
Biocide Syntheses Bee Venom-Conjugated ZnO@αFe2O3 Nanoflowers as an Advanced Platform Targeting Multidrug-Resistant Fecal Coliform Bacteria Biofilm Isolated from Treated Wastewater

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
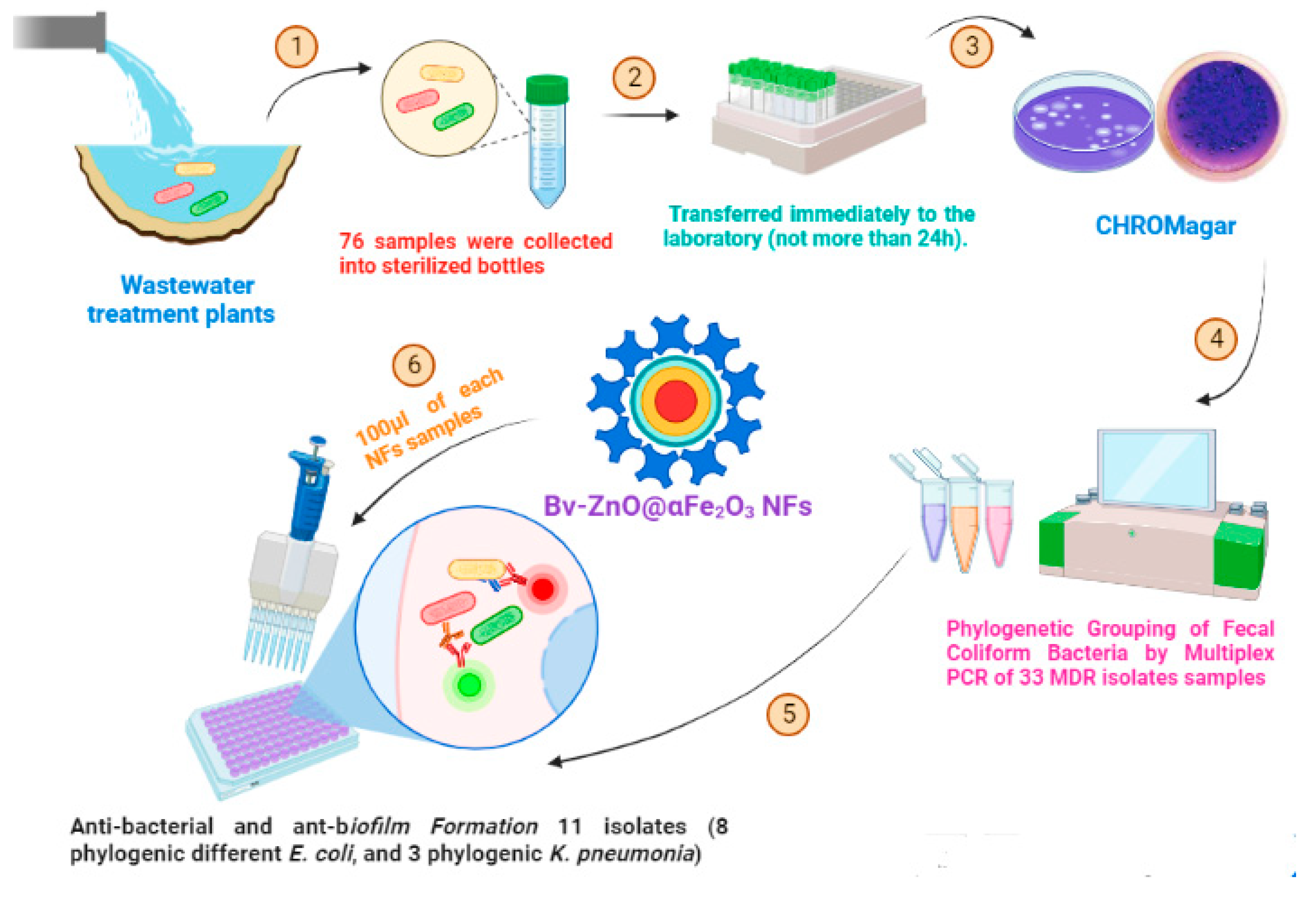
2. Materials and Methods
2.1. Bee Venom Collection
2.2. Bv-Collecting Device
2.3. Bee Venom Collection
2.4. Isolation of Fecal Coliform Bacteria from Treated Wastewater Samples
2.5. Detection of Isolates’ Multidrug-Resistant Fecal Coliform
2.6. Phylogenetic Grouping of Fecal Coliform Bacteria by Multiplex PCR
2.7. Preparation of ZnO@αFe2O3 NFs
2.8. Bee Venom-Coated ZnO@αFe2O3 NFs
2.9. Characterizations of Prepared NFs
2.10. Cell Viability and Cytotoxicity Study of Prepared NFs
2.11. Antibacterial Susceptibility Assay of Prepared NFs
2.12. Determination of MIC and MBC of NFs against Tested Isolates of Bacteria
2.13. Screening of MDR Isolates for Biofilm Formation by Tissue Culture Plate Method
2.14. In Vitro Evaluation of Anti-Biofilm Activity of NFs
2.15. Scanning Electron Microscopy (SEM)
2.16. Statistical Analysis
3. Results
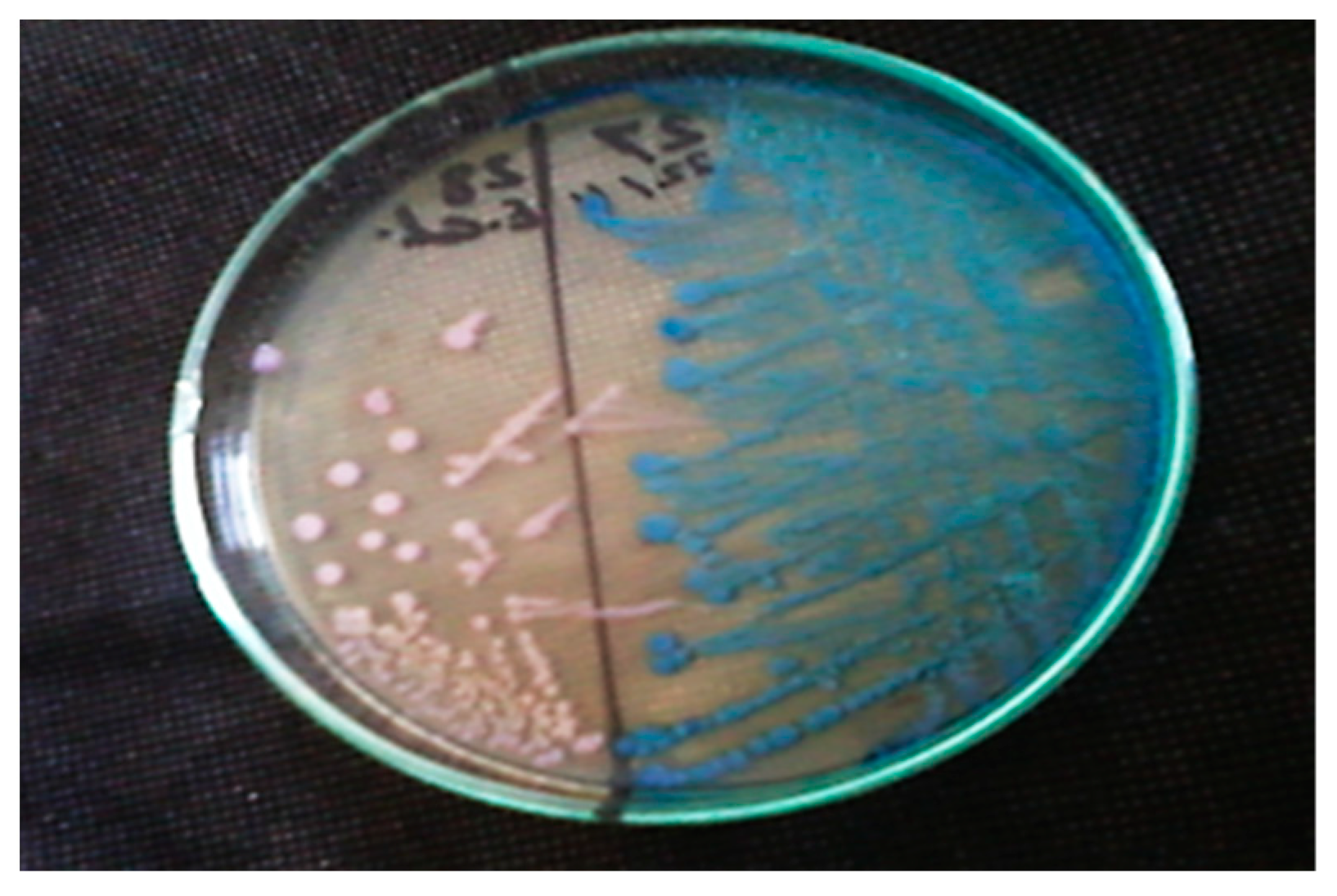
3.1. Culture and Identification of Fecal Coliform Bacteria from Treated Wastewater Samples
3.2. Detection of Antibiotic Susceptibility Pattern of Fecal Isolates
3.3. Phylogenetic Grouping by Multiplex PCR
3.4. Characterization of Prepared NFs
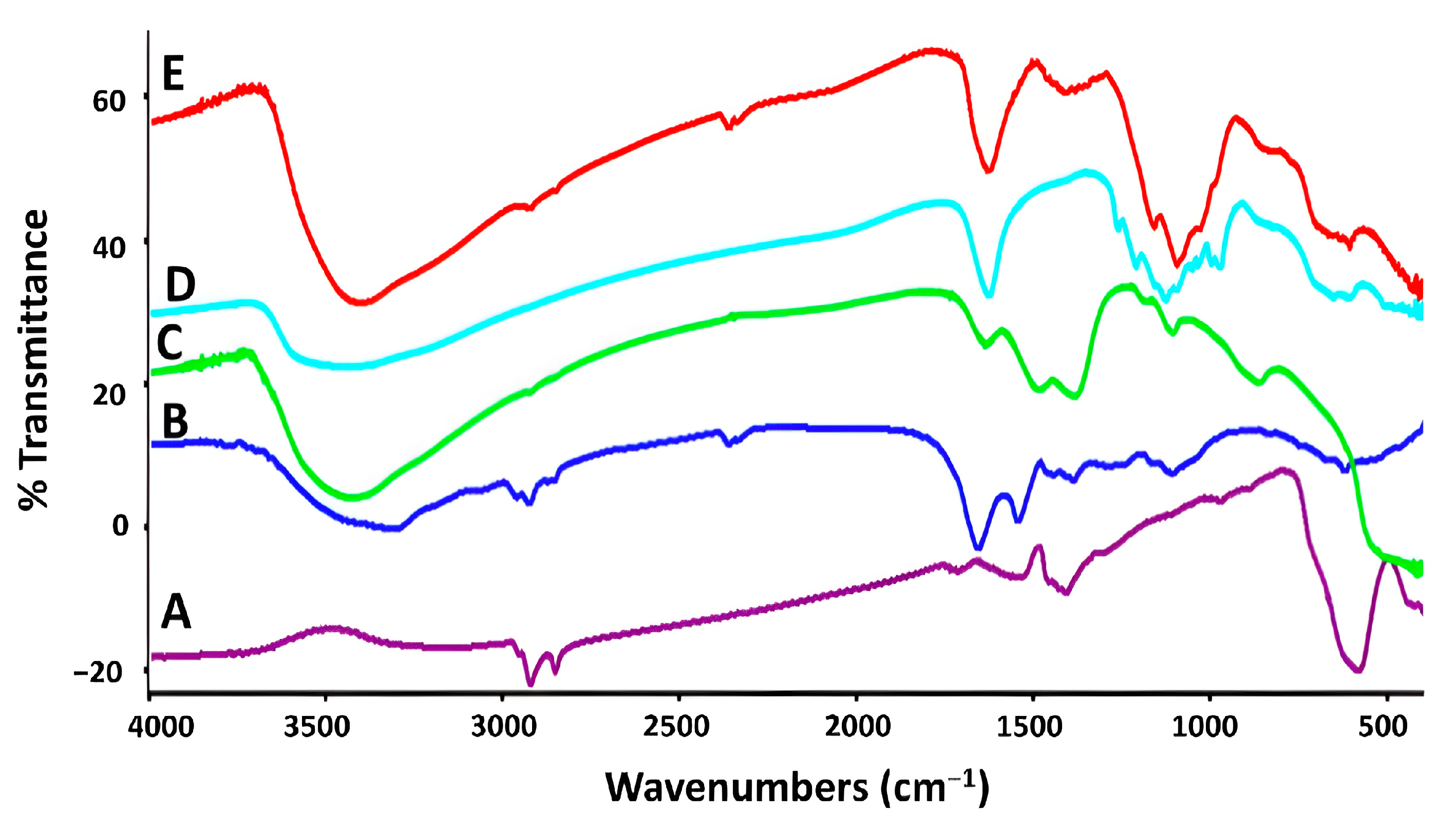
3.4.1. FTIR Spectra
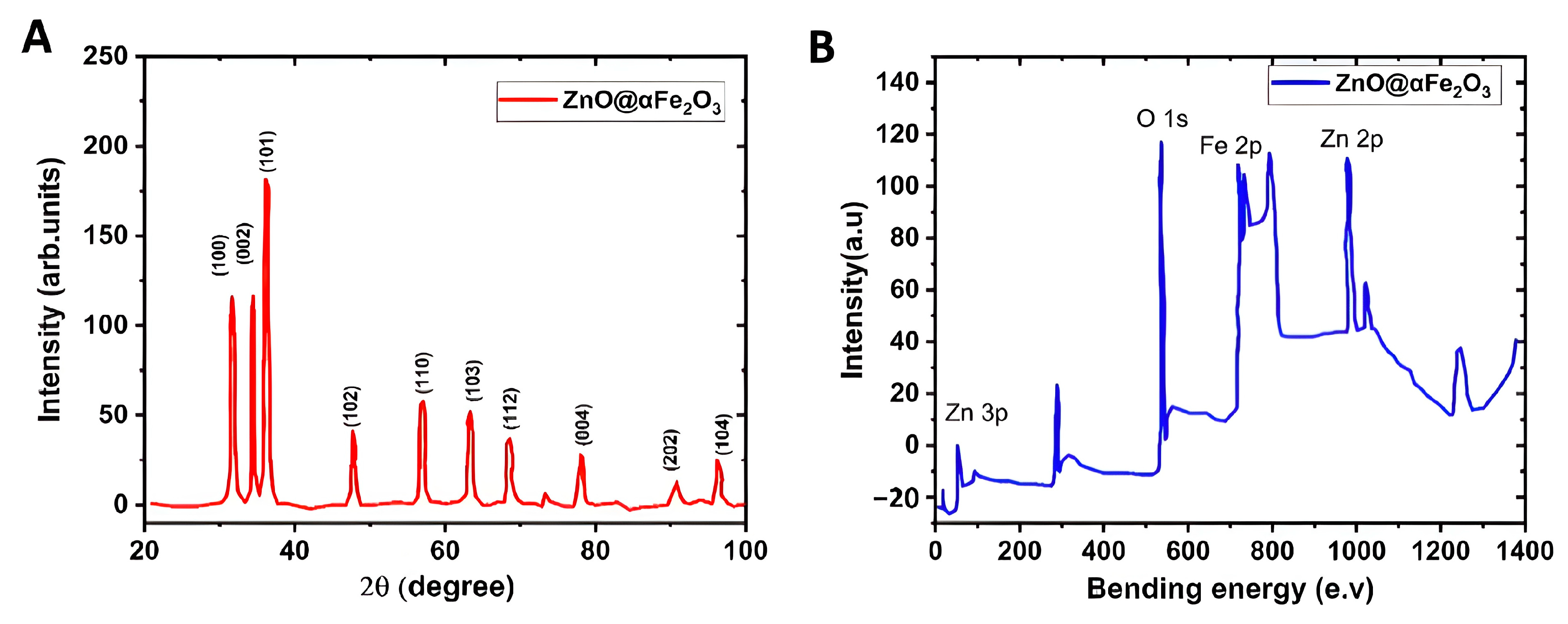
3.4.2. XRD and XPS
3.5. Surface Morphology Analysis
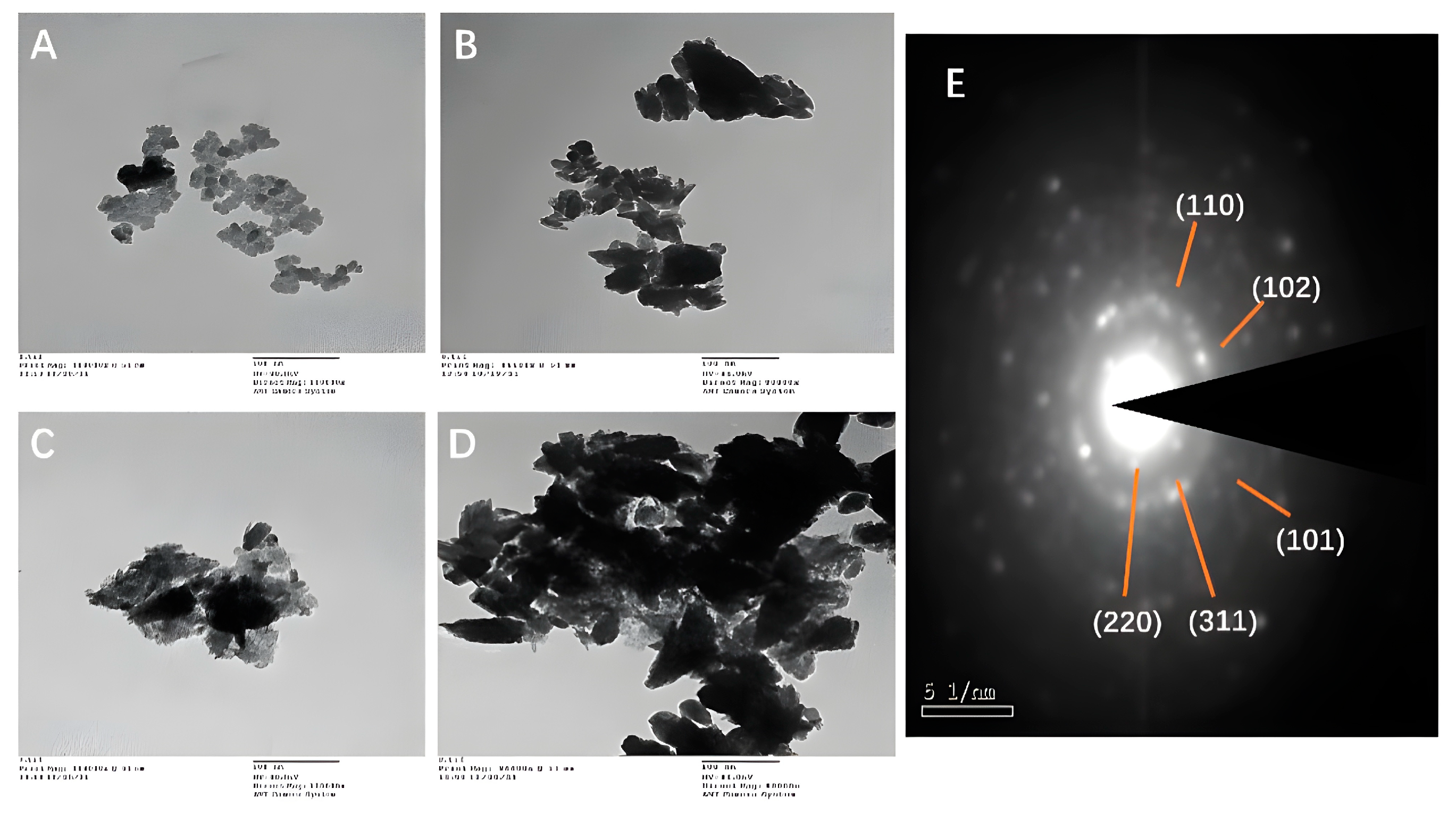
3.5.1. TEM and Selected Area Electron Diffraction (SAED)
3.5.2. SEM-EDX and HRTEM
3.6. Cell Viability and Cytotoxicity of NPs and NFs
3.7. Antibacterial Susceptibility Assay of NPs and NFs
3.8. MIC and MBC Determination of NFs
3.9. Biofilm Formation of Isolates and Anti-Biofilm Activity of NPs and NFs
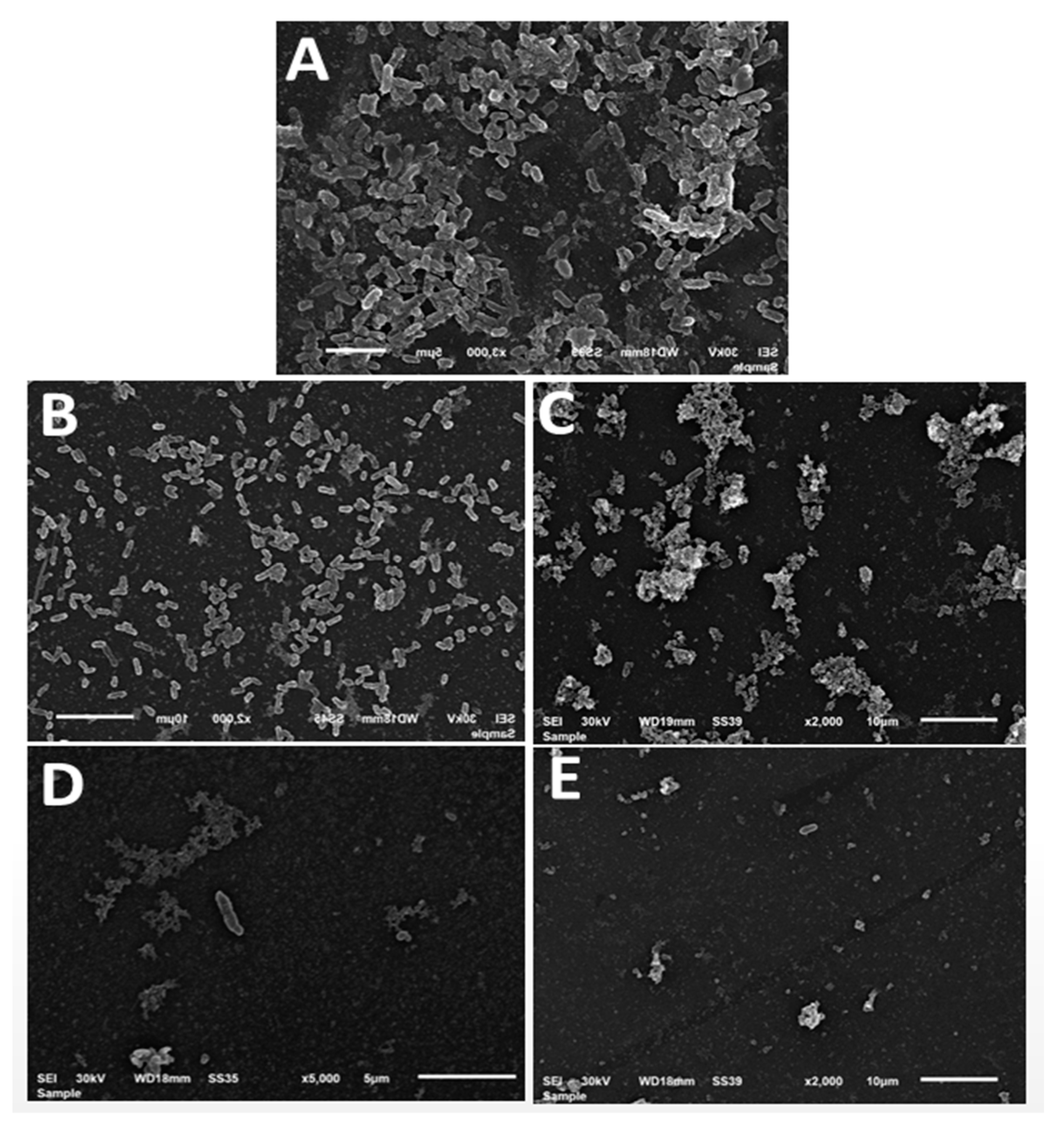
3.10. Biofilm Inhibition for E. coli by SEM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Escobar-Páramo, P.; Le Menac’h, A.; Le Gall, T.; Amorin, C.; Gouriou, S.; Picard, B.; Skurnik, D.; Denamur, E. Identification of forces shaping the commensal Escherichia coli genetic structure by comparing animal and human isolates. Environ. Microbiol. 2006, 8, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Wegener, H.C.; Collignon, P. Resistance in bacteria of the food chain: Epidemiology and control strategies. Expert Rev. Anti-Infect. Ther. 2008, 6, 733–750. [Google Scholar] [PubMed]
- Alsanie, W.F.; Felemban, E.M.; Farid, M.A.; Hassan, M.M.; Sabry, A.; Gaber, A. Molecular identification and phylogenetic analysis of multidrug-resistant bacteria using 16S rDNA sequencing. J. Pure Appl. Microbiol. 2018, 12, 489–496. [Google Scholar] [CrossRef]
- Tesh, S.J.; Scott, T.B. Iron nanoparticles for water treatment: Is the future free or fixed? Iron oxides: From nature to applications. In Iron Oxides: From Nature to Applications; Wiley-VCH: Weinheim, Germany, 2016; pp. 473–522. [Google Scholar]
- Badshah, I.; Mustafa, N.; Khan, R.; Mashwani, Z.-U.-R.; Raja, N.I.; Almutairi, M.H.; Aleya, L.; Sayed, A.A.; Zaman, S.; Sawati, L.; et al. Biogenic Titanium Dioxide Nanoparticles Ameliorate the Effect of Salinity Stress in Wheat Crop. Agronomy 2023, 13, 352. [Google Scholar]
- Shin, S.-H.; Ye, M.-K.; Choi, S.-Y.; Park, K.-K. The effects of melittin and apamin on airborne fungi-induced chemical mediator and extracellular matrix production from nasal polyp fibroblasts. Toxins 2017, 9, 348. [Google Scholar] [CrossRef] [PubMed]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.; Cunha, W.R.; Almeida, R.D.; Martins, C.H. Antimicrobial activity of apitoxin, melittin and phospholipase A 2 of honey bee (Apis mellifera) venom against oral pathogens. An. Acad. Bras. Ciências 2015, 87, 147–155. [Google Scholar] [CrossRef]
