
Microorganisms and Climate Change: A Not So Invisible Effect


Abstract
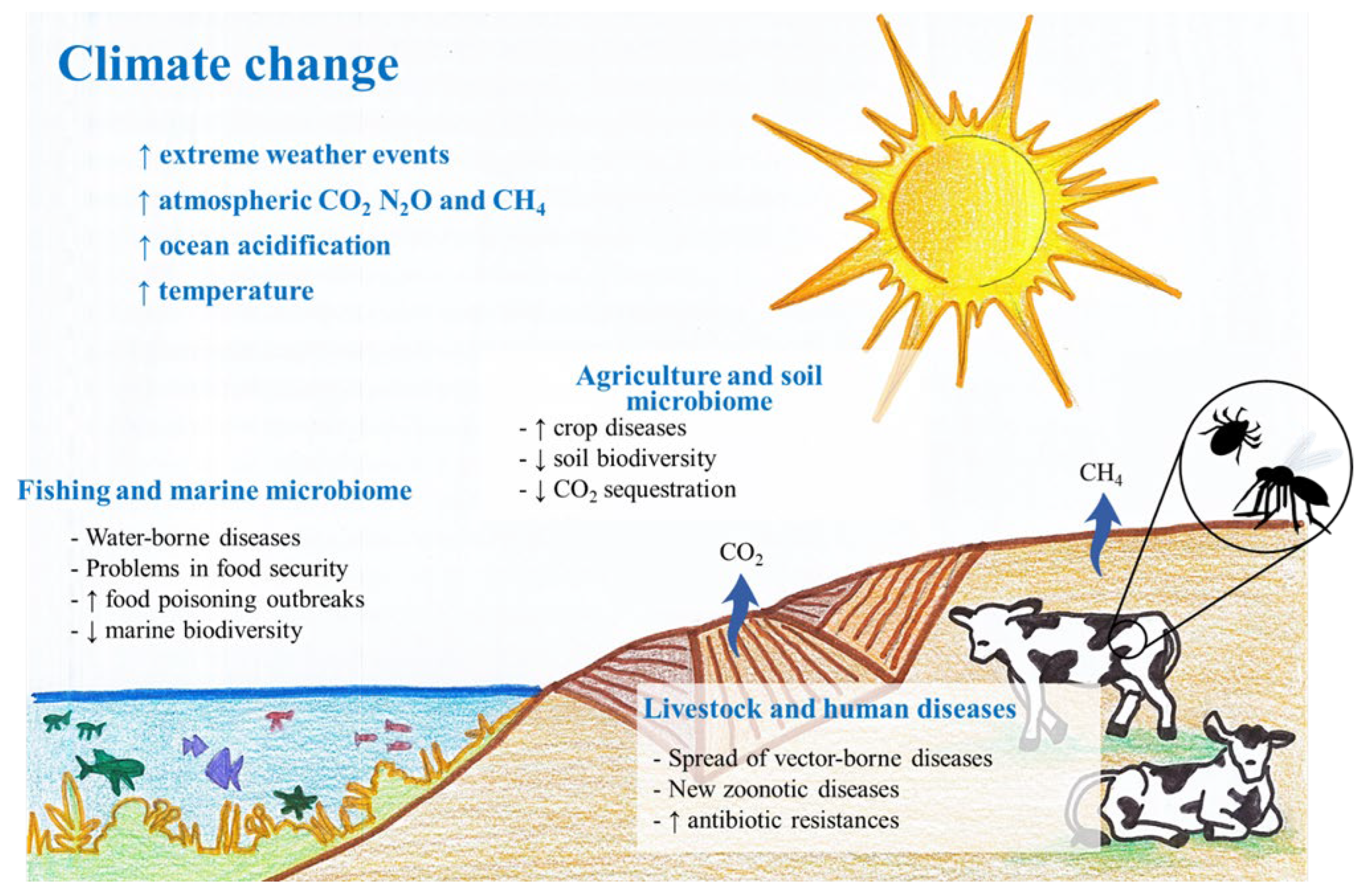
1. Introduction
2. Impact of Climate Change on Microbial Diversity
3. Microorganism Involvement in Production/Consumption of Greenhouse Gases
4. Microorganism-Mediated Effect on Productive Sectors
4.1. Agriculture and Soil Microbiome: Eternal Feedback
4.2. Plant Pests’ and Diseases’ Effect on Agriculture and Forest: The Uncomfortable Travelers
- Brown rust (mainly caused by Puccinia recondita), in the case of wheat, is forecast to increase its pressure on the crop by 20–100%, and yellow rust (caused by Puccinia striiformis) will increase by 5–20% in cold regions.
- Rice pathogens such as Pyricularia oryzae (the main cause of blast) and Bipolaris oryzae (or brown spot) are favored in all European rice districts, with the most critical situation in northern Italy (with an increase of close to 100%).
- In the case of grape, Plasmopara viticola (also called downy mildew) will increase by 5–20% across Europe, while Botrytis cinerea (or bunch rot) will have diverse impacts, ranging from a 20% decrease to a 100% increase in infection events [44].
- An increase in cases of diseases in leafy vegetable and cereal crops has already been reported in Italy, as a result of several pathogens’ effect, such as Plectosphaerella cucumerina, Alternaria sp., Fusarium equiseti, Myrothecium verrucaria, Myrothecium roridum, Phoma valerianellae, Pleospora betae, Peronospora belbahrii and Pythium ultimum, as well as the appearance of new pathogens like different species of Pythium (Pythium aphanidermatum, Pythium irregulare, Pythium dissotocum, Pythium coloratum, Pythium diclinum or Pythium lutarium) and new species causing yellow rust or stem rust [44,45,46].
- Among the new infections that have been reported in recent years in southern Europe and the Mediterranean coast, different species of the fungus Diaporthe have been found infecting several citruses, and the bacterium Xylella fastidiosa has also been discovered in olive trees [47].

{kind=link}
{kind=link}
{kind=link}
| Disease | Pathogen | Host | Origin | Spread and Development | Refs. |
|---|---|---|---|---|---|
| Ash dieback | Hymenoscyphus fraxineus | Ash trees | Asia | Asia, Europe and Africa | [49,50] |
| Bacterial blight or Bacterial leaf blight | Xanthomonas oryzae | Rice | Japan | Worldwide (especially Asia and Africa) | [51,52] |
| Bacterial canker | Pseudomonas syringae | Fruit trees | Depends on pathovar | Worldwide | [53,54] |
| Brown rust or Leaf rust | Puccinia recondita | Cereals (wheat, rye and barley) | Eastern Australia | Worldwide | [55,56] |
| Brown spot | Bipolaris oryzae | Rice | USA | Asia, Europe and South America | [44,57] |
| Bunch rot or Gray mold | Botrytis cinerea | Wide range | Unknown | Worldwide | [58] |
| Chestnut canker | Cryphonectria parasitic | Chestnut tree | Asia | North America | [59] |
| Disease dependent on the Diaporthe species | Diaporthe spp. | Wide range | Germany | Europe, Australia and Asia | [47,60] |
| Disease dependent on the X. fastidious subspecies | Xylella fastidiosa | Wide range | USA | South and North America and Europe | [47,61] |
| Downy mildew | Plasmopara viticola | Grape | North America | Worldwide | [62,63,64] |
| Dry root rot | Rhizoctonia bataticola (also Macrophomina phaseolina) | Chickpea | India | North America, Asia and Africa | [65] |
| Dutch Elm disease | Ceratocystis ulmi (also Ophistoma ulmi) | Elm | Asia | Worldwide | [66] |
| Fire blight | Erwinia amylovora | Apple, Pearl and some Rosaceae | North America | Europe and Asia | [67] |
| Rice blast | Magnaporthe oryzae (anamorph Pyricularia oryzae) | Rice | Brazil | South America, Asia, Africa and Europe | [44,68] |
| Stewart’s wilt | Pantoea stewartii (formerly Erwinia stewartii) | Corn | USA | Italy, Malaysia | [69,70] |
| Yellow rust or Stripe rust | Puccinia striiformis | Wheat | Transcaucasia (Armenia, Georgia and Azerbaijan) | Worldwide | [55,71] |
| Wheat blast | Pyricularia graminis-tritici | Wheat | Brazil | North and South America and Asia | [72] |
- Chestnut blight, caused by the pathogenic fungus Cryphonectria parasitica, native to Southeast Asia, which has killed more than 4 billion trees in the US to date.
- Ash dieback, caused by the newly identified Hymenoscyphus fraxineus, first detected in Asia, which has been recently detected in the UK and Northern Ireland [59].
- Dutch Elm disease, caused by Ceratocystis ulmi, a fungus responsible for its spread from Asia to Europe in 1918 that has killed millions of elms in Europe, western Asia and North America all throughout the 20th century [74].
- Needle blight of Pinus contora in north-western British Columbia, Canada, caused by outbreaks of Dothistroma septosporum as a result of a summer rainfall increase [75].
