1. Introduction
According to the International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics, postbiotics are defined as preparations of nonliving microorganisms and/or their components that have a health benefit for the host [
1]. In addition to inactivated nonliving cells, postbiotics include metabolic byproducts released from living cells and enzymes, peptides, teichoic acids, muropeptides, polysaccharides, and cell surface proteins released after cell lysis [
2]. The beneficial effects of postbiotics on restoring symbiotic microbiota, strengthening epithelial barrier function, and supporting the immune response have been reported [
3,
4].
Lactiplantibacillus plantarum, formerly known as
Lactobacillus plantarum, is a well-documented and extensively studied species of lactic acid bacteria. Its postbiotics consist of various beneficial metabolites, including organic acids, particularly lactic acid, which have a wide range of antimicrobial effects against various pathogens [
5,
6,
7].
The symbiotic microbial community in the oral microbiota can deteriorate and transform into dysbiotic communities [
8].
Prevotella is one of the most commonly identified genera in the oral microbiome [
9]. Comparative genomic analysis shows that there is a large gene repertoire of
Prevotella strains in the human oral cavity, allowing adaptation to different niches [
10,
11].
Prevotella strains are non-spore-forming, immotile rods, and most of them are saccharolytic and can use complex carbohydrates [
12].
Prevotella denticola,
Prevotella loescheii, and
Prevotella intermedia accelerate biofilm formation by enabling interspecies adhesion and providing a suitable environment for late colonizers to form complex biofilms [
13,
14].
F. nucleatum, a commonly isolated bacterium of the oral cavity, promotes coaggregation of early and late colonizers and generates an oxido-reduction potential low enough to allow survival of, for example,
Porphyromonas gingivalis, a potential periodontal pathogen [
15,
16].
Streptococcus sanguinis, another commensal member of the oral microbiota, is associated with the formation of healthy plaque but also favors the accumulation of some periodontal pathogens such as
P.
gingivalis and
F.
nucleatum [
17,
18].
The Inhibitory properties of postbiotics have been studied in periodontitis-associated pathogens. Byproducts of lactobacilli extracted from cell-free supernatants of culture media reduce the biofilm formation of
Aggregatibacter actinomycetemcomitans [
19]. Additionally, the cellular response of epithelial cells against
P.
gingivalis can be normalized in the presence of metabolic byproducts of the
Lactobacillus rhamnosus Lr-32 strain [
20]. However, it is important to remember that periodontitis-associated pathogens are mostly oxygen- and pH-sensitive and require commensal oral bacteria to grow, form biofilms, and cause destructive periodontal disease [
21]. Therefore, current studies focus only on the inhibitory effect of postbiotics on the virulence of periodontal pathogens. However, these studies do not address how postbiotics may affect commensal-opportunistic bacterial groups that have the ability to adapt to the environment. In our study, we hypothesized that oral bacteria with the ability to adapt to pH would resist the antimicrobial and antibiofilm effects of postbiotics. Considering the complexity of oral microbial ecology, in this study, we evaluated the responses to postbiotics of
P.
denticola, a health-associated organism with the ability to adapt to pH,
F.
nucleatum, a bridging organism in oral biofilms, and
S.
sanguinis, a Gram-positive representative in interspecies interactions. In this context, crude and neutralized forms of postbiotics derived from the strain
L.
plantarum EIR/IF-1 were tested on individual strains of
P.
denticola,
F.
nucleatum, and
S.
sanguinis and their multispecies consortia. Finally, the crude postbiotics were tested on hPDLF cells to determine their biocompatible potential and possible cytotoxic effects.
2. Materials and Methods
2.1. Bacterial Strains, Culture Media, and Growth Conditions
The previously isolated and identified L. plantarum EIR/IF-1 strain (former NCBI GenBank accession number: MW057714.1 and known as L. plantarum F10, current accession number: OP810909.1) was available at Pharmabiotic Technologies Research Laboratory, Department of Biology, Faculty of Science, Ankara University. P. denticola ATCC 33185 (type strain), P. denticola AHN 32366 (clinical strain), P. denticola AHN 32482 (clinical strain), F. nucleatum ATCC 25586 (type strain), and S. sanguinis NCTC 10904 (type strain) were obtained from the culture collections of the Institute of Dentistry, University of Turku, and used as test microorganisms in antimicrobial screening tests. Since it has not been adequately studied in the literature, clinical isolates of P. denticola were also included in the study for comparison with the reference strain.