- Socarras, K.M.; Theophilus, P.A.; Torres, J.P.; Gupta, K.; Sapi, E. Antimicrobial activity of bee venom and melittin against Borrelia burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, T.; Rima, M.; Karam, M.; Sabatier, J.-M.; Fajloun, Z. Antimicrobials from venomous animals: An overview. Molecules 2020, 25, 2402. [Google Scholar] [CrossRef]
- Neves, R.C.D.; Mortari, M.R.; Schwartz, E.F.; Kipnis, A.; Junqueira-Kipnis, A.P. Antimicrobial and antibiofilm effects of peptides from venom of social Wasp and scorpion on multidrug-resistant Acinetobacter baumannii. Toxins 2019, 11, 216. [Google Scholar] [CrossRef]
- Purushothaman, S.; Nair, R.R.; Harikrishnan, V.; Fernandez, A. Temporal relation of cardiac hypertrophy, oxidative stress, and fatty acid metabolism in spontaneously hypertensive rat. Mol. Cell. Biochem. 2011, 351, 59–64. [Google Scholar] [CrossRef]
- Weis, W.A.; Ripari, N.; Conte, F.L.; da Silva Honorio, M.; Sartori, A.A.; Matucci, R.H.; Sforcin, J.M. An overview about apitherapy and its clinical applications. Phytomed. Plus 2022, 2, 100239. [Google Scholar]
- Pereira, A.F.M.; Albano, M.; Alves, F.C.B.; Andrade, B.F.M.T.; Furlanetto, A.; Rall, V.L.M.; Santos, L.D.D.; Orsi, R.D.O.; Junior, A.F. Influence of apitoxin and melittin from Apis mellifera bee on Staphylococcus aureus strains. Microb. Pathog. 2020, 141, 104011. [Google Scholar] [CrossRef] [PubMed]
- Carpena, M.; Nuñez-Estevez, B.; Soria-Lopez, A.; Simal-Gandara, J. Bee venom: An updating review of its bioactive molecules and its health applications. Nutrients 2020, 12, 3360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, Y.; Ye, Y.; Wang, X.-R.; Lin, L.-T.; Xiao, L.-Y.; Zhou, P.; Shi, G.-X.; Liu, C.-Z. Bee venom therapy: Potential mechanisms and therapeutic applications. Toxicon 2018, 148, 64–73. [Google Scholar]
- Kocyigit, A.; Guler, E.M.; Kaleli, S. Anti-inflammatory and antioxidative properties of honey bee venom on Freund’s Complete Adjuvant-induced arthritis model in rats. Toxicon 2019, 161, 4–11. [Google Scholar]
- Sobral, F.; Sampaio, A.; Falcão, S.; Queiroz, M.J.R.; Calhelha, R.C.; Vilas-Boas, M.; Ferreira, I.C. Chemical characterization, antioxidant, anti-inflammatory and cytotoxic properties of bee venom collected in Northeast Portugal. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef]
- Han, S.; Yeo, J.; Baek, H.; Lin, S.-M.; Meyer, S.; Molan, P. Postantibiotic effect of purified melittin from honeybee (Apis mellifera) venom against Escherichia coli and Staphylococcus aureus. J. Asian Nat. Prod. Res. 2009, 11, 796–804. [Google Scholar] [CrossRef]
- Debnath, S.; Sharma, D.; Chaudhari, S.Y.; Sharma, R.; Shaikh, A.A.; Buchade, R.S.; Kesari, K.K.; Abdel-Fattah, A.M.; Algahtani, M.; Mheidat, M.; et al. Wheat ergot fungus-derived and modified drug for inhibition of intracranial aneurysm rupture due to dysfunction of TLR-4 receptor in Alzheimer’s disease. PLoS ONE 2023, 18, e0279616. [Google Scholar] [CrossRef]
- Akash, S.; Baeza, J.; Mahmood, S.; Mukerjee, N.; Subramaniyan, V.; Islam, M.R.; Gupta, G.; Rajakumari, V.; Chinni, S.V.; Ramachawolran, G.; et al. Development of a new drug candidate for the inhibition of Lassa virus glycoprotein and nucleoprotein by modification of evodiamine as promising therapeutic agents. Front. Microbiol. 2023, 14, 1206872. [Google Scholar] [CrossRef]
- Rayaprolu, B.M.; Strawser, J.J.; Strawser, G. Anyarambhatla, i. pharmacy, Excipients in parenteral formulations: Selection considerations and effective utilization with small molecules and biologics. Drug Dev. Ind. Pharm. 2018, 44, 1565–1571. [Google Scholar] [CrossRef]