4.3. Livestock and Climate Change: An Arthropoda Matter
| Disease | Pathogen | Vector | Host | Origin | Spread and Development | Refs. |
|---|---|---|---|---|---|---|
| Anaplasmosis | Anaplasma phagocytophilum | Ixodes scapularis, Ixodes pacificus | Sheep and cattle | Scotland | Worldwide | [81,82] |
| Babesiosis | Babesia microti, Babesia venatorum and Babesia divergens | Ixodes ricinus, I. scapularis | Mammals | Romania | Europe and North America | [83] |
| Babesia bovis and Babesia bigemina | Rhipicephalus microplus | Mammals | Asia, Africa, South and Central America | Europe and North America | [84] | |
| Bluetongue Virus | Orbivirus | Culicoides imicola | Ruminants | South Africa | USA, Canada, Australia, South and Central Europe | [85] |
| Canine Babesiosis | Babesia spp. | Dermacentor reticulatus | Mammals (especially cattle) and birds | Romania | Worldwide | [26] |
| Colorado tick fever | Coltivirus | Dermacentor andersoni | Mammals | Western US | Europe and North America | [86,87] |
| Ehrlichiosis | Ehrlichia chaffeensis and Ehrlichia ewingi | Amblyomma americanum | Mammals | Canada | North America and Europe | [88,89] |
| Leptospirosis | Leptospira spp. | Environmental transmission | Mammals | Japan and Europe | Asia, Australia, America and South Europe | [90,91] |
| Lyme disease | Borrelia burgdorferi | Ixodes scapularis | Rodents | USA | North America and Eurasia | [92,93,94] |
| Powassan virus disease | Powassan virus | Several tick species | Mammals | Unknown | North America | [95] |
| Q fever | Coxiella burnetii | D. reticulatus | Ruminants (cattle, goat and sheep) | Australia | Europe and North America | [96] |
| Rocky Mountain spotted fever | Rickettsia rickettsii | D. andersoni | Mammals | South and Central America | North America | [97] |
| Rickettsia parkeri | A. maculatum | Central and North America |
- Ixodes scapularis and Ixodes pacificus, which transmit anaplasmosis disease, mainly caused by the bacterium Anaplasma phagocytophilum (formerly Erilichia phagocytophilum), which has also been reported in new regions worldwide [98]. In addition, they can also transmit the pathogen bacteria Borrelia miyamotoi, which has recently been reported in new areas of North America [81,82,99].
- Dermacentor reticulatus, which can transmit canine babesiosis, tularemia or Q fever [100,101]. Canine babesiosis is caused by the intracellular protozoan Babesia spp., and the number of reported cases has increased in northern countries, including Canada, Germany, Hungary, Switzerland and the Netherlands [26,78,95]. Q fever, meanwhile, is an important zoonotic disease caused by the bacterium Coxiella burnetiid. It was originally described in Australia in 1933, although it began to spread across North America and Europe only two decades ago. In Europe, some cases were initially reported in southern countries such as France, Spain and Germany, although in recent years, significant outbreaks have been reported even in countries further north like the Netherlands [96].
- Ixodes ricinus, which transmits encephalitis and Lyme disease (borreliosis).
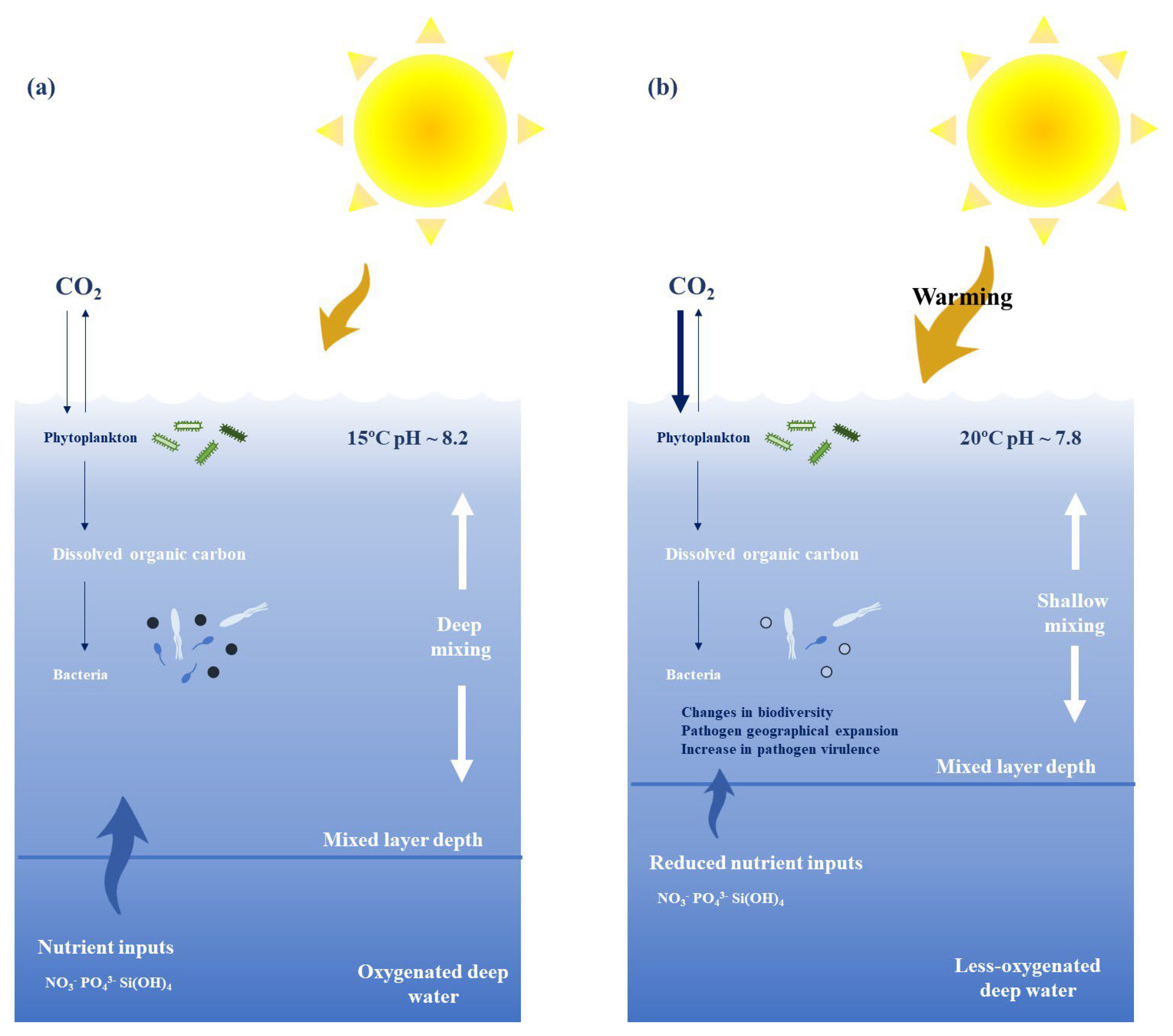
4.4. Fishing and Marine Microbiome
- First, environmental changes like those caused by climate change may lead to stress in both fish and shellfish species, leading to lower immune responses against various pathogens and diseases [2]. Urchins are a clear example of this. Both tank and real-world experiments showed a strong correlation between mass mortality events and long-term elevated temperatures. While urchins experience thermal stress, leading to a decrease in the immune response (increasing infection rates) and fertility (minimizing population recovery) at higher temperatures, pathogens increase their replication and transmission rates [122].
- Second, a rise in temperatures leads to an increase in several marine pathogens’ virulence by increasing their metabolism and inducing higher rates of transmission [122]. Such as the case of the host–pathogen interaction between Pocillopora damicornis and Vibrio coralliilyticus. At temperatures above 27 °C, pathogen virulence increases because multiple virulence factors are upregulated, including extracellular proteases that cause lysis and mortality in P. damicornis [123].
- Third, pathogen geographical expansion comes with increasing temperatures. A shining example is Perkinsus marinus, a protist parasite of the eastern oyster Crassostrea virginica that has expanded its range from the mid-Atlantic to the northeast in recent decades, primarily due to increased winter water temperatures, since both P. marinus infection patterns and C. virginica immune response are dependent on the temperature [124,125]. In addition, P. marinus infection intensity increases above 20 °C, especially at temperatures close to 25 °C, and some models predict longer periods of sustained higher temperatures after 2100, which may allow the geographical spread of P. marinus and its establishment farther north along the coast [122]. Other cases come from the Vibrio genus, which includes more than 110 different species. Some of them are well-known animal and human pathogens, like Vibrio cholerae, responsible for cholera disease, which causes between 100,000 and 120,000 deaths every year globally. Zooplankton is one of the main environmental reservoirs of vibrios in aquatic environments, and some species can be found infecting molluscan and crustacean shellfishes, as in the case of Vibrio parahaemolyticus and Vibrio vulnificus. Both species are related to typical human infections associated with seafood consumption, and the risk area for both vibrio infections has greatly increased during warmer water temperature episodes in the last years [113,126]. A similar effect is expected in other aquaculture species due to Vibrio harveyi, which poses a great risk to some of the most important species in the fish market, such as rainbow trout (Oncorhynchus mykiss), Atlantic salmon (Salmo salar L.), Senegalese sole (Solea senegalensis), Japanese seabass (Lateolabrax japonicus), cobia (Rachycentron canadum) and common dentex (Dentex dentex), among others [127].