Before postbiotics preparation, the L. plantarum EIR/IF-1 strain stored in 50% glycerol at −86 °C was first grown for 24 h at 37 °C under static conditions on De Man, Rogosa, and Sharpe Agar (MRS Agar, Merck, Darmstadt, Germany). Before antimicrobial screening, strains of P. denticola ATCC 33185, P. denticola AHN 32366, P. denticola AHN 32482, and F. nucleatum ATCC 25586 were grown on Brucella Blood Agar Medium supplemented with 750 mg/mL cysteine, 5 mg/mL hemin, and 10 mg/mL vitamin K1. The S. sanguinis NCTC 10904 strain was grown on Brain Heart Infusion Agar (BHI Agar, Merck, Darmstadt, Germany). All strains were incubated under anaerobic conditions (10% H2, 5% CO2, and 85% N2, Whitley A35 Anaerobic Workstation, Don Whitley Scientific Ltd., West Yorkshire, UK) at 37 °C for 4–5 days.
2.2. Postbiotics Preparation
A characteristic colony of the previously activated
L. plantarum EIR/IF-1 strain on MRS Agar was taken and suspended in 20 mL of MRS Broth (MRS Broth, Merck, Darmstadt, Germany). After overnight incubation (corresponding to the late log phase) at 37 °C, this bacterial culture (≈10
8 CFU/mL) was used to inoculate 1 L of MRS Broth at an inoculation ratio of 2% (
v/
v). After another overnight incubation at 37 °C, the culture broth was centrifuged at 15,000×
g for 20 min at room temperature, and the supernatant (spent culture medium) was obtained. The supernatant was sterilized using membrane filters (0.22 μm pore size, Sartorius, Göttingen, Germany) [
22]. The filtered samples were then freeze-dried and powdered (freezing conditions of −20 °C, a vacuum pressure of 0.120 mB, and a condenser temperature of −58 °C; Christ freeze dryer, Harz, Germany) Approximately 10 g of dried powder was obtained from 1-L culture supernatant. MRS Broth was also freeze-dried and powdered to use as a control for further experiments.
2.3. Minimum Inhibitory Concentrations (MIC) and Minimum Bactericidal Concentrations (MBC) Tests
MIC and MBC values for postbiotics were determined by the microdilution method according to Clinical and Laboratory Standards Institute (CLSI) guidelines [
23]. Briefly, Todd-Hewitt Broth media (casein peptone 10 g/L, heart infusion 3.1 g/L, sodium carbonate 2.5 g/L, dextrose 2 g/L, sodium chloride 2 g/L, disodium phosphate 0.4 g/L; supplemented with 750 mg/mL cysteine, 5 mg/mL hemin, and 10 mg/mL vitamin K1) contained various concentrations of crude postbiotics or neutralized (pH 7) postbiotics (0–50 mg/mL). The activated bacterial cultures were adjusted to 0.5 McFarland standard and transferred to the wells of the microtiter plates. The test groups contained postbiotics and inoculum. The wells containing postbiotics without inocula served as negative controls, whereas the wells containing positive controls contained only media and inocula. After 48 h of incubation at 37 °C under anaerobic conditions, cell density was measured at a wavelength of 490 nm in a microplate reader and the lowest concentration without growth (MIC values) was determined.
After incubation, 100 µL of the culture suspensions were taken from each well and diluted in phosphate-buffered saline (PBS, pH 7.4) to perform colony counting. Each dilution was spread on Brucella Blood Agar plates, and the plates were incubated at 37 °C under anaerobic conditions for at least 4 days. Colonies were counted, and log reduction compared to the control groups was calculated. Values corresponding to a log reduction of ≥99.9% were determined as MBC values.