- Vargason, A.M.; Anselmo, A.C.; Mitragotri, S. The evolution of commercial drug delivery technologies. Nat. Biomed. Eng. 2021, 5, 951–967. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Celi, N.; Zhang, D.; Cai, J. Magnetic biohybrid microrobot multimers based on chlorella cells for enhanced targeted drug delivery. ACS Appl. Mater. Interfaces 2022, 14, 6320–6330. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Sharaf, M.; Ahmed, I.; Khan, T.U.; Shabana, S.; Arif, M.; Kazmi, S.S.U.H.; Liu, C. Potential utility of nano-based treatment approaches to address the risk of Helicobacter pylori. Expert Rev. Anti-Infect. Ther. 2022, 20, 407–424. [Google Scholar] [CrossRef]
- Slavin, Y.N.; Asnis, J.; Häfeli, U.O.; Bach, H. Metal nanoparticles: Understanding the mechanisms behind antibacterial activity. J. Nanobiotechnol. 2017, 15, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Alshawwa, S.Z.; Mohammed, E.J.; Hashim, N.; Sharaf, M.; Selim, S.; Alhuthali, H.M.; Alzahrani, H.A.; Mekky, A.E.; Elharrif, M.G. In Situ Biosynthesis of Reduced Alpha Hematite (α-Fe2O3) Nanoparticles by Stevia Rebaudiana L. Leaf Extract: Insights into Antioxidant, Antimicrobial, and Anticancer Properties. Antibiotics 2022, 11, 1252. [Google Scholar] [CrossRef]
- Javidi, S.; Nafchi, A.M.; Moghadam, H.H. Synergistic effect of nano-ZnO and Mentha piperita essential oil on the moisture sorption isotherm, antibacterial activity, physicochemical, mechanical, and barrier properties of gelatin film. J. Food Meas. Charact. 2022, 16, 964–974. [Google Scholar] [CrossRef]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- Kahlmeter, G.; Giske, C.G.; Kirn, T.J.; Sharp, S.E. Point-counterpoint: Differences between the European Committee on Antimicrobial Susceptibility Testing and Clinical and Laboratory Standards Institute recommendations for reporting antimicrobial susceptibility results. J. Clin. Microbiol. 2019, 57, e01129-19. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef]
- Singla, M.; Gupta, S.; Porwal, O.; Binjawhar, D.N.; Sayed, A.A.; Mittal, P.; El-Demerdash, F.M.; Algahtani, M.; Singh, S.K.; Dua, K.; et al. Theoretical design for covering Engeletin with functionalized nanostructure-lipid carriers as neuroprotective agents against Huntington’s disease via the nasal-brain route. Front. Pharmacol. 2023, 14, 1218625. [Google Scholar]
- Sharaf, M.; Sewid, A.H.; Hamouda, H.; Elharrif, M.G.; El-Demerdash, A.S.; Alharthi, A.; Hashim, N.; Hamad, A.A.; Selim, S.; Alkhalifah, D.H.M. Rhamnolipid-Coated Iron Oxide Nanoparticles as a Novel Multitarget Candidate against Major Foodborne E. coli Serotypes and Methicillin-Resistant S. aureus. Microbiol. Spectr. 2022, 10, e00250-22. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Neoh, K.; Kang, E. Synthesis and in vitro anti-cancer evaluation of tamoxifen-loaded magnetite/PLLA composite nanoparticles. Biomaterials 2006, 27, 5725–5733. [Google Scholar] [CrossRef] [PubMed]
- Kant, R.; Pathak, S.; Dutta, V.J.S.E.M.; Cells, S. Design and fabrication of sandwich-structured α-Fe2O3/Au/ZnO photoanode for photoelectrochemical water splitting. Sol. Energy Mater. Sol. Cells 2018, 178, 38–45. [Google Scholar] [CrossRef]
- Mutz, B.; Belimov, M.; Wang, W.; Sprenger, P.; Serrer, M.-A.; Wang, D.; Pfeifer, P.; Kleist, W.; Grunwaldt, J.-D. Potential of an alumina-supported Ni3Fe catalyst in the methanation of CO2: Impact of alloy formation on activity and stability. ACS Catal. 2017, 7, 6802–6814. [Google Scholar] [CrossRef]
- Magaldi, S.; Mata-Essayag, S.; De Capriles, C.H.; Pérez, C.; Colella, M.; Olaizola, C.; Ontiveros, Y. Well diffusion for antifungal susceptibility testing. Int. J. Infect. Dis. 2004, 8, 39–45. [Google Scholar] [CrossRef]
- Valgas, C.; Souza, S.M.D.; Smânia, E.F.; Jr, A.S. Screening methods to determine antibacterial activity of natural products. Braz. J. Microbiol. 2007, 38, 369–380. [Google Scholar] [CrossRef]
