonas sp. 2mo | 39.71± 0.03 | 50.34 ± 0.379 | 36.77± 0.09 | 28.32± 0.05 | |
| Nocardiopsis sp. 3mo | 48.61 ± 0.097 | 29.98 ± 0.098 | 45.34± 0.185 | 3.45± 0.025 | |
| Bacillus sp. 4mo | 47.9 ± 0.648 | 64.75 ± 0.654 | 45.1± 0.205 | 52.11± 0.432 | |
| Ochrobactrum sp. 5mo | 29.15 ± 0.077 | 50.85 ± 0.125 | 33.89± 0.1 | 34.49± 0.061 | |
| Shouchella sp. 6mo | 59.15 ± 0.389 | 29.15 ± 0.082 | 42.58± 0.186 | 3.14± 0.02 | |
| Sphingomonas sp. 7mo | 47.88 ± 0.355 | 55.95 ± 0.820 | 48.91± 0.307 | 53.98± 0.399 | |
| oil contaminated soil | Bacillus sp. 1os | 53.45 ± 0.484 | 33.07 ± 0.480 | 45.13± 0.205 | 24.19± 0.056 |
| Bacillus sp. 2os | 60.35 ± 0.564 | 30.86 ± 0.257 | 32.56± 0.11 | 3.99± 0.01 | |
| Bacillus sp. 3os | 50.11 ± 0.494 | 31.02 ± 0.234 | 35.76± 0.101 | 29.17± 0.055 | |
| Staphylococcus sp. 4os | 45.76 ± 0.284 | 53.30 ± 0.384 | 44.55± 0.234 | 49.75± 0.155 | |
| Bacillus sp. 5os | 29.42 ± 0.042 | 28.8 ± 0.090 | 58.63± 0.566 | 28.66± 0.04 | |
| Sphingomonas sp. 6os | 63.43 ± 0.355 | 62.11 ± 0.588 | 61.23± 0.453 | 63.54± 0.666 | |
| Sphingomonas sp. 7os | 64.91 ± 0.566 | 43.37 ± 0.948 | 50.08± 0.334 | 51.1± 0.456 | |
| Sphingomonas sp. 8os | 59.78 ± 0.67 | 49.91 ± 0.142 | 53.87± 0.788 | 53.08± 0.489 | |
| Sphingomonas sp. 9os | 68.08 ± 0.561 | 45.12 ± 0.074 | 45.31± 0.305 | 47.55± 0.311 | |
| 10os | 57.51 ± 0.475 | 50.34 ± 0.086 | 53.45± 0.398 | 55.67± 0.499 | |
 reduction of surface tension to 40 mN m−1 is highlighted in red.
reduction of surface tension to 40 mN m−1 is highlighted in red.| Type of Wastes | Strains | E24, % | |||
|---|---|---|---|---|---|
| Glycerol | Potato Peelings | Waste Frying Oil | Sunflower Cake | ||
| potato peelings | Enterobacter sp. 1pp | 20 ± 1 | 7 ± 3 | 20 ± 5 | 15 ± 5 |
| Enterobacter sp. 2pp | 50 ± 5 | 36 ± 5 | 35 ± 5 | 50 ± 3 | |
| 3pp | 20 ± 2 | 29 ± 3 | 20 ± 2 | 20 ± 1 | |
| 4pp | 50 ± 5 | 14 ± 2 | 20 ± 1 | 21 ± 5 | |
| Enterobacter sp. 5pp | 30 ± 5 | 9 ± 1 | 10 ± 3 | 5 ± 3 | |
| rapeseed oil | Pantoea sp. 1rc | 50 ± 5 | 4 ± 0 | 4 ± 0 | 30 ± 3 |
| grease trap | Proteus sp. 1gt | 5 ± 0 | 21 ± 0 | 10 ± 1 | 10 ± 2 |
| Bacillus sp. 2gt | 0 ± 0 | 7 ± 3 | 5 ± 0 | 5 ± 0 | |
| oily sludge | Citrobacter sp. 1s | 5 ± 0 | 7 ± 4 | 5 ± 0 | 5 ± 0 |
| Sphingomonas sp. 2s | 10 ± 2 | 7 ± 4 | 10 ± 1 | 10 ± 2 | |
| waste frying oil | Sphingomonas sp. 1wfo | 0 ± 0 | 14 ± 5 | 7 ± 2 | 0 ± 0 |
| Sphingomonas sp. 2wfo | 30 ± 2 | 29 ± 5 | 30 ± 2 | 7 ± 4 | |
| Pseudomonas sp. 2wfo | 20 ± 1 | 50 ± 5 | 30 ± 3 | 20 ± 2 | |
| Sphingomonas sp. 4wfo | 5 ± 0 | 14 ± 3 | 10 ± 1 | 5 ± 0 | |
| Sphingomonas sp. 5wfo | 5 ± 0 | 14 ± 2 | 10 ± 2 | 10 ± 3 | |