2.4. Antimicrobial Screening Assay
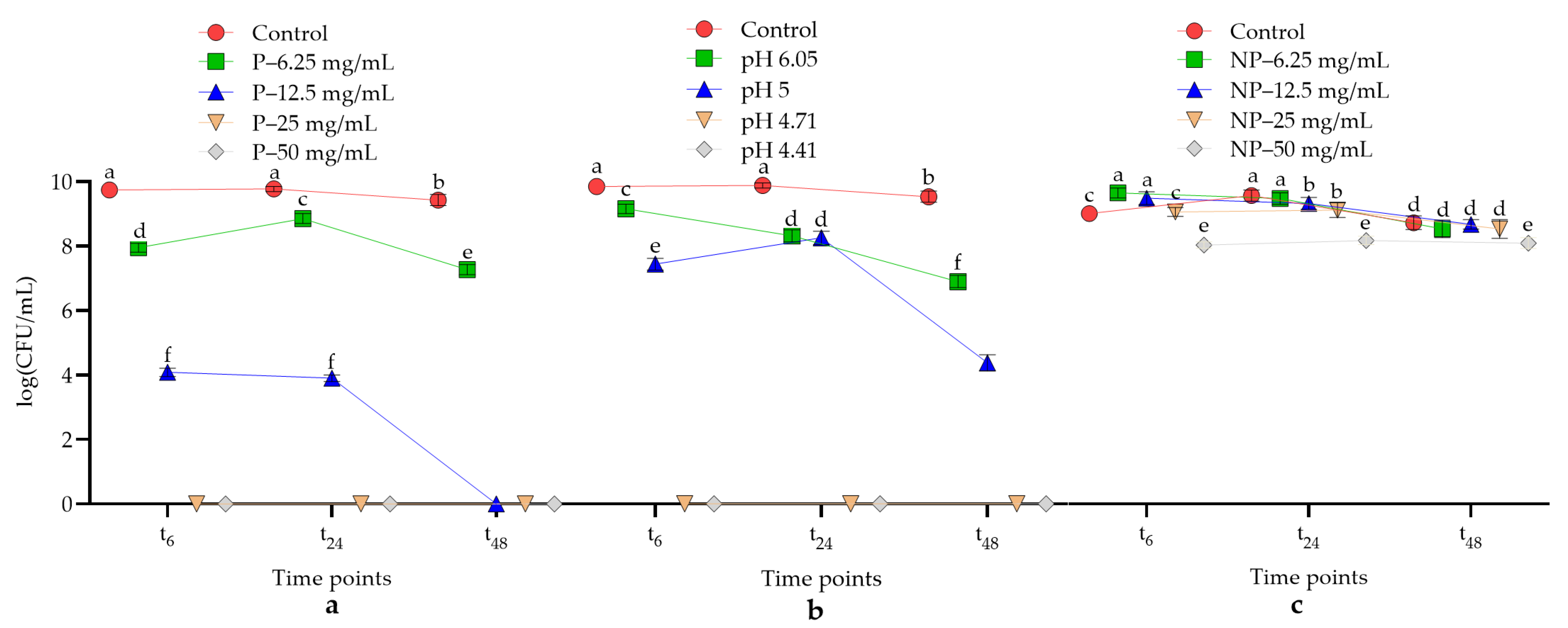
Culture media (Todd-Hewitt Broth) containing various concentrations (0, 6.25, 12.5, 25, and 50 mg/mL) of crude postbiotics were prepared, and the pH of the media was measured with a pH meter. The pH values of the media that contained crude postbiotics at different concentrations are shown in
Table 1. To test the acid-dependent antimicrobial activities of the postbiotics, pH-adjusted postbiotics-free media were used as positive controls. Neutralized postbiotic samples were also prepared with 2 M NaOH to test acid-independent antimicrobial activity.