- Salim, M.; Ullah, I.; Saljoqi, A.U.R.; Gökçe, A.; Ahmad, S.; Almutairi, M.H.; Sayed, A.A.; Aleya, L.; Abdel-Daim, M.M.; Shah, M. Life table study of Sitotroga cerealella on different cereals and its implications on the performance of the egg parasitoid (Trichogramma chilonis) under laboratory conditions. Sci. Rep. 2023, 13, 10961. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Jadhav, S.; Shah, R.; Bhave, M.; Palombo, E.A. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control 2013, 29, 125–130. [Google Scholar] [CrossRef]
- Abdallah, F.B.; Lagha, R.; Gaber, A. Biofilm inhibition and eradication properties of medicinal plant essential oils against methicillin-resistant Staphylococcus aureus clinical isolates. Pharmaceuticals 2020, 13, 369. [Google Scholar] [CrossRef]
- Watkinson, A.; Micalizzi, G.; Graham, G.; Bates, J.; Costanzo, S. Antibiotic-resistant Escherichia coli in wastewaters, surface waters, and oysters from an urban riverine system. Appl. Environ. Microbiol. 2007, 73, 5667–5670. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Han, B.; Gu, J.; Wang, C.; Wang, P.; Ma, Y.; Cao, J.; He, Z. Fate of antibiotic resistant cultivable heterotrophic bacteria and antibiotic resistance genes in wastewater treatment processes. Chemosphere 2015, 135, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Zanotto, C.; Bissa, M.; Illiano, E.; Mezzanotte, V.; Marazzi, F.; Turolla, A.; Antonelli, M.; Morghen, C.D.G.; Radaelli, A. Identification of antibiotic-resistant Escherichia coli isolated from a municipal wastewater treatment plant. Chemosphere 2016, 164, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Faried, O.A.; Almuhayawi, M.S.; Saleh, F.M.; Sharaf, M.; El Nahhas, N.; Warrad, M. Incidence of Vancomycin-Resistant Staphylococcus aureus Strains among Patients with Urinary Tract Infections. Antibiotics 2022, 11, 408. [Google Scholar] [CrossRef]
- Salim, M.; Kamran, M.; Khan, I.; Saljoqi, A.U.R.; Ahmad, S.; Almutairi, M.H.; Sayed, A.A.; Aleya, L.; Abdel-Daim, M.M.; Shah, M. Effect of larval diets on the life table parameters of dengue mosquito, Aedes aegypti (L.) (Diptera: Culicidae) using age-stage two sex life table theory. Sci. Rep. 2023, 13, 11969. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Vishwas, S.; Kaur, A.; Kaur, H.; Kakoty, V.; Khursheed, R.; Chaitanya, M.; Babu, M.; Awasthi, A.; Corrie, L.; et al. Harnessing role of sesamol and its nanoformulations against neurodegenerative diseases. Biomed. Pharmacother. 2023, 1, 115512. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, A.S.; Layton, A.C.; Mailloux, B.J.; Culligan, P.J.; Williams, D.E.; Smartt, A.E.; Sayler, G.S.; Feighery, J.; McKay, L.D.; Knappett, P.S. Comparison of fecal indicators with pathogenic bacteria and rotavirus in groundwater. Sci. Total Environ. 2012, 431, 314–322. [Google Scholar] [CrossRef]
- Gilbert, R.; De Louvois, J.; Donovan, T.; Little, C.; Nye, K.; Ribeiro, C.; Richards, J.; Roberts, D.; Bolton, F. Guidelines for the microbiological quality of some ready-to-eat foods sampled at the point of sale. PHLS Advisory Committee for Food and Dairy Products. Commun. Dis. Public Health 2000, 3, 163–167. [Google Scholar]
- Gruber, J.S.; Ercumen, A.; Colford, J.M., Jr. Coliform bacteria as indicators of diarrheal risk in household drinking water: Systematic review and meta-analysis. PLoS ONE 2014, 9, e107429. [Google Scholar] [CrossRef]
- Alouache, S.; Estepa, V.; Messai, Y.; Ruiz, E.; Torres, C.; Bakour, R. Characterization of ESBLs and associated quinolone resistance in Escherichia coli and Klebsiella pneumoniae isolates from an urban wastewater treatment plant in Algeria. Microb. Drug Resist. 2014, 20, 30–38. [Google Scholar] [CrossRef]
- Surleac, M.; Barbu, I.C.; Paraschiv, S.; Popa, L.I.; Gheorghe, I.; Marutescu, L.; Popa, M.; Sarbu, I.; Talapan, D.; Nita, M. Whole genome sequencing snapshot of multi-drug resistant Klebsiella pneumoniae strains from hospitals and receiving wastewater treatment plants in Southern Romania. PLoS ONE 2020, 15, e0228079. [Google Scholar] [CrossRef] [PubMed]