- Last, habitat expansion is likely to cause novel contact among populations, pathogens and vectors, potentially increasing interspecies infections. For example, Brucella, a group of Gram-negative bacteria and the causative agents of brucellosis. Novel species of Brucella named Brucella ceti and Brucella pinnipedialis have been reported to cause infection in marine mammals like cetaceans and seals, respectively [122].
5. Human Health and Climate Change
5.1. Human Infective Diseases: Evolution and Expansion
5.1.1. Human Vector-Borne Diseases
| Disease | Microorganism | Vector | Host | Origin | Spread and Development | Effects of Climate Change | Refs. | |
|---|---|---|---|---|---|---|---|---|
| Vector-borne diseases | Chikungunya | Chikungunya virus | Aedes albopictus | Human | Tanzania | Asia and Africa | Increase in temperatures, changes in rainfall and increase in the number and severity of extreme weather events multiply the risk and the ranges of infection and the spread of diseases | [139] |
| Dengue | Dengue virus (DENV) | Aedes aegypti | Human | Africa | Tropical and subtropical areas and Europe | [133,142] | ||
| Aedes albopictus | ||||||||
| Leishmaniasis | Leishmania spp. | Female sand-fly | Human | Africa | Southern countries of the European continent | [138,139] | ||
| Zika | Zika virus (ZIKV) | Aedes albopictus | Human | Africa | North America and South America | [138,139] | ||
| Foodborne diseases | Diarrheal disease | Campylobacter spp. | Contaminated food | Human | America | Worldwide | Temperatures rise favors the contamination of food by pathogens that cause diseases | [138,139] |
| Salmonellosis | Salmonella spp. | Contaminated food | Human | America | Worldwide | [139,143] | ||
| Waterborne diseases | Cholera | Vibrio cholerae | Contaminated water | Human | Asia and Africa | Africa, Asia and North America | Increased precipitation can wash pathogens into waterways, while rising sea temperatures activate impulses for pathogen spread and development | [1,111,112,117,119,120] |
| Leptospirosis | Leptospira spp. | Contaminated water | Human | Germany | Tropical and subtropical areas | Warmer temperatures and extreme weather events can create favorable conditions for the survival and persistence of Leptospira bacteria in the environment. Alterations in land use and deforestation can also increase human contact with animal reservoirs of leptospirosis | [2] |
5.1.2. Foodborne Diseases
5.1.3. Waterborne Diseases
5.2. Antibiotic Resistance
6. Microbial Mitigation of Climate Change
- Inoculants: one of the main strategies to deal with the inevitable effects of climate change on crops is the use of biostimulants, which recover plant resistance and resilience to biotic and abiotic stresses showing low toxicity and avoiding the appearance of new resistant strains of pests and pathogens. Hence, the biofertilizer market has steadily increased in the last years, with Europe being the world industry leader [174]. Several microorganisms can act as biostimulants providing crop-limiting nutrients: (i) by nitrogen fixation (Azospirillum, Azotobacter or Rhizobium), (ii) mineral solubilization like phosphorus, potassium or zinc (Pseudomonas, Bacillus or arbuscular mycorrhiza) or (iii) siderophore production (Pseudomonas and Acinetobacter) [174,175].
- Biofertilizers tackle climate change concerns in different positions:
- (i)
- Some microbial species have shown crop protection from warmer temperatures, such as Paraburkholderia phytofirmans in potato crops, Bacillus and Azospirillum species in wheat and soy or Pseudomonas species in wheat [174],
- (ii)
- (iii)
- Methanotrophic bacteria are gaining attention as biofertilizers, since it has been estimated that they are able to consume approximately 40–60% of the methane produced in wetland environments. Thus, methanotrophs can be used in landfills and agricultural soils, ultimately helping to reduce atmospheric methane levels (e.g., Methylococcus or Methylococcus species) [22,178,179].
- Microorganisms are leaders in carbon sequestration:
- (i)
- Several soil microbes contribute to carbon sequestration through different mechanisms, although certain microbes possess faster metabolic rates and, therefore, sequester carbon faster. It is possible to enrich soils with these species of interest through the introduction of microbial formulations respecting the environment, as well as by enhancing the capacity to collect carbon in agricultural soil [180].
- (ii)
- In addition, studies on the production of biofuels at an industrial level by means of algae, which are one of the most powerful microorganisms for carbon fixation (1.83 kg of CO2 is needed for each kg of dry algae biomass production), connect two relevant concepts of the circular bioeconomy. On the one hand, CO2 sequestration is closely connected to the carbon credits that pollute the air, and on the other hand, the biomass produced by some of these algae genera (e.g., Botryococcus sp., Scenedesmus, Neochloris) is ideal for energy fuel production and multiple value-added products (e.g., feeds). Therefore, it would be a particularly interesting cycle, since concerns about the depletion of fossil fuels have increased general interest in recent years, and this process could be interesting not only from an ecological point of view for the biological sequestration of carbon from punctual sources but also from an economic one [181].
- Livestock rumen: manipulation of rumen microbiota and breeding programs has been proposed as a suitable solution to reduce methane emissions from cattle. The objective would be to obtain cattle lines producing less methane without affecting the health and productivity of animals [2].
- The Rs concept is aimed at the transition to a circular bioeconomy by means of different wording that collects the spirit of the materials’ reinsertion into the production system. Initially, three Rs were considered (Reduce, Reuse and Recycle); later, it was increased to six Rs (the three Rs plus Rethink, Refuse, Repair), and, nowadays, it continues increasing the number of Rs (the six Rs plus Refurbish, Remanufacture, Repurpose, Recover). In this context, the recent isolation of the poly(ethylene terephthalate) (PET)-degrading bacterium Ideonella sakaiensis [182], the description of epoxy-degrader microorganisms, such as Pseudomonas putida [183], or the description of the wax moth caterpillars (Galleria mellonella) as polyethylene degraders [184] have boosted the research on the biotechnological degradation of plastic. Excessive use of these materials increases plastic accumulation on land and sea. Microbes are the predominant organisms able to face this problem, the so-called plastic biodegradation [185]. The degradation to these basic molecular components presents the microorganisms or their enzymatic activities as relevant players in environmental sustainability.
- New protein sources: the search for new sources of protein to cover the increasing demand for meat protein has led to single-cell protein (SCP) development. In the first place, efforts have been made to convert plant protein into meat protein, getting low yields. Thus, the production of SCP was initiated, which are dried microbial biomass or the total amount of protein extracted from bacteria, yeasts, fungi or algae cultures [186]. A clear example is the production of burger patties from the fungal protein of Aspergillus oryzae, with similar organoleptic properties to meat burgers and rich nutritional content [187].
- Bioremediation: oil spills are one example of pollution that can have devastating effects on ecosystems and habitats. The bacterium Alcanivorax borkumensis thrives in hydrocarbon-rich environments and has been shown to play a crucial role in the degradation of oil spills [188]. This microorganism has been used in several bioremediation efforts, including the clean-up of the Deepwater Horizon oil spill in the Gulf of Mexico in 2010 [189].
- Liquid-3: Pollution is growing steadily around major urban centers. In 2021, Dr. Ivan Spasojević from the University of Belgrade launched Liquid-3 on the market, a 600-L tank filled with microalgae design to remove CO2. The tank effect is able to replace two 10-year-old trees or 200 square meters of lawn (https://liquid3.rs/, accessed on 19 July 2023).
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sobel, A.H.; Tippett, M.K. Extreme Events: Trends and Risk Assessment Methodologies. In Resilience: The Science of Adaptation to Climate Change; Zommers, Z., Alverson, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 3–12. ISBN 9780128118917. [Google Scholar]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Buis, A. Flip Flop: Why Variations in Earth’s Magnetic Field Aren’t Causing Today’s Climate Change. Available online: https://climate.nasa.gov/explore/ask-nasa-climate/3104/flip-flop-why-variations-in-earths-magnetic-field-arent-causing-todays-climate-change/#:~:text=AirIsn’tFerrous,%3Aairisn’tferrous (accessed on 19 July 2023).