Colonies from 4-day bacterial pure cultures were harvested and suspended in PBS (pH 7.4). The optical density (OD) of the suspensions was adjusted to 2.0 at 490 nm. Colony counting was performed to calculate the CFU/mL values of the suspensions, and each bacterial suspension was confirmed to be ≈109 CFU/mL. Twenty-five microliters of these suspensions were inoculated into 2 mL of Todd-Hewitt Broth media containing various concentrations of crude and neutralized postbiotics and acidified Todd-Hewitt Broth media. After inoculation, the culture suspensions were incubated for 48 h at 37 °C under anaerobic conditions. During incubation, the culture suspensions were shaken with the vortexer and at 6, 24, and 48 h time points, a 300 µL sample was taken from each culture suspension and transferred to microcentrifuge tubes. A sample of 100 µL was measured with a pH meter with the appropriate probe and a sample of 100 µL was used for colony counting. The last 100 µL sample was used for growth recovery. For this purpose, the collected samples were inoculated in 5 mL of fresh Todd-Hewitt Broth and the samples were incubated for another 4 days. In this way, it was checked whether the growth recovered at the concentrations where the antimicrobial activity was observed.
2.5. Antibiofilm Screening Assay
To determine the antibiofilm effects of crude postbiotics, polymicrobial biofilm models of
P.
denticola,
F.
nucleatum, and
S.
sanguinis were developed based on their interactions in oral biofilms [
24,
25].
Bacterial strains were cultured as stated in
Section 2.1. Suspensions of the cultures were prepared by adjusting each of them to OD: 2.0 at a wavelength of 490 nm (≈10
9 CFU/mL). An equal volume of each bacterial suspension was mixed to prepare different combinations as stated above. Pasteurized human saliva was used to coat the wells of the polystyrene plates as described previously [
26]. Next, 135 µL of Todd-Hewitt Broth adjusted with various concentrations of crude postbiotics was added to the saliva-coated wells (3.12, 6.25, 12.5, and 25 mg/mL postbiotics concentrations). Mixed bacterial suspensions (15 µL) were inoculated into the wells and then the plates were incubated for 48 h at 37 °C under anaerobic conditions. The wells containing only inocula served as positive control groups, while the wells containing only different concentrations of postbiotics without inoculum served as negative control groups. After incubation, the biofilm formation was determined using a crystal violet binding assay [
27].
2.6. Determination of the Cytotoxic Effects of Postbiotics on hPDLFCs
Previously isolated hPDLFCs were used in this study [
28]. Healthy premolar teeth of 20–25-year-old patients were harvested and washed in sterile PBS containing 5% penicillin-streptomycin.
Periodontal membranes were incised, minced, and digested in a protease solution (0.1% collagenase, type I, 0.25–1 mg/mL solid FALGPA (N-(3-[2-furyl] acryloyl)-LeuGly-Pro-Ala) for 45 min in a CO2 incubator (Panasonic, Osaka, Japan). The digested tissue was then centrifuged at 300× g for 5 min (Hettich, Westphalia, Germany). The pellet was transferred to DMEM (Dulbecco’s modified Eagle’s medium) containing 10% fetal bovine serum (FBS), 1% penicillin-streptomycin, 1% non-essential amino acid stock solution, and %1 L-glutamine (all from Biological Industries, USA) and incubated at 37 °C, 5% CO2. The morphology of hPDLFCs was examined using an inverted-phase contrast microscope (PrimoVert, Zeiss, Mainz, Germany).
The 96-well plates were seeded with hPDLFCs at a density of 1 × 104 cells/well. After 24 h of incubation, the media were removed, and the wells were rinsed with sterile PBS. Growth media adjusted with various concentrations of crude postbiotics ranging from 0–5000 µg/mL were added to the wells. Wells without crude postbiotics were used as positive controls. The plates were incubated for an additional 24 h. A cytotoxicity assay was performed using MTT 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] according to the manufacturer’s recommendation with the Cell Growth Determination Kit, MTT based, CGD1, Darmstadt, Germany, Merck).
2.7. Statistical Analysis
Each experiment was performed in triplicate, at least at two independent time points. Data are presented as mean values and standard deviations. Multiple comparisons were performed with the One-Way ANOVA test, and Tukey’s test was used for post-hoc comparisons. A p value of 0.05 was accepted as statistically significant. All analyses were carried out with GraphPad software (version 8.0, Boston, MA, USA).