- Loudermilk, E.M.; Kotay, S.M.; Barry, K.E.; Parikh, H.I.; Colosi, L.M.; Mathers, A.J. Tracking Klebsiella pneumoniae carbapenemase gene as an indicator of antimicrobial resistance dissemination from a hospital to surface water via a municipal wastewater treatment plant. Water Res. 2022, 213, 118151. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-Y.; Cai, J.-C.; Zhou, H.-W.; Chi, D.; Zhang, X.-F.; Chen, W.-L.; Zhang, R.; Chen, G.-X. Molecular typing of CTX-M-producing Escherichia coli isolates from environmental water, swine feces, specimens from healthy humans, and human patients. Appl. Environ. Microbiol. 2013, 79, 5988–5996. [Google Scholar] [CrossRef]
- Zhi, S.; Stothard, P.; Banting, G.; Scott, C.; Huntley, K.; Ryu, K.; Otto, S.; Ashbolt, N.; Checkley, S.; Dong, T. Characterization of water treatment-resistant and multidrug-resistant urinary pathogenic Escherichia coli in treated wastewater. Water Res. 2020, 182, 115827. [Google Scholar] [CrossRef]
- Długosz, O.; Szostak, K.; Krupiński, M.; Banach, M. Synthesis of Fe3O4/ZnO nanoparticles and their application for the photodegradation of anionic and cationic dyes. Int. J. Environ. Sci. Technol. 2021, 18, 561–574. [Google Scholar] [CrossRef]
- Smith, R.D.; Prévot, M.S.; Fagan, R.D.; Zhang, Z.; Sedach, P.A.; Siu, M.K.J.; Trudel, S.; Berlinguette, C.P.J.S. Photochemical route for accessing amorphous metal oxide materials for water oxidation catalysis. Science 2013, 340, 60–63. [Google Scholar] [CrossRef]
- Thirunavukkarasu, K.; Sankaranarayanan, T.; Pandurangan, A.; Shanthi, R.V.; Sivasanker, S. The role of surface Zn 2+ ions in the transesterification of vegetable oils over ZnO supported on Al2O3 and Fe2O3. Catal. Sci. Technol. 2014, 4, 851–860. [Google Scholar] [CrossRef]
- Mao, N.J.S.R. Investigating the heteronjunction between ZnO/Fe2O3 and g-C3N4 for an enhanced photocatalytic H2 production under visible-light irradiation. Sci. Rep. 2019, 9, 12383. [Google Scholar] [CrossRef]
- Fakhr, M.A.; Mazrou, Y.S.; Ellmouni, F.Y.; ElSaied, A.; Elhady, M.; Elkelish, A.; Nour, I.H. Investigating the Phenotypic Plasticity of the Invasive Weed Trianthema portulacastrum L. Plants 2022, 11, 77. [Google Scholar] [CrossRef]
- Sawant, V.; Bamane, S. PEG-beta-cyclodextrin functionalized zinc oxide nanoparticles show cell imaging with high drug payload and sustained pH responsive delivery of curcumin in to MCF-7 cells. J. Drug Deliv. Sci. Technol. 2018, 43, 397–408. [Google Scholar] [CrossRef]
- Kumar, S.R.; Paulpandi, M.; ManivelRaja, M.; Mangalaraj, D.; Viswanathan, C.; Kannan, S.; Ponpandian, N. An in vitro analysis of H1N1 viral inhibition using polymer coated superparamagnetic Fe 3 O 4 nanoparticles. RSC Adv. 2014, 4, 13409–13418. [Google Scholar] [CrossRef]
- Alsamadany, H.; Mansour, H.; Elkelish, A.; Ibrahim, M.F.M. Folic Acid Confers Tolerance against Salt Stress-Induced Oxidative Damages in Snap Beans through Regulation Growth, Metabolites, Antioxidant Machinery and Gene Expression. Plants 2022, 30, 1459. [Google Scholar] [CrossRef]
- Rossi, T.; Penfold, T.; Rittmann-Frank, M.; Reinhard, M.; Rittmann, J.; Borca, C.; Grolimund, D.; Milne, C.; Chergui, M. Characterizing the structure and defect concentration of ZnO nanoparticles in a colloidal solution. J. Phys. Chem. C 2014, 118, 19422–19430. [Google Scholar] [CrossRef]
- Rajan, A.S.; Khan, A.; Asrar, S.; Raza, H.; Das, R.K.; Sahu, N.K. Synthesis of ZnO/Fe3 O4/rGO nanocomposites and evaluation of antibacterial activities towards E. coli and S. aureus. IET Nanobiotechnol. 2019, 13, 682–687. [Google Scholar] [CrossRef]