- Mikhaylov, A.; Moiseev, N.; Aleshin, K.; Burkhardt, T. Global climate change and greenhouse effect. Entrep. Sustain. Issues 2020, 7, 2897–2913. [Google Scholar] [CrossRef] [PubMed]
- Fourier, J. Remarques générales sur les températures du globe terrestre et des espaces planétaires. Ann. Chim. Phys. 1824, 27, 136–167. [Google Scholar]
- Broecker, W.S. Climatic Change: Are We on the Brink of a Pronounced Global Warming? Science 1975, 189, 460–463. [Google Scholar] [CrossRef]
- Knoll, A.H. Paleobiological Perspectives on Early Microbial Evolution. Cold Spring Harb. Perspect. Biol. 2015, 7, a018093. [Google Scholar] [CrossRef] [PubMed]
- Khursheed, S. Soil Biodiversity and Climate Change. Adv. Plants Agric. Res. 2016, 3, 166–168. [Google Scholar] [CrossRef][Green Version]
- Griffith, G.W. Do we need a global strategy for microbial conservation? Trends Ecol. Evol. 2012, 27, 1–2. [Google Scholar] [CrossRef]
- Banerjee, A.; Cornejo, J.; Bandopadhyay, R. Emergent climate change impact throughout the world: Call for “Microbiome Conservation” before it’s too late. Biodivers. Conserv. 2020, 29, 345–348. [Google Scholar] [CrossRef]
- Jain, P.K.; Purkayastha, S.D.; De Mandal, S.; Passari, A.K.; Govindarajan, R.K. Effect of climate change on microbial diversity and its functional attributes. In Recent Advancements in Microbial Diversity; Elsevier: Amsterdam, The Netherlands, 2020; pp. 315–351. [Google Scholar]
- Guo, X.; Zhou, X.; Hale, L.; Yuan, M.; Ning, D.; Feng, J.; Shi, Z.; Li, Z.; Feng, B.; Gao, Q.; et al. Climate warming accelerates temporal scaling of grassland soil microbial biodiversity. Nat. Ecol. Evol. 2019, 3, 612–619. [Google Scholar] [CrossRef]
- Guo, X.; Feng, J.; Shi, Z.; Zhou, X.; Yuan, M.; Tao, X.; Hale, L.; Yuan, T.; Wang, J.; Qin, Y.; et al. Climate warming leads to divergent succession of grassland microbial communities. Nat. Clim. Change 2018, 8, 813–818. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Shen, L.; Wen, C.; Yan, Q.; Ning, D.; Qin, Y.; Xue, K.; Wu, L.; He, Z.; et al. Temperature mediates continental-scale diversity of microbes in forest soils. Nat. Commun. 2016, 7, 12083. [Google Scholar] [CrossRef]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Xiao, N.; Ning, D.; Shi, Z.; Zhou, X.; Wu, L.; Yang, Y.; et al. Climate warming enhances microbial network complexity and stability. Nat. Clim. Change 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Singh, V.K.; Shukla, A.K.; Singh, A.K. Impact of Climate Change on Plant–Microbe Interactions under Agroecosystems. In Climate Change and Agricultural Ecosystems; Elsevier: Amsterdam, The Netherlands, 2019; pp. 153–179. [Google Scholar]
- Sharma, B.; Singh, B.N.; Dwivedi, P.; Rajawat, M.V.S. Interference of Climate Change on Plant-Microbe Interaction: Present and Future Prospects. Front. Agron. 2022, 3, 113. [Google Scholar] [CrossRef]
- Zhang, C.; Xi, N. Precipitation Changes Regulate Plant and Soil Microbial Biomass Via Plasticity in Plant Biomass Allocation in Grasslands: A Meta-Analysis. Front. Plant Sci. 2021, 12, 614968. [Google Scholar] [CrossRef]
- Vicuña, R.; González, B. The microbial world in a changing environment. Rev. Chil. Hist. Nat. 2021, 94, 2. [Google Scholar] [CrossRef]
- Fleming, A. On the antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzae. Br. J. Exp. Pathol. 1929, 10, 226–236. [Google Scholar] [CrossRef]
- Tiedje, J.M.; Bruns, M.A.; Casadevall, A.; Criddle, C.S.; Eloe-Fadrosh, E.; Karl, D.M.; Nguyen, N.K.; Zhou, J. Microbes and Climate Change: A Research Prospectus for the Future. mBio 2022, 13, e00800-22. [Google Scholar] [CrossRef]
- Masood, F.; Ahmad, S.; Malik, A. Role of Methanotrophs in Mitigating Global Warming. In Microbiomes and the Global Climate Change; Springer: Singapore, 2021; pp. 43–60. [Google Scholar]
- Mangodo, C.; Adeyemi, T.O.A.; Bakpolor, V.R.; Adegboyega, D.A. Impact of microorganisms on climate change: A review. World News Nat. Sci. 2020, 31, 36–47. [Google Scholar]
- FAO. Statistical Yearbook 2021: World Food and Agriculture; FAO (Food and Agriculture Organization): Rome, Italy, 2021. [Google Scholar]
- Mangalassery, S.; Sjögersten, S.; Sparkes, D.L.; Sturrock, C.J.; Craigon, J.; Mooney, S.J. To what extent can zero tillage lead to a reduction in greenhouse gas emissions from temperate soils? Sci. Rep. 2015, 4, 4586. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Sainz, Á.; Roura, X.; Estrada-Peña, A.; Miró, G. A review of canine babesiosis: The European perspective. Parasites Vectors 2016, 9, 336. [Google Scholar] [CrossRef]
- Kannojia, P.; Sharma, P.K.; Sharma, K. Climate Change and Soil Dynamics: Effects on Soil Microbes and Fertility of Soil. In Climate Change and Agricultural Ecosystems; Elsevier: Amsterdam, The Netherlands, 2019; pp. 43–64. [Google Scholar]
- Mekala, S.; Polepongu, S. Impact of Climate Change on Soil Microbial Community. In Plant Biotic Interactions; Springer International Publishing: Cham, Switzerland, 2019; pp. 31–41. ISBN 978-3-030-26657-8. [Google Scholar]
- Pugnaire, F.I.; Morillo, J.A.; Peñuelas, J.; Reich, P.B.; Bardgett, R.D.; Gaxiola, A.; Wardle, D.A.; van der Putten, W.H. Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Sci. Adv. 2019, 5, eaaz1834. [Google Scholar] [CrossRef] [PubMed]
- Sheik, C.S.; Beasley, W.H.; Elshahed, M.S.; Zhou, X.; Luo, Y.; Krumholz, L.R. Effect of warming and drought on grassland microbial communities. ISME J. 2011, 5, 1692–1700. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Classen, A.T.; Sundqvist, M.K.; Henning, J.A.; Newman, G.S.; Moore, J.A.M.; Cregger, M.A.; Moorhead, L.C.; Patterson, C.M. Direct and indirect effects of climate change on soil microbial and soil microbial-plant interactions: What lies ahead? Ecosphere 2015, 6, art130. [Google Scholar] [CrossRef]
- Lobell, D.B.; Asseng, S. Comparing estimates of climate change impacts from process-based and statistical crop models. Environ. Res. Lett. 2017, 12, 015001. [Google Scholar] [CrossRef]
- Liu, B.; Asseng, S.; Müller, C.; Ewert, F.; Elliott, J.; Lobell, D.B.; Martre, P.; Ruane, A.C.; Wallach, D.; Jones, J.W.; et al. Similar estimates of temperature impacts on global wheat yield by three independent methods. Nat. Clim. Change 2016, 6, 1130–1136. [Google Scholar] [CrossRef]
- Zampieri, M.; Ceglar, A.; Dentener, F.; Toreti, A. Wheat yield loss attributable to heat waves, drought and water excess at the global, national and subnational scales. Environ. Res. Lett. 2017, 12, 064008. [Google Scholar] [CrossRef]
- Matiu, M.; Ankerst, D.P.; Menzel, A. Interactions between temperature and drought in global and regional crop yield variability during 1961–2014. PLoS ONE 2017, 12, e0178339. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, J.; Chen, M. Economic impacts of climate change on agriculture: The importance of additional climatic variables other than temperature and precipitation. J. Environ. Econ. Manag. 2017, 83, 8–31. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The Still Underestimated Problem of Fungal Diseases Worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.-T.; Eisenhauer, N.; Singh, B.K.; Maestre, F.T. The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Clim. Change 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Change 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Miedaner, T.; Juroszek, P. Climate change will influence disease resistance breeding in wheat in Northwestern Europe. Theor. Appl. Genet. 2021, 134, 1771–1785. [Google Scholar] [CrossRef]
- FAO. Plant Pests and Diseases in the Context of Climate Change and Climate Variability: Food Security and Biodiversity Risks; FAO (Food and Agriculture Organization): Budapest, Hungary, 2019; Volume 41. [Google Scholar]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Plant pathogen infection risk tracks global crop yields under climate change. Nat. Clim. Change 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Bregaglio, S.; Donatelli, M.; Confalonieri, R. Fungal infections of rice, wheat, and grape in Europe in 2030–2050. Agron. Sustain. Dev. 2013, 33, 767–776. [Google Scholar] [CrossRef]
- Bhattacharya, S. Deadly new wheat disease threatens Europe’s crops. Nature 2017, 542, 145–146. [Google Scholar] [CrossRef]
- Gilardi, G.; Gullino, M.L.; Garibaldi, A. Emerging foliar and soil-borne pathogens of leafy vegetable crops: A possible threat to Europe. EPPO Bull. 2018, 48, 116–127. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Childers, A.K.; Chen, Y.; Madella, S.; Nessa, A.; VanEngelsdorp, D.; Evans, J.D. Recent spread of Varroa destructor virus-1, a honey bee pathogen, in the United States. Sci. Rep. 2017, 7, 17447. [Google Scholar] [CrossRef]