4. Discussion
This study shows for the first time that postbiotics from the L. plantarum EIR/IF-1 strain have antimicrobial and antibiofilm activities against P. denticola, F. nucleatum, and S. sanguinis. It was also found that the acidity of the postbiotics from the L. plantarum EIR/IF-1 strain was mainly related to the observed antimicrobial and antibiofilm properties. Finally, our results suggest that the antibacterial activity of L. plantarum EIR/IF-1 postbiotics is not solely dependent on their acidity.
In this study,
P.
denticola,
F.
nucleatum, and
S.
sanguinis were preferred to evaluate the antimicrobial and antibiofilm activities of postbiotics because these bacteria are abundant in both healthy and disease-associated biofilms. A wide range of interspecies interactions have been observed in oral biofilms, with
Prevotella in particular interacting with a wide range of microorganisms [
29]. Prevotella is also involved in the formation of complex oral biofilms, and
P.
denticola occurs as a commensal in normal oral flora but can also be detected in dysbiotic biofilms [
14,
30]. Because it has not been adequately studied in the literature, clinical isolates of
P.
denticola were also included in the study for comparison with the reference strain.
S.
sanguinis also has an arsenal of surface proteins that can facilitate adhesion between different species [
31]. Another important component of oral biofilms is
F.
nucleatum due to its abundance and ability to aggregate with other bacterial species in the oral cavity [
32]. Consistent with the literature, the complex ecology of many microorganisms is thought to be at the root of the link between oral health and disease. In recent years, considerable attention has been paid to the use of postbiotics against oral microorganisms. However, the design of current studies is usually based on the evaluation of only one type of microorganism. In the current study, the strain
L.
plantarum EIR/IF-1 was preferred due to its antimicrobial activity against various pathogens. The strain EIR/IF-1 was rated as the best strain in the current collection [
33].
A limitation of this study concerns the partial chemical characterization of the postbiotics, as they are a cocktail of metabolites. In our previous studies, we determined the composition of organic acids of the strain
L.
plantarum EIR/IF-1 by high-performance liquid chromatography (HPLC) to investigate the antimicrobial activities of the postbiotics [
27]. Among the organic acids in the composition of
L.
plantarum EIR/IF-1 postbiotics, lactic acid was interpreted as the highest concentration (35.82 mg/mL), and the concentrations of formic and maleic acids were also relatively high (3.55 and 2.47 mg/mL, respectively). Malic acid, succinic acid, butyric acid, acetic acid, and tartaric acid were also detected at concentrations of 0.18, 0.20, 0.22, 0.98, and 1.62 mg/mL, respectively. However, if we consider the other metabolites of postbiotics, which have a complex chemical composition, organic acids make up only a small part of this complex composition. This complexity can make it difficult to evaluate potential benefits because the ingredients are not well documented. Most of the literature points to the health benefits of live bacteria, but scientific evidence for the use of postbiotics is growing. Postbiotics also offer advantages over the use of live organisms, including the possibility of chemical characterization and dose adjustment [
2]. Another limitation is that the slightly acidic environment (6.01–6.10) caused by the low concentration (6.25 mg/mL) of postbiotics may favor the growth of
P.
denticola strains. Therefore, the appropriate use of postbiotics containing organic acids should be tested on oral bacteria that can adapt to acidic conditions. Appropriate dosing strategies should also be tested. This is because, as in this study, inappropriate postbiotic concentrations can lead to unexpected results, especially with
P.
denticola strains. Although postbiotics can usually be used directly as a complex cocktail due to their various beneficial properties, it would be difficult to separately evaluate the potential antimicrobial and antibiofilm capabilities of the numerous and diverse individual metabolites in postbiotics. Despite this difficulty, this challenge can also be partially overcome by the fact that postbiotics can be used directly as a complex component in many applications. Another limitation would be that only cell-free supernatant was used as postbiotics in this study. This is because intracellular and cell wall/membrane components also fall under the definition of postbiotics. To some extent, cell-free culture supernatant is both easier to obtain and standardize than the other postbiotic components. In this study, significant antimicrobial and antibiofilm effects were observed by using cell-free culture supernatant as postbiotics source.