- Bahari, A.; Roeinfard, M.; Ramzannezhad, A.; Khodabakhshi, M.; Mohseni, M. Nanostructured features and antimicrobial properties of Fe3O4/ZnO Nanocomposites. Natl. Acad. Sci. Lett. 2019, 42, 9–12. [Google Scholar] [CrossRef]
- Oprea, O.; Andronescu, E.; Ficai, D.; Ficai, A.; Oktar, F.N.; Yetmez, M. ZnO applications and challenges. Curr. Org. Chem. 2014, 18, 192–203. [Google Scholar] [CrossRef]
- Pradhan, L.; Srivastava, R.; Bahadur, D. pH-and thermosensitive thin lipid layer coated mesoporous magnetic nanoassemblies as a dual drug delivery system towards thermochemotherapy of cancer. Acta Biomater. 2014, 10, 2976–2987. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Burmistrov, D.E.; Serov, D.A.; Rebezov, M.B.; Semenova, A.A.; Lisitsyn, A.B. Do iron oxide nanoparticles have significant antibacterial properties? Antibiotics 2021, 10, 884. [Google Scholar] [CrossRef]
- Sharma, N.; Jandaik, S.; Kumar, S.; Chitkara, M.; Sandhu, I.S. Synthesis, characterisation and antimicrobial activity of manganese-and iron-doped zinc oxide nanoparticles. J. Exp. Nanosci. 2016, 11, 54–71. [Google Scholar] [CrossRef]
- Neupane, B.P.; Chaudhary, D.; Paudel, S.; Timsina, S.; Chapagain, B.; Jamarkattel, N.; Tiwari, B.R. Himalayan honey loaded iron oxide nanoparticles: Synthesis, characterization and study of antioxidant and antimicrobial activities. Int. J. Nanomed. 2019, 14, 3533. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res. 2012, 51, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Čujová, S.; Slaninová, J.; Monincová, L.; Fučík, V.; Bednárová, L.; Štokrová, J.; Hovorka, O.; Voburka, Z.; Straka, J.; Čeřovský, V. Panurgines, novel antimicrobial peptides from the venom of communal bee Panurgus calcaratus (Hymenoptera: Andrenidae). Amino Acids 2013, 45, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, L.-L.; Zhang, A.-T.; Xie, P.; Lu, J.-J.; Zou, X.-T. Antibacterial effects of zinc oxide nanoparticles on Escherichia coli K88. Afr. J. Biotechnol. 2012, 11, 10248–10254. [Google Scholar]
- Yousef, J.M.; Danial, E.N. In vitro antibacterial activity and minimum inhibitory concentration of zinc oxide and nano-particle zinc oxide against pathogenic strains. J. Health Sci. 2012, 2, 38–42. [Google Scholar] [CrossRef]
- Elmi, F.; Alinezhad, H.; Moulana, Z.; Salehian, F.; Tavakkoli, S.M.; Asgharpour, F.; Fallah, H.; Elmi, M.M. The use of antibacterial activity of ZnO nanoparticles in the treatment of municipal wastewater. Water Sci. Technol. 2014, 70, 763–770. [Google Scholar] [CrossRef]
- Samy, R.P.; Gopalakrishnakone, P.; Thwin, M.; Chow, T.; Bow, H.; Yap, E.; Thong, T. Antibacterial activity of snake, scorpion and bee venoms: A comparison with purified venom phospholipase A2 enzymes. J. Appl. Microbiol. 2007, 102, 650–659. [Google Scholar] [CrossRef]
- Hegazi, A.G.; El-Feel, M.; Abdel-Rahman, E.; Al-Fattah, A. Antibacterial activity of bee venom collected from Apis mellifera carniolan pure and hybrid races by two collection methods. Int. J. Curr. Microbiol. App Sci. 2015, 4, 141–149. [Google Scholar]
- Park, S.; Park, B.; Yun, S.; Kang, H.; So, B. Antimicrobial activities of honey bee venom against pathogens isolated from clinical bovine mastitis in Korea. Planta Medica 2013, 79, PL16. [Google Scholar] [CrossRef]
- Fadl, A.E.-W. Antibacterial and antibiofilm effects of bee venom from (Apis mellifera) on multidrug-resistant bacteria (MDRB). Al-Azhar J. Pharm. Sci. 2018, 58, 60–80. [Google Scholar] [CrossRef]
- Marques, S.C.; Rezende, J.D.G.O.S.; Alves, L.A.D.F.; Silva, B.C.; Alves, E.; Abreu, L.R.D.; Piccoli, R.H. Formação de biofilme por Staphylococcus aureus na superfície de aço inoxidável e vidro e sua resistência a alguns sanificantes químicos. Braz. J. Microbiol. 2007, 38, 538–543. [Google Scholar] [CrossRef]