- Wallen-Russell, C.; Wallen-Russell, S. Topical Probiotics Do Not Satisfy New Criteria for Effective Use Due to Insufficient Skin Microbiome Knowledge. Cosmetics 2021, 8, 90. [Google Scholar] [CrossRef]
- Europe’s Changing Woods and Forests: From Wildwood to Managed Landscapes; Kirby, K.J., Watkins, C., Eds.; CABI: Wallingford, UK, 2015; ISBN 9781780643373. [Google Scholar]
- Yasmin, S.; Hafeez, F.Y.; Mirza, M.S.; Rasul, M.; Arshad, H.M.I.; Zubair, M.; Iqbal, M. Biocontrol of Bacterial Leaf Blight of Rice and Profiling of Secondary Metabolites Produced by Rhizospheric Pseudomonas aeruginosa BRp3. Front. Microbiol. 2017, 8, 1895. [Google Scholar] [CrossRef]
- Banerjee, A.; Roy, S.; Bag, M.K.; Bhagat, S.; Kar, M.K.; Mandal, N.P.; Mukherjee, A.K.; Maiti, D. A survey of bacterial blight (Xanthomonas oryzae pv. oryzae) resistance in rice germplasm from eastern and northeastern India using molecular markers. Crop Prot. 2018, 112, 168–176. [Google Scholar] [CrossRef]
- Wang, R.; Li, Q.; He, S.; Liu, Y.; Wang, M.; Jiang, G. Modeling and mapping the current and future distribution of Pseudomonas syringae pv. actinidiae under climate change in China. PLoS ONE 2018, 13, e0192153. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Łochyńska, M. Identification and characterization of Pseudomonas syringae pv. mori affecting white mulberry (Morus alba) in Poland. Eur. J. Plant Pathol. 2020, 158, 281–291. [Google Scholar] [CrossRef]
- Park, R.F.; Burdon, J.J.; McIntosh, R.A. Studies on the origin, spread, and evolution of an important group of Puccinia recondita f. sp. tritici pathotypes in Australasia. Eur. J. Plant Pathol. 1995, 101, 613–622. [Google Scholar] [CrossRef]
- Peksa, K.; Bankina, B. Characterization of Puccinia recondita, the causal agent of brown rust: A review. Res. Rural Dev. 2019, 2, 70–76. [Google Scholar]
- Schwanck, A.A.; Meneses, P.R.; Farias, C.R.J.; Funck, G.R.D.; Maia, A.H.N.; Del Ponte, E.M. Bipolaris oryzae seed borne inoculum and brown spot epidemics in the subtropical lowland rice-growing region of Brazil. Eur. J. Plant Pathol. 2015, 142, 875–885. [Google Scholar] [CrossRef][Green Version]
- Rosslenbroich, H.-J.; Stuebler, D. Botrytis cinerea—History of chemical control and novel fungicides for its management. Crop Prot. 2000, 19, 557–561. [Google Scholar] [CrossRef]
- FAO. The Impact of Disasters and Crises on Agriculture and Food Security; FAO (Food and Agriculture Organization): Rome, Italy, 2021. [Google Scholar]
- Thompson, S.M.; Tan, Y.P.; Shivas, R.G.; Neate, S.M.; Morin, L.; Bissett, A.; Aitken, E.A.B. Green and brown bridges between weeds and crops reveal novel Diaporthe species in Australia. Persoonia Mol. Phylogeny Evol. Fungi 2015, 35, 39–49. [Google Scholar] [CrossRef]
- Rapicavoli, J.; Ingel, B.; Blanco-Ulate, B.; Cantu, D.; Roper, C. Xylella fastidiosa: An examination of a re-emerging plant pathogen. Mol. Plant Pathol. 2018, 19, 786–800. [Google Scholar] [CrossRef]
- Kim, B.; Lee, J.S.; Choi, Y.-J. Plasmopara viticola Causing Downy Mildew on Vitis davidii in Korea. Plant Dis. 2021, 105, 3309. [Google Scholar] [CrossRef]
- Williams, M.G.; Magarey, P.A.; Sivasithamparam, K. Effect of temperature and light intensity on early infection behaviour of a Western Australian isolate of Plasmopara viticola, the downy mildew pathogen of grapevine. Australas. Plant Pathol. 2007, 36, 325. [Google Scholar] [CrossRef]
- Thines, M. Recent outbreaks of downy mildew on grape ivy (Parthenocissus tricuspidata, Vitaceae) in Germany are caused by a new species of Plasmopara. Mycol. Prog. 2011, 10, 415–422. [Google Scholar] [CrossRef]
- Sharma, M.; Ghosh, R.; Pande, S. Dry root rot (Rhizoctonia bataticola (Taub.) Butler): An emerging disease of chickpea—Where do we stand? Arch. Phytopathol. Plant Prot. 2015, 48, 797–812. [Google Scholar] [CrossRef]
- Hantula, J.; Vainio, E.J. Viruses as components of forest microbiome. In Forest Microbiology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 371–382. [Google Scholar]
- Gaganidze, D.; Sadunishvili, T.; Aznarashvili, M.; Abashidze, E.; Gurielidze, M.; Carnal, S.; Rezzonico, F.; Zubadalashvili, M. Fire blight distribution in Georgia and characterization of selected Erwinia amylovora isolates. J. Plant Pathol. 2021, 103, 121–129. [Google Scholar] [CrossRef]
- Singh, P.K.; Gahtyari, N.C.; Roy, C.; Roy, K.K.; He, X.; Tembo, B.; Xu, K.; Juliana, P.; Sonder, K.; Kabir, M.R.; et al. Wheat Blast: A Disease Spreading by Intercontinental Jumps and Its Management Strategies. Front. Plant Sci. 2021, 12, 1467. [Google Scholar] [CrossRef]
- Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; et al. Risk assessment of the entry of Pantoea stewartii subsp. stewartii on maize seed imported by the EU from the USA. EFSA J. 2019, 17, e05851. [Google Scholar] [CrossRef]
- Abidin, N.; Ismail, S.I.; Vadamalai, G.; Yusof, M.T.; Hakiman, M.; Karam, D.S.; Ismail-Suhaimy, N.W.; Ibrahim, R.; Zulperi, D. Genetic diversity of Pantoea stewartii subspecies stewartii causing jackfruit-bronzing disease in Malaysia. PLoS ONE 2020, 15, e0234350. [Google Scholar] [CrossRef]
- Ali, S.; Gladieux, P.; Leconte, M.; Gautier, A.; Justesen, A.F.; Hovmøller, M.S.; Enjalbert, J.; de Vallavieille-Pope, C. Origin, Migration Routes and Worldwide Population Genetic Structure of the Wheat Yellow Rust Pathogen Puccinia striiformis f.sp. tritici. PLoS Pathog. 2014, 10, e1003903. [Google Scholar] [CrossRef]
- McDonald, B.A.; Stukenbrock, E.H. Rapid emergence of pathogens in agro-ecosystems: Global threats to agricultural sustainability and food security. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160026. [Google Scholar] [CrossRef]
- DeAngelis, K.M.; Pold, G.; Topçuoğlu, B.D.; van Diepen, L.T.A.; Varney, R.M.; Blanchard, J.L.; Melillo, J.; Frey, S.D. Long-term forest soil warming alters microbial communities in temperate forest soils. Front. Microbiol. 2015, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Karnosky, D.F. Dutch Elm Disease: A Review of the History, Environmental Implications, Control, and Research Needs. Environ. Conserv. 1979, 6, 311–322. [Google Scholar] [CrossRef]
- Sukumar Chakraborty, S.C. Impacts of global change on diseases of agricultural crops and forest trees. CABI Rev. 2008, 3, 1–15. [Google Scholar] [CrossRef]
- Ali, M.Z.; Carlile, G.; Giasuddin, M. Impact of global climate change on livestock health: Bangladesh perspective. Open Vet. J. 2020, 10, 178–188. [Google Scholar] [CrossRef]
- FAO. Meat Market Review; FAO (Food and Agriculture Organization): Rome, Italy, 2019. [Google Scholar]
- Bett, B.; Kiunga, P.; Gachohi, J.; Sindato, C.; Mbotha, D.; Robinson, T.; Lindahl, J.; Grace, D. Effects of climate change on the occurrence and distribution of livestock diseases. Prev. Vet. Med. 2017, 137, 119–129. [Google Scholar] [CrossRef]
- Voyiatzaki, C.; Papailia, S.I.; Venetikou, M.S.; Pouris, J.; Tsoumani, M.E.; Papageorgiou, E.G. Climate Changes Exacerbate the Spread of Ixodes ricinus and the Occurrence of Lyme Borreliosis and Tick-Borne Encephalitis in Europe—How Climate Models Are Used as a Risk Assessment Approach for Tick-Borne Diseases. Int. J. Environ. Res. Public Health 2022, 19, 6516. [Google Scholar] [CrossRef]
- Gale, P.; Drew, T.; Phipps, L.P.; David, G.; Wooldridge, M. The effect of climate change on the occurrence and prevalence of livestock diseases in Great Britain: A review. J. Appl. Microbiol. 2009, 106, 1409–1423. [Google Scholar] [CrossRef]
- Woldehiwet, Z. The natural history of Anaplasma phagocytophilum. Vet. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef]
- Uminski, K.; Kadkhoda, K.; Houston, B.L.; Lopez, A.; MacKenzie, L.J.; Lindsay, R.; Walkty, A.; Embil, J.; Zarychanski, R. Anaplasmosis: An emerging tick-borne disease of importance in Canada. IDCases 2018, 14, e00472. [Google Scholar] [CrossRef]
- Gray, J.S.; Ogden, N.H. Ticks, Human Babesiosis and Climate Change. Pathogens 2021, 10, 1430. [Google Scholar] [CrossRef]
- Marques, R.; Krüger, R.F.; Peterson, A.T.; de Melo, L.F.; Vicenzi, N.; Jiménez-García, D. Climate change implications for the distribution of the babesiosis and anaplasmosis tick vector, Rhipicephalus (Boophilus) microplus. Vet. Res. 2020, 51, 81. [Google Scholar] [CrossRef]
- Wilson, A.J.; Mellor, P.S. Bluetongue in Europe: Past, present and future. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2669–2681. [Google Scholar] [CrossRef]
- Hughes, H.R.; Velez, J.O.; Fitzpatrick, K.; Davis, E.H.; Russell, B.J.; Lambert, A.J.; Staples, J.E.; Brault, A.C. Genomic Evaluation of the Genus Coltivirus Indicates Genetic Diversity among Colorado Tick Fever Virus Strains and Demarcation of a New Species. Diseases 2021, 9, 92. [Google Scholar] [CrossRef]
- Viral CNS Infections. In Hunter’s Tropical Medicine and Emerging Infectious Disease; Elsevier: Amsterdam, The Netherlands, 2013; pp. 344–379.