| sunflower cake | Sphingomonas sp. 1sc | 20 ± 3 | 14 ± 2 | 14 ± 4 | 14 ± 5 |
| Sphingomonas sp. 2sc | 30 ± 5 | 0 ± 0 | 5 ± 0 | 5 ± 0 | |
| 3sc | 50 ± 5 | 14 ± 2 | 30 ± 2 | 21 ± 4 | |
| Sphingomonas sp. 4sc | 10 ± 2 | 14 ± 3 | 10 ± 1 | 14 ± 5 | |
| Sphingomonas sp. 5sc | 20 ± 1 | 14 ± 4 | 14 ± 2 | 29 ± 6 | |
| soil contaminated with motor oil | Peribacillus sp. 1mo | 50 ± 3 | 14 ± 0 | 50 ± 3 | 50 ± 2 |
| Sphingomonas sp. 2mo | 50 ± 2 | 14 ± 0 | 50 ± 2 | 71 ± 4 | |
| Nocardiopsis sp. 3mo | 30 ± 1 | 14 ± 1 | 7 ± 1 | 7 ± 2 | |
| Bacillus sp. 4mo | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| Ochrobactrum sp. 5mo | 50 ± 5 | 14 ± 2 | 20 ± 2 | 21 ± 1 | |
| Shouchella sp. 6mo | 30 ± 5 | 29 ± 3 | 29 ± 2 | 7 ± 3 | |
| Sphingomonas sp. 7mo | 10 ± 2 | 50 ± 3 | 30 ± 1 | 20 ± 5 | |
| oil contaminated soil | Bacillus sp. 1os | 50 ± 2 | 29 ± 5 | 50 ± 1 | 57 ± 5 |
| Bacillus sp. 2os | 5 ± 0 | 50 ± 1 | 30 ± 4 | 7 ± 0 | |
| Bacillus sp. 3os | 20 ± 0 | 21 ± 5 | 20 ± 1 | 14 ± 2 | |
| Staphylococcus sp. 4os | 5 ± 0 | 14 ± 3 | 5 ± 0 | 5 ± 0 | |
| Bacillus sp. 5os | 20 ± 0 | 0 ± 0 | 10 ± 0 | 14 ± 3 | |
| Sphingomonas sp. 6os | 30 ± 2 | 14 ± 2 | 10 ± 0 | 10 ± 3 | |
| Sphingomonas sp. 7os | 10 ± 1 | 14 ± 1 | 10 ± 2 | 10 ± 4 | |
| Sphingomonas sp. 8os | 30 ± 3 | 14 ± 3 | 20 ± 3 | 20 ± 2 | |
| Sphingomonas sp. 9os | 20 ± 1 | 29 ± 5 | 20 ± 5 | 20 ± 1 | |
| 10os | 20 ± 1 | 50 ± 2 | 20 ± 1 | 20 ± 4 | |
E24 > 50% are highlighted in red.Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biktasheva, L.; Gordeev, A.; Kirichenko, A.; Kuryntseva, P.; Selivanovskaya, S. Screening of Microorganisms from Wastes and Identification of the Optimal Substrate for Biosurfactant Production. Microbiol. Res. 2024, 15, 152-163. https://doi.org/10.3390/microbiolres15010010
Biktasheva L, Gordeev A, Kirichenko A, Kuryntseva P, Selivanovskaya S. Screening of Microorganisms from Wastes and Identification of the Optimal Substrate for Biosurfactant Production. Microbiology Research. 2024; 15(1):152-163. https://doi.org/10.3390/microbiolres15010010
Chicago/Turabian StyleBiktasheva, Liliya, Alexander Gordeev, Anastasia Kirichenko, Polina Kuryntseva, and Svetlana Selivanovskaya. 2024. "Screening of Microorganisms from Wastes and Identification of the Optimal Substrate for Biosurfactant Production" Microbiology Research 15, no. 1: 152-163. https://doi.org/10.3390/microbiolres15010010
APA StyleBiktasheva, L., Gordeev, A., Kirichenko, A., Kuryntseva, P., & Selivanovskaya, S. (2024). Screening of Microorganisms from Wastes and Identification of the Optimal Substrate for Biosurfactant Production. Microbiology Research, 15(1), 152-163. https://doi.org/10.3390/microbiolres15010010