- Batoni, G.; Maisetta, G.; Esin, S. Antimicrobial peptides and their interaction with biofilms of medically relevant bacteria. Biochim. Biophys. Acta BBA-Biomembr. 2016, 1858, 1044–1060. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Tossi, A.; Sandri, L. Giangaspero, Amphipathic, α-helical antimicrobial peptides. Pept. Sci. 2000, 55, 4–30. [Google Scholar] [CrossRef]
- Dosler, S.; Karaaslan, E.; Gerceker, A.A. Antibacterial and anti-biofilm activities of melittin and colistin, alone and in combination with antibiotics against Gram-negative bacteria. J. Chemother. 2016, 28, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Picoli, T.; Peter, C.M.; Zani, J.L.; Waller, S.B.; Lopes, M.G.; Boesche, K.N.; de Oliveira Hübner, S.; Fischer, G. Melittin and its potential in the destruction and inhibition of the biofilm formation by Staphylococcus aureus, Escherichia coli and Pseudomonas aeruginosa isolated from bovine milk. Microb. Pathog. 2017, 112, 57–62. [Google Scholar] [CrossRef]
- Bandara, H.; Nguyen, D.; Mogarala, S.; Osiñski, M.; Smyth, H. Magnetic fields suppress Pseudomonas aeruginosa biofilms and enhance ciprofloxacin activity. Biofouling 2015, 31, 443–457. [Google Scholar] [CrossRef]
- Estevão-Costa, M.-I.; Sanz-Soler, R.; Johanningmeier, B.; Eble, J.A. Snake venom components in medicine: From the symbolic rod of Asclepius to tangible medical research and application. Int. J. Biochem. Cell Biol. 2018, 104, 94–113. [Google Scholar] [CrossRef]
- Ranmadugala, D.; Ebrahiminezhad, A.; Manley-Harris, M.; Ghasemi, Y.; Berenjian, A. The effect of iron oxide nanoparticles on Bacillus subtilis biofilm, growth and viability. Process Biochem. 2017, 62, 231–240. [Google Scholar] [CrossRef]
- Rumyantceva, V.; Rumyantceva, V.; Koshel, E.; Vinogradov, V. Biocide-conjugated magnetite nanoparticles as an advanced platform for biofilm treatment. Ther. Deliv. 2019, 10, 241–250. [Google Scholar] [CrossRef]
- Shakerimoghaddam, A.; Ghaemi, E.A.; Jamalli, A. Zinc oxide nanoparticle reduced biofilm formation and antigen 43 expressions in uropathogenic Escherichia coli. Iran. J. Basic Med. Sci. 2017, 20, 451. [Google Scholar] [PubMed]
- Applerot, G.; Lellouche, J.; Perkas, N.; Nitzan, Y.; Gedanken, A.; Banin, E. ZnO nanoparticle-coated surfaces inhibit bacterial biofilm formation and increase antibiotic susceptibility. Rsc. Adv. 2012, 2, 2314–2321. [Google Scholar] [CrossRef]
- Khan, S.T.; Ahamed, M.; Musarrat, J.; Al-Khedhairy, A.A. Anti-biofilm and antibacterial activities of zinc oxide nanoparticles against the oral opportunistic pathogens R othia dentocariosa and R othia mucilaginosa. Eur. J. Oral Sci. 2014, 122, 397–403. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agent | Disk Content | Interpretive Categories and Zone Diameter Breakpoints | ||
|---|---|---|---|---|
| S | I | R | ||
| Ampicillin (AMP) | 10 μg | 0 | 0 | 100% |
| Piperacillin (PRL) | 30 μg | 0 | 0 | 100% |
| Ofloxacin (OFX) | 5 μg | 0 | 0 | 100% |
| Cefotaxime (CTX) | 10 μg | 0 | 0 | 100% |
| Ceftazidime (CAZ) | 30 μg | 20.4% | 3.7% | 75.9% |
| Cefpodoxime (CPD) | 10/10 μg | 16.7% | 3.7% | 79.6% |
| Cefepime (FEP) | 10 μg | 11.1% | 5.6% | 83.3% |
| Aztreonam (ATM) | 10 μg | 14.8% | 7.4% | 77.8% |
| Ampicillin/sulbactam (AMC) | 20 μg | 14.8% | 3.7% | 81.5% |
| Sulphamethoxazole-trimethoprim (SXT) | 5 μg | 24.1% | 11.1% | 64.8% |
| Chloramphenicol (C) | 30 μg | 48.1% | 22.2% | 29.6% |
| Gentamicin (CN) | 100 μg | 50% | 16.7% | 33.3% |
| Ciprofloxacin (CIP) | 1.25/23.75 μg | 64.8% | 20.4% | 14.8% |
| Meropenem (MEM) | 30 μg | 72.2% | 27.8% | 0% |
| Phylogroup/Subgroup | Number (%) of Subgroup for E. coli | Number (%) of Phylogroup for E. coli | Number (%) of Subgroup for K. pneumoniae | Number (%) of Phylogroup for K. pneumoniae | |
|---|---|---|---|---|---|
| A0 | (chuA−, yjaA−, TspE4.C2−) | 2 | A = 13 (61.9%) | 1 | A = 7 (58.3%) |
| A1 | (chuA−, yjaA+, TspE4.C2−) | 11 | 6 | ||