- Paddock, C.D.; Childs, J.E. Ehrlichia chaffeensis: A Prototypical Emerging Pathogen. Clin. Microbiol. Rev. 2003, 16, 37–64. [Google Scholar] [CrossRef]
- Marcdante, K.J.; Kliegman, R.M. Zoonoses and Vector Borne Infections. In Nelson Essentials of Pediatrics; Marcdante, K.J., Kliegman, R.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 439–447. ISBN 0323511457. [Google Scholar]
- Adler, B. History of leptospirosis and leptospira. Curr. Top. Microbiol. Immunol. 2015, 387, 1–9. [Google Scholar] [CrossRef]
- Lau, C.L.; Smythe, L.D.; Craig, S.B.; Weinstein, P. Climate change, flooding, urbanisation and leptospirosis: Fuelling the fire? Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 631–638. [Google Scholar] [CrossRef]
- Howard, S.; Ginsberg, P.; Jannelle Couret, P.M.; Jason Garrett, B.M.; Thomas, N.; Mather, P.; Roger, A.; Lebrun, P. Potential effects of climate change on tick-borne diseases in Rhode Island. Rhode Isl. Med. J. 2021, 104, 29–33. [Google Scholar]
- Steinbrink, A.; Brugger, K.; Margos, G.; Kraiczy, P.; Klimpel, S. The evolving story of Borrelia burgdorferi sensu lato transmission in Europe. Parasitol. Res. 2022, 121, 781–803. [Google Scholar] [CrossRef]
- Brownstein, J.S.; Holford, T.R.; Fish, D. Effect of Climate Change on Lyme Disease Risk in North America. Ecohealth 2005, 2, 38–46. [Google Scholar] [CrossRef]
- Bouchard, C.; Dibernardo, A.; Koffi, J.; Wood, H.; Leighton, P.; Lindsay, L. Increased risk of tick-borne diseases with climate and environmental changes. Can. Commun. Dis. Rep. 2019, 45, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, B. Dutch Q Fever Epidemic in a ‘One Health’ Context: Outbreaks, Seroprevalence and Occupational Risks. Ph.D. Thesis, University of Utrecht, Utrecht, The Netherlands, 2018. [Google Scholar]
- Alkishe, A.; Peterson, A.T. Climate change influences on the geographic distributional potential of the spotted fever vectors Amblyomma maculatum and Dermacentor andersoni. PeerJ 2022, 10, e13279. [Google Scholar] [CrossRef] [PubMed]
- Guzman, N.; Yarrarapu, S.N.S.; Beidas, S.O. Anaplasma Phagocytophilum; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Zinck, C.B.; Lloyd, V.K. Borrelia burgdorferi and Borrelia miyamotoi in Atlantic Canadian wildlife. PLoS ONE 2022, 17, e0262229. [Google Scholar] [CrossRef]
- Cameron, B.; Bierl, C.; Davenport, T.; Vollmer-Conna, U.; Hickie, I.; Wakefield, D.; Lloyd, A. Drought and Q fever: The association between trends in the incidence of infection and rainfall in rural Australia. In The Principles and Practice of Q Fever: The One Health Paradigm; Caetano Simões, J.C., Ferreira Anastácio, S., Jorge da Silva, G., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2017; ISBN 978-1-53610-851-4. [Google Scholar]
- Tan, T.S.; Hernandez-Jover, M.; Hayes, L.M.; Wiethoelter, A.K.; Firestone, S.M.; Stevenson, M.A.; Heller, J. Identifying scenarios and risk factors for Q fever outbreaks using qualitative analysis of expert opinion. Zoonoses Public Health 2022, 69, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Holekamp, K.; MacLachlan, N.J.; Frank, L.G.; Sawyer, M.; Osburn, B.I.; Mills, M.G.L.; Lerche, N.W.; Laurenson, M.K.; McNutt, J.W.; Alexander, K.A.; et al. Evidence of Natural Bluetongue Virus Infection among African Carnivores. Am. J. Trop. Med. Hyg. 1994, 51, 568–576. [Google Scholar] [CrossRef]
- More, S.; Bicout, D.; Bøtner, A.; Butterworth, A.; Depner, K.; Edwards, S.; Garin-Bastuji, B.; Good, M.; Gortázar Schmidt, C.; Michel, V.; et al. Assessment of listing and categorisation of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Bluetongue. EFSA J. 2017, 15, e04957. [Google Scholar] [CrossRef]
- Flahault, A.; de Castaneda, R.R.; Bolon, I. Climate change and infectious diseases. Public Health Rev. 2016, 37, 21. [Google Scholar] [CrossRef]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. Rev. Sci. Tech. 2008, 27, 499–510. [Google Scholar]
- Etxegarai-Legarreta, O.; Sanchez-Famoso, V. The Role of Beekeeping in the Generation of Goods and Services: The Interrelation between Environmental, Socioeconomic, and Sociocultural Utilities. Agriculture 2022, 12, 551. [Google Scholar] [CrossRef]
- Giliba, R.A.; Mpinga, I.H.; Ndimuligo, S.A.; Mpanda, M.M. Changing climate patterns risk the spread of Varroa destructor infestation of African honey bees in Tanzania. Ecol. Process. 2020, 9, 48. [Google Scholar] [CrossRef]
- Rowland, B.W.; Rushton, S.P.; Shirley, M.D.F.; Brown, M.A.; Budge, G.E. Identifying the climatic drivers of honey bee disease in England and Wales. Sci. Rep. 2021, 11, 21953. [Google Scholar] [CrossRef]
- Veen, N.; Yadav, A.S. Seasonal Incidence of European Foul Brood and Sac Brood Disease in Apis mellifera L. and their Correlation with Colony and Weather Parameters. Int. J. Curr. Microbiol. Appl. Sci. 2021, 10, 13–24. [Google Scholar] [CrossRef]
- Budge, G.E.; Simcock, N.K.; Holder, P.J.; Shirley, M.D.F.; Brown, M.A.; Van Weymers, P.S.M.; Evans, D.J.; Rushton, S.P. Chronic bee paralysis as a serious emerging threat to honey bees. Nat. Commun. 2020, 11, 2164. [Google Scholar] [CrossRef]
- Stephan, J.G.; de Miranda, J.R.; Forsgren, E. American foulbrood in a honeybee colony: Spore-symptom relationship and feedbacks between disease and colony development. BMC Ecol. 2020, 20, 16. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; FAO (Food and Agriculture Organization): Budapest, Hungary, 2020. [Google Scholar]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef]
- Lafferty, K.D. Marine Infectious Disease Ecology. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 473–496. [Google Scholar] [CrossRef]
- Hillebrand, H.; Brey, T.; Gutt, J.; Hagen, W.; Metfies, K.; Meyer, B.; Lewandowska, A. Climate Change: Warming Impacts on Marine Biodiversity. In Handbook on Marine Environment Protection; Springer International Publishing: Cham, Swizterland, 2018; pp. 353–373. [Google Scholar]
- Canonico, G.; Buttigieg, P.L.; Montes, E.; Muller-Karger, F.E.; Stepien, C.; Wright, D.; Benson, A.; Helmuth, B.; Costello, M.; Sousa-Pinto, I.; et al. Global Observational Needs and Resources for Marine Biodiversity. Front. Mar. Sci. 2019, 6, 367. [Google Scholar] [CrossRef]
- Bunse, C.; Lundin, D.; Karlsson, C.M.G.; Akram, N.; Vila-Costa, M.; Palovaara, J.; Svensson, L.; Holmfeldt, K.; González, J.M.; Calvo, E.; et al. Response of marine bacterioplankton pH homeostasis gene expression to elevated CO2. Nat. Clim. Change 2016, 6, 483–487. [Google Scholar] [CrossRef]