Cell-free culture supernatants, which fall under the definition of postbiotics, may have cytotoxic effects on eukaryotic cells because of the organic acids they contain. Although there are few studies on the effects of cell-free culture metabolites on periodontal cells, some of these studies have shown that postbiotics from lactic acid bacteria have toxic effects. For example, culture supernatant of the
L.
rhamnosus Lr-32 strain impairs the viability of gingival epithelial cells at increasing concentrations [
20]. Although there is no study in the literature on the possible cytotoxic effects of postbiotics on hPDLF cells, it is also known that metabolites such as acetate and propionate, which may be present in postbiotics, can have cytotoxic effects on various human cell lines [
34]. Although the antimicrobial and antibiofilm benefits of postbiotics should not be disregarded, the potential cytotoxic effects of these ingredients on healthy cells in the oral flora should be considered and dose-dependent approaches should be evaluated in the future use of these ingredients in periodontal treatment. In particular, organic acids that may be present in postbiotics and have a potentially toxic effect on oral cell lines should be evaluated separately.
The crude postbiotics were also tested against polymicrobial consortia. Finally, as an important contribution to the literature, bacterial recovery after the application of
L.
plantarum EIR/IF-1 postbiotics was performed to understand whether the antimicrobial effect is durable. Simply put, agents with bacteriostatic activity can prevent the growth of microorganisms by keeping them in the stationary phase, and agents with bactericidal activity can kill microorganisms [
35]. Essentially, the application of growth recovery was performed only to evaluate whether the efficacy of
L.
plantarum EIR/IF-1 postbiotics was bacteriostatic or bactericidal on the oral microorganisms tested. Again, we demonstrated that the antimicrobial activity of
L.
plantarum EIR/IF-1 postbiotics was bactericidal against the oral bacteria tested.
In the oral cavity, pH is an important regulator of bacterial presence, colonization, and biofilm formation [
36]. Fluctuations in pH affect the structure of proteins and phospholipids in the cell membrane, increase the permeability of the cell membrane, and may eventually lead to leakage of internal cell components. In addition to this basic effect, acids can cause cell death by inhibiting processes such as DNA replication and protein synthesis in the target cell [
37].
P.
intermedia can grow under acidic conditions in the range of 5.0–7.0 and
F.
nucleatum ATCC 25586 can grow at acidic pH values in the range of 5.5–7.0. To survive under acidic conditions, these bacteria deaminate amino acids and provide acid neutralization [
38]. According to our results, both
P.
denticola and
F.
nucleatum survive in a pH range of 5–7.4. Moreover, the strain
F.
nucleatum ATCC 25586 is more sensitive to acidic postbiotics than the strains of
P.
denticola, and the antimicrobial activity of postbiotics proved to be stronger against
F.
nucleatum ATCC 25586 than that of HCl-acidified media. Bosch et al. [
39] demonstrated that lactic acid-producing probiotic bacteria exhibited strong antimicrobial activity against
P.
denticola and
F.
nucleatum. Considering that the predominant organic acid in the postbiotics tested in this study is lactic acid, the antimicrobial effect observed in the present study and in the study by Bosch et al. [
39] could be due to lactic acid. When the pH of the environment is lowered to 4.04 in the presence of a carbohydrate source, non-mutans streptococci such as
S.
sanguinis can still survive within 60 min. The acid tolerance of
S.
sanguinis may be due to the activity of the enzyme arginine deiminase [
39]. In contrast, the strain
S.
sanguinis NCTC 10904 used in this study did not survive under the influence of acidic postbiotics, which lowered the pH to 4.40–6.0. This result suggests that the acid tolerance of
S.
sanguinis is less effective in long-term cultures.
Lactic acid is the dominant organic acid in
L.
plantarum EIR/IF-1 postbiotics. When lactic acid interacts with the bacterial cell membrane, it leads to acidification of the membrane and consequent failure of the proton drive. Lactic acid can act on many bacteria, including Gram-negative species. Lactic acid has a high affinity for water-filled porins on the outer membrane of Gram-negative bacteria. Acidic agents with high affinity such as lactic acid cause the release of LPS from the outer membrane. This increases membrane permeability and leads to cell death [
40]. The antimicrobial activity of acidic postbiotics on Gram-negative
F.
nucleatum and
P.
denticola can be explained by this general mode of action.