| B1 | (chuA−, yjaA−, TspE4.C2+) | 1 | B1 = 1 (4.8%) | 1 | B1 = 1 (8.3%) |
| B22 | (chuA+, yjaA+, TspE4.C2−) | 3 | B2 = 5 (23.8%) | 1 | B2 = 4 (33.3%) |
| B23 | (chuA+, yjaA+, TspE4.C2+) | 2 | 3 | ||
| D1 | (chuA+, yjaA−, TspE4.C2−) | 1 | D = 1 (4.8%) | 0 | D = 0 |
| D2 | (chuA+, yjaA−, TspE4.C2+) | 0 | 0 | ||
| Unknown | (chuA−, yjaA+, TspE4.C2+) | 1 | Unknown = 1 (4.7%) | 0 | Unknown = 0 |
| Total | 21 | 63.6% | 12 | 36.4% | |
| Isolate Code | Biofilm Category (Average OD570 ± SD) | Resistance Profile | MAR Index a | Zone of Inhibition of Different Nanoparticles (cm) | |||
|---|---|---|---|---|---|---|---|
| ZnO | ZnO@αFe2O3 | αFe2O3 | Bv-ZnO@αFe2O3 | ||||
| E1 | Weak 0.92 ± 0.014 | Amp, PRL, OFX, CTX, CPD, CIP, STX, ATM, CN | 0.64 | 2 | 3 | 1.8 | 3.8 |
| E2 |
Moderate 1.92 ± 0.024 | Amp, PRL, OFX, CTX, CPD, CIP, ATM, C, CAZ | 0.64 | 1 | 1.5 | 2 | 3 |
| E3 |
Moderate 2.40 ± 0.047 | Amp, PRL, OFX, CTX, STX, CN, C, CAZ, CPD, FEP, ATM | 0.79 | 1 | 1.5 | 2.2 | 3.3 |
| E4 |
Weak 1.19 ± 0.051 | Amp, PRL, OFX, CTX, ATM, AMC, CPD | 0.5 | 1 | 1.8 | 2 | 2.5 |
| E5 |
Strong 3.4 ± 0.141 | Amp, PRL, OFX, CTX, AMC, C, CIP, CAZ, CN, ATM, | 0.71 | 0 | 1.6 | 1.5 | 2.4 |
| E6 |
Moderate 1.75 ± 0.008 | Amp, PRL, OFX, CTX, SXT, CN | 0.42 | 1 | 2.2 | 2 | 3 |
| E7 |
Strong 4.23 ± 0.012 | Amp, PRL, OFX, CTX, CPD, SXT, CIP, C, ATM, AMC, FEP, CAZ, CN | 0.93 | 0.5 | 3.5 | 2.6 | 4 |
| E8 |
Moderate 1.89 ± 0.008 | Amp, PRL, OFX, CTX, AMC, CN | 0.43 | 0 | 1.3 | 1.2 | 1.9 |
| K1 | Weak 1.06 ± 0.029 | Amp, PRL, OFX, CTX, AMC, CIP, ATM, CAZ | 0.57 | 1.8 | 2.4 | 2 | 3.2 |
| K2 |
Moderate 2.61 ± 0.017 | Amp, PRL, OFX, CTX, AMC, C, CN, CAZ | 0.57 | 1 | 3.7 | 1 | 3.9 |
| K3 |
Strong 3.66 ± 0.029 | Amp, PRL, OFX, CTX, CPD, SXT, CAZ, CN, CIP, C, ATM, AMC | 0.85 | 0 | 2.5 | 2 | 4.5 |
| Nanomaterials | E. coli (E7) | K. pneumonia (K3) | ||
|---|---|---|---|---|
| MIC µg/mL−1 | MBC µg/mL−1 | MIC µg/mL−1 | MBC µg/mL−1 | |
| ZnO | 320 | 100 | 160 | 80 |
| αFe2O3 | 80 | 40 | 40 | 10 |
| ZnO@αFe2O3 | 320 | 97 | 80 | 40 |
| Bv-ZnO@αFe2O3 | 10 | 20 | 5 | 2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharaf, M.; Mohammed, E.J.; Farahat, E.M.; Alrehaili, A.A.; Alkhudhayri, A.; Ali, A.M.; Zahra, A.A.; Zakai, S.A.; Elkelish, A.; AlHarbi, M.; et al. Biocide Syntheses Bee Venom-Conjugated ZnO@αFe2O3 Nanoflowers as an Advanced Platform Targeting Multidrug-Resistant Fecal Coliform Bacteria Biofilm Isolated from Treated Wastewater. Microbiol. Res. 2023, 14, 1489-1510. https://doi.org/10.3390/microbiolres14040102
Sharaf M, Mohammed EJ, Farahat EM, Alrehaili AA, Alkhudhayri A, Ali AM, Zahra AA, Zakai SA, Elkelish A, AlHarbi M, et al. Biocide Syntheses Bee Venom-Conjugated ZnO@αFe2O3 Nanoflowers as an Advanced Platform Targeting Multidrug-Resistant Fecal Coliform Bacteria Biofilm Isolated from Treated Wastewater. Microbiology Research. 2023; 14(4):1489-1510. https://doi.org/10.3390/microbiolres14040102
Chicago/Turabian StyleSharaf, Mohamed, Eman Jassim Mohammed, Eman M. Farahat, Amani A. Alrehaili, Abdulsalam Alkhudhayri, Ahmed Mohamed Ali, Abdullah A. Zahra, Shadi A. Zakai, Amr Elkelish, Maha AlHarbi, and et al. 2023. "Biocide Syntheses Bee Venom-Conjugated ZnO@αFe2O3 Nanoflowers as an Advanced Platform Targeting Multidrug-Resistant Fecal Coliform Bacteria Biofilm Isolated from Treated Wastewater" Microbiology Research 14, no. 4: 1489-1510. https://doi.org/10.3390/microbiolres14040102
APA StyleSharaf, M., Mohammed, E. J., Farahat, E. M., Alrehaili, A. A., Alkhudhayri, A., Ali, A. M., Zahra, A. A., Zakai, S. A., Elkelish, A., AlHarbi, M., & Saad, M. F. (2023). Biocide Syntheses Bee Venom-Conjugated ZnO@αFe2O3 Nanoflowers as an Advanced Platform Targeting Multidrug-Resistant Fecal Coliform Bacteria Biofilm Isolated from Treated Wastewater. Microbiology Research, 14(4), 1489-1510. https://doi.org/10.3390/microbiolres14040102