- Kling, J.D.; Lee, M.D.; Fu, F.; Phan, M.D.; Wang, X.; Qu, P.; Hutchins, D.A. Transient exposure to novel high temperatures reshapes coastal phytoplankton communities. ISME J. 2020, 14, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tsementzi, D.; Williams, T.C.; Juarez, D.L.; Blinebry, S.K.; Garcia, N.S.; Sienkiewicz, B.K.; Konstantinidis, K.T.; Johnson, Z.I.; Hunt, D.E. Environmental stability impacts the differential sensitivity of marine microbiomes to increases in temperature and acidity. ISME J. 2021, 15, 19–28. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Fu, F. Microorganisms and ocean global change. Nat. Microbiol. 2017, 2, 17058. [Google Scholar] [CrossRef]
- Abirami, B.; Radhakrishnan, M.; Kumaran, S.; Wilson, A. Impacts of global warming on marine microbial communities. Sci. Total Environ. 2021, 791, 147905. [Google Scholar] [CrossRef]
- Cohen, R.; James, C.; Lee, A.; Martinelli, M.; Muraoka, W.; Ortega, M.; Sadowski, R.; Starkey, L.; Szesciorka, A.; Timko, S.; et al. Marine Host-Pathogen Dynamics: Influences of Global Climate Change. Oceanography 2018, 31, 182–193. [Google Scholar] [CrossRef]
- Ben-Haim, Y.; Zicherman-Keren, M.; Rosenberg, E. Temperature-Regulated Bleaching and Lysis of the Coral Pocillopora damicornis by the Novel Pathogen Vibrio coralliilyticus. Appl. Environ. Microbiol. 2003, 69, 4236–4242. [Google Scholar] [CrossRef]
- Malek, J.C.; Byers, J.E. Responses of an oyster host (Crassostrea virginica) and its protozoan parasite (Perkinsus marinus) to increasing air temperature. PeerJ 2018, 6, e5046. [Google Scholar] [CrossRef]
- Marcos-López, M.; Gale, P.; Oidtmann, B.C.; Peeler, E.J. Assessing the Impact of Climate Change on Disease Emergence in Freshwater Fish in the United Kingdom. Transbound. Emerg. Dis. 2010, 57, 293–304. [Google Scholar] [CrossRef]
- De Souza Valente, C.; Wan, A.H.L. Vibrio and major commercially important vibriosis diseases in decapod crustaceans. J. Invertebr. Pathol. 2021, 181, 107527. [Google Scholar] [CrossRef]
- Firmino, J.; Furones, M.D.; Andree, K.B.; Sarasquete, C.; Ortiz-Delgado, J.B.; Asencio-Alcudia, G.; Gisbert, E. Contrasting outcomes of Vibrio harveyi pathogenicity in gilthead seabream, Sparus aurata and European seabass, Dicentrachus labrax. Aquaculture 2019, 511, 734210. [Google Scholar] [CrossRef]
- Li, X.; Yang, B.; Shi, C.; Wang, H.; Yu, R.; Li, Q.; Liu, S. Synergistic Interaction of Low Salinity Stress With Vibrio Infection Causes Mass Mortalities in the Oyster by Inducing Host Microflora Imbalance and Immune Dysregulation. Front. Immunol. 2022, 13, 859975. [Google Scholar] [CrossRef]
- Dorfmeier, E.M. Ocean Acidification and Disease: How Will a Changing Climate Impact Vibrio tubiashii Growth and Pathogenicity to Pacific Oyster Larvae? Master’s Thesis, University of Washington, Seattle, WA, USA, 2012. [Google Scholar]
- Bartlow, A.W.; Manore, C.; Xu, C.; Kaufeld, K.A.; Del Valle, S.; Ziemann, A.; Fairchild, G.; Fair, J.M. Forecasting Zoonotic Infectious Disease Response to Climate Change: Mosquito Vectors and a Changing Environment. Vet. Sci. 2019, 6, 40. [Google Scholar] [CrossRef]
- Raffel, T.R.; Romansic, J.M.; Halstead, N.T.; McMahon, T.A.; Venesky, M.D.; Rohr, J.R. Disease and thermal acclimation in a more variable and unpredictable climate. Nat. Clim. Change 2013, 3, 146–151. [Google Scholar] [CrossRef]
- Tong, S.; Ebi, K.; Olsen, J. Infectious disease, the climate, and the future. Environ. Epidemiol. 2021, 5, e133. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.B. Epidemics on the move: Climate change and infectious disease. PLoS Biol. 2020, 18, e3001013. [Google Scholar] [CrossRef] [PubMed]
- Duchenne-Moutien, R.A.; Neetoo, H. Climate Change and Emerging Food Safety Issues: A Review. J. Food Prot. 2021, 84, 1884–1897. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the Emergence of Candida auris: Climate Change, Azoles, Swamps, and Birds. mBio 2019, 10, e01397-19. [Google Scholar] [CrossRef] [PubMed]
- Kurane, I. The effect of global warming on infectious diseases. Osong Public Health Res. Perspect. 2010, 1, 4–9. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef]
- Coates, S.J.; Norton, S.A. The effects of climate change on infectious diseases with cutaneous manifestations. Int. J. Women’s Dermatol. 2021, 7, 8–16. [Google Scholar] [CrossRef]
- Semenza, J.C.; Paz, S. Climate change and infectious disease in Europe: Impact, projection and adaptation. Lancet Reg. Health Eur. 2021, 9, 100230. [Google Scholar] [CrossRef]
- Semenza, J.C.; Suk, J.E. Vector-borne diseases and climate change: A European perspective. FEMS Microbiol. Lett. 2018, 365, fnx244. [Google Scholar] [CrossRef]
- Caminade, C.; McIntyre, K.M.; Jones, A.E. Impact of recent and future climate change on vector-borne diseases. Ann. N. Y. Acad. Sci. 2019, 1436, 157–173. [Google Scholar] [CrossRef]
- The Lancet Microbe. Climate change: Fires, floods, and infectious diseases. Lancet Microbe 2021, 2, e415. [Google Scholar] [CrossRef]
- Fouladkhah, A. Changing climate: A ‘threat multiplier’ for foodborne and waterborne infectious diseases and antibiotic resistance. Res. Outreach 2020, 114, 130–133. [Google Scholar] [CrossRef]
- Williams, P.C.M.; Bartlett, A.W.; Howard-Jones, A.; McMullan, B.; Khatami, A.; Britton, P.N.; Marais, B.J. Impact of climate change and biodiversity collapse on the global emergence and spread of infectious diseases. J. Paediatr. Child Health 2021, 57, 1811–1818. [Google Scholar] [CrossRef]
- Koch, L.K.; Kochmann, J.; Klimpel, S.; Cunze, S. Modeling the climatic suitability of leishmaniasis vector species in Europe. Sci. Rep. 2017, 7, 13325. [Google Scholar] [CrossRef]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl. Trop. Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef]
- Ashford, R.W. The leishmaniases as emerging and reemerging zoonoses. Int. J. Parasitol. 2000, 30, 1269–1281. [Google Scholar] [CrossRef]
- Hotez, P.J.; Bottazzi, M.E.; Franco-Paredes, C.; Ault, S.K.; Periago, M.R. The Neglected Tropical Diseases of Latin America and the Caribbean: A Review of Disease Burden and Distribution and a Roadmap for Control and Elimination. PLoS Negl. Trop. Dis. 2008, 2, e300. [Google Scholar] [CrossRef]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; Boer, M. den Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef]
- Impactos del Cambio Climático en la Salud; Ministerio De Sanidad Igualdad y Servicios: Madrid, Spain, 2013.