Postbiotics may contain not only bacteriocins but also various other antimicrobial substances that exhibit antimicrobial activity in addition to organic acids [
2]. To test this hypothesis, we used neutralized postbiotics and found that the neutralized postbiotics (pH 7) exhibited significant antimicrobial activity, especially in the case of
P.
denticola ATCC 33185. A short-term antibacterial effect with lower efficiency was also observed in other test strains. Interestingly, the growth of clinical strains of
P.
denticola increased with increasing concentration of neutralized postbiotics, which may be explained by the large abundance of postbiotics metabolites that these strains can use [
27]. Thus, our results suggest that the effect of
L.
plantarum EIR/IF-1 postbiotics on the growth of the tested oral bacteria is strain-dependent. Finally, the fact that postbiotics exhibit stronger antimicrobial activity than acidified media, especially in the case of
S.
sanguinis, may indicate antimicrobial components other than organic acids. The neutralized crude extract of postbiotics was also treated with proteinase K and catalase enzymes to determine whether a peptide-based antimicrobial agent or hydrogen peroxide-induced antimicrobial activity was present. While a decrease in antimicrobial activity was observed in postbiotic activity after treatment with proteinase K, no hydrogen peroxide-derived antimicrobial activity was observed after treatment with catalase [
33].
Postbiotics of
L.
rhamnosus and
Lactobacillus acidophilus reduce the biofilm formation and virulence of
A.
actinomycetemcomitans [
19]. Additionally, postbiotics of strain
L.
rhamnosus Lr-32 show immunomodulatory effects on gingival epithelial cells infected with
P.
gingivalis, more effectively than live probiotic cells [
20]. The spent culture medium of
Lactobacillus salivarius strain MG4265 was found to inhibit
Streptococcus mutans biofilm formation and tartrate-resistant acid phosphatase activity (TRAP) [
41]. Sub-MIC concentrations of postbiotics from the strains
L.
plantarum EIR/IF-1,
Lactiplantibacillus curvatus EIR/DG -1, and
L. curvatus EIR/BG-2 are effective in inhibiting the expression of
gtfC gene, which plays a role in the glucosyltransferase enzyme of
S.
mutans, and
comA and
comX genes, which play a role in regulating the two-component system [
27]. As indicated in the literature, postbiotics from lactic acid bacteria have antibiotic activity against oral pathogens. However, these studies were conducted on only one type of pathogenic microbial species. Therefore, we investigated the effects of postbiotics on biofilm formation of consortia of
F.
nucleatum,
P.
denticola, and
S.
sanguinis. In particular, the antibacterial effect of postbiotics was different among
P.
denticola strains. The postbiotics of
L.
plantarum EIR/IF-1 strains showed antibiofilm activity against the formation of polymicrobial biofilms even at sub-MIC concentrations in which the strain
P.
denticola AHN 32366 is present. Finally, considering the efficacy of the postbiotics of
L.
plantarum EIR/IF-1 in terms of antimicrobial and antibiofilm activity, the effective concentrations were 25 and 50 mg/mL, corresponding to the preferred oral bacterial strains and polymicrobial biofilm consortia under defined in vitro conditions. It seems possible that the antibiofilm activity of
L.
plantarum EIR/IF-1 postbiotics is independent of the antimicrobial activity, especially in polymicrobial consortia containing the strain
P.
denticola AHN 33266. This situation requires further studies to show how the related postbiotics exhibit antibiofilm activity against oral bacteria.
Our data suggest that postbiotics from the L. plantarum strain EIR/IF-1 can inhibit polymicrobial oral biofilm formation. However, because no data are available in the literature on the effects of postbiotics on polymicrobial biofilms of the species examined in this study, our results are of limited comparability. Additionally, because the method used to determine the amount of biofilm was an indirect method, it was difficult to determine which bacteria in the polymicrobial biofilm were affected by the use of postbiotics.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}