- Pozio, E. How globalization and climate change could affect foodborne parasites. Exp. Parasitol. 2020, 208, 107807. [Google Scholar] [CrossRef]
- Kuhn, K.G.; Nygård, K.M.; Guzman-Herrador, B.; Sunde, L.S.; Rimhanen-Finne, R.; Trönnberg, L.; Jepsen, M.R.; Ruuhela, R.; Wong, W.K.; Ethelberg, S. Campylobacter infections expected to increase due to climate change in Northern Europe. Sci. Rep. 2020, 10, 13874. [Google Scholar] [CrossRef] [PubMed]
- Semenza, J.C.; Herbst, S.; Rechenburg, A.; Suk, J.E.; Höser, C.; Schreiber, C.; Kistemann, T. Climate Change Impact Assessment of Food and Waterborne Diseases. Crit. Rev. Environ. Sci. Technol. 2012, 42, 857–890. [Google Scholar] [CrossRef]
- Morgado, M.E.; Jiang, C.; Zambrana, J.; Upperman, C.R.; Mitchell, C.; Boyle, M.; Sapkota, A.R.; Sapkota, A. Climate change, extreme events, and increased risk of salmonellosis: Foodborne diseases active surveillance network (FoodNet), 2004–2014. Environ. Health 2021, 20, 105. [Google Scholar] [CrossRef] [PubMed]
- Semenza, J.C. Cascading risks of waterborne diseases from climate change. Nat. Immunol. 2020, 21, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Chala, B.; Hamde, F. Emerging and Re-emerging Vector-Borne Infectious Diseases and the Challenges for Control: A Review. Front. Public Health 2021, 9, 715759. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef]
- Trærup, S.L.M.; Ortiz, R.A.; Markandya, A. The costs of climate change: A study of cholera in Tanzania. Int. J. Environ. Res. Public Health 2011, 8, 4386–4405. [Google Scholar] [CrossRef]
- Riebesell, U.; Aberle-Malzahn, N.; Achterberg, E.P.; Algueró-Muñiz, M.; Alvarez-Fernandez, S.; Arístegui, J.; Bach, L.T.; Boersma, M.; Boxhammer, T.; Guan, W.; et al. Toxic algal bloom induced by ocean acidification disrupts the pelagic food web. Nat. Clim. Change 2018, 8, 1082–1086. [Google Scholar] [CrossRef]
- Barreiro, C.; Barredo, J.L. Worldwide Clinical Demand for Antibiotics: Is It a Real Countdown? Methods Mol. Biol. 2021, 2296, 3–15. [Google Scholar] [CrossRef]
- O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of nations. Rev. Antimicrob. Resist. 2014, 1–18. [Google Scholar]
- IACG. No Time to Wait: Securing the Future from Drug-Resistant Infections; WHO (World Health Organization): New York, NY, USA, 2019. [Google Scholar]
- McGough, S.F.; MacFadden, D.R.; Hattab, M.W.; Mølbak, K.; Santillana, M. Rates of increase of antibiotic resistance and ambient temperature in Europe: A cross-national analysis of 28 countries between 2000 and 2016. Eurosurveillance 2020, 25, 1900414. [Google Scholar] [CrossRef]
- Barreiro, C.; Gutiérrez, S.; Olivera, E.R. Fungal Horizontal Gene Transfer: A History Beyond the Phylogenetic Kingdoms. In Horizontal Gene Transfer; Villa, T., Viñas, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 315–336. ISBN 978-3-030-21861-4. [Google Scholar]
- Souli, M.; Galani, I.; Giamarellou, H. Emergence of extensively drug-resistant and pandrug-resistant Gram-negative bacilli in Europe. Eurosurveillance 2008, 13, 19045. [Google Scholar] [CrossRef]
- Gifford, D.R.; Moss, E.; MacLean, R.C. Environmental variation alters the fitness effects of rifampicin resistance mutations in Pseudomonas aeruginosa. Evolution 2016, 70, 725–730. [Google Scholar] [CrossRef]
- Rodríguez-Verdugo, A.; Lozano-Huntelman, N.; Cruz-Loya, M.; Savage, V.; Yeh, P. Compounding Effects of Climate Warming and Antibiotic Resistance. iScience 2020, 23, 101024. [Google Scholar] [CrossRef]
- MacFadden, D.R.; McGough, S.F.; Fisman, D.; Santillana, M.; Brownstein, J.S. Antibiotic Resistance Increases with Local Temperature. Nat. Clim. Change 2018, 8, 510–514. [Google Scholar] [CrossRef]
- Burnham, J.P. Climate change and antibiotic resistance: A deadly combination. Ther. Adv. Infect. Dis. 2021, 8, 2049936121991374. [Google Scholar] [CrossRef]
- Waits, A.; Emelyanova, A.; Oksanen, A.; Abass, K.; Rautio, A. Human infectious diseases and the changing climate in the Arctic. Environ. Int. 2018, 121, 703–713. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Antibiotic resistance and its cost: Is it possible to reverse resistance? Nat. Rev. Microbiol. 2010, 8, 260–271. [Google Scholar] [CrossRef]
- Rodríguez-Verdugo, A.; Gaut, B.S.; Tenaillon, O. Evolution of Escherichia coli rifampicin resistance in an antibiotic-free environment during thermal stress. BMC Evol. Biol. 2013, 13, 50. [Google Scholar] [CrossRef]
- Tan, E.C.D.; Lamers, P. Circular Bioeconomy Concepts—A Perspective. Front. Sustain. 2021, 2, 701509. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Cellini, A.; Donati, I.; Pastore, C.; Onofrietti, C.; Spinelli, F. Facing Climate Change: Application of Microbial Biostimulants to Mitigate Stress in Horticultural Crops. Agronomy 2020, 10, 794. [Google Scholar] [CrossRef]
- Ibáñez, A.; Diez-Galán, A.; Cobos, R.; Calvo-Peña, C.; Barreiro, C.; Medina-Turienzo, J.; Sánchez-García, M.; Coque, J.J.R. Using Rhizosphere Phosphate Solubilizing Bacteria to Improve Barley (Hordeum vulgare) Plant Productivity. Microorganisms 2021, 9, 1619. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Zhou, Y.; Suenaga, T.; Oba, K.; Lu, J.; Wang, G.; Zhang, L.; Yoon, S.; Terada, A. Organic carbon determines nitrous oxide consumption activity of clade I and II nosZ bacteria: Genomic and biokinetic insights. Water Res. 2022, 209, 117910. [Google Scholar] [CrossRef] [PubMed]
- Bueno, E.; Mania, D.; Mesa, S.; Bedmar, E.J.; Frostegård, Å.; Bakken, L.R.; Delgado, M.J. Regulation of the Emissions of the Greenhouse Gas Nitrous Oxide by the Soybean Endosymbiont Bradyrhizobium diazoefficiens. Int. J. Mol. Sci. 2022, 23, 1486. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Cruz, S.; Vaksmaa, A.; Horn, M.A.; Niemann, H.; Pijuan, M.; Ho, A. Methanotrophs: Discoveries, Environmental Relevance, and a Perspective on Current and Future Applications. Front. Microbiol. 2021, 12, 678057. [Google Scholar] [CrossRef]
- Aimen, H.; Khan, A.S.; Kanwal, N. Methanotrophs: The Natural Way to Tackle Greenhouse Effect. J. Bioremediat. Biodegrad. 2018, 9, 432. [Google Scholar] [CrossRef]
- Gayathri, R.; Mahboob, S.; Govindarajan, M.; Al-Ghanim, K.A.; Ahmed, Z.; Al-Mulhm, N.; Vodovnik, M.; Vijayalakshmi, S. A review on biological carbon sequestration: A sustainable solution for a cleaner air environment, less pollution and lower health risks. J. King Saud Univ. 2021, 33, 101282. [Google Scholar] [CrossRef]
- Paul, V.; Chandra Shekharaiah, P.S.; Kushwaha, S.; Sapre, A.; Dasgupta, S.; Sanyal, D. Role of Algae in CO2 Sequestration Addressing Climate Change: A Review. In Renewable Energy and Climate Change; Deb, D., Dixit, A., Chandra, L., Eds.; Springer: Singapore, 2020; pp. 257–265. [Google Scholar]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef]
- Wang, G.; Chai, K.; Wu, J.; Liu, F. Effect of Pseudomonas putida on the degradation of epoxy resin varnish coating in seawater. Int. Biodeterior. Biodegrad. 2016, 115, 156–163. [Google Scholar] [CrossRef]
- Sanluis-Verdes, A.; Colomer-Vidal, P.; Rodriguez-Ventura, F.; Bello-Villarino, M.; Spinola-Amilibia, M.; Ruiz-Lopez, E.; Illanes-Vicioso, R.; Castroviejo, P.; Cigliano, R.A.; Montoya, M.; et al. Wax worm saliva and the enzymes therein are the key to polyethylene degradation by Galleria mellonella. Nat. Commun. 2022, 13, 5568. [Google Scholar] [CrossRef]
- Zeenat; Elahi, A.; Bukhari, D.A.; Shamim, S.; Rehman, A. Plastics degradation by microbes: A sustainable approach. J. King Saud Univ. 2021, 33, 101538. [Google Scholar] [CrossRef]
- Junaid, F.; Khawaja, L.A.; Ali, S. Single cell protein as a potential meat substitute: A review. World J. Pharm. Res. 2020, 9, 141–161. [Google Scholar]
- Gamarra-Castillo, O.; Echeverry-Montaña, N.; Marbello-Santrich, A.; Hernández-Carrión, M.; Restrepo, S. Meat Substitute Development from Fungal Protein (Aspergillus oryzae). Foods 2022, 11, 2940. [Google Scholar] [CrossRef]
- Yakimov, M.M.; Golyshin, P.N.; Lang, S.; Moore, E.R.B.; Abraham, W.-R.; Lünsdorf, H.; Timmis, K.N. Alcanivorax borkumensis gen. nov., sp. nov., a new, hydrocarbon-degrading and surfactant-producing marine bacterium. Int. J. Syst. Evol. Microbiol. 1998, 48, 339–348. [Google Scholar] [CrossRef]
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L.; et al. Deep-Sea Oil Plume Enriches Indigenous Oil-Degrading Bacteria. Science 2010, 330, 204–208. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibáñez, A.; Garrido-Chamorro, S.; Barreiro, C. Microorganisms and Climate Change: A Not So Invisible Effect. Microbiol. Res. 2023, 14, 918-947. https://doi.org/10.3390/microbiolres14030064
Ibáñez A, Garrido-Chamorro S, Barreiro C. Microorganisms and Climate Change: A Not So Invisible Effect. Microbiology Research. 2023; 14(3):918-947. https://doi.org/10.3390/microbiolres14030064
Chicago/Turabian StyleIbáñez, Ana, Sonia Garrido-Chamorro, and Carlos Barreiro. 2023. "Microorganisms and Climate Change: A Not So Invisible Effect" Microbiology Research 14, no. 3: 918-947. https://doi.org/10.3390/microbiolres14030064
APA StyleIbáñez, A., Garrido-Chamorro, S., & Barreiro, C. (2023). Microorganisms and Climate Change: A Not So Invisible Effect. Microbiology Research, 14(3), 918-947. https://doi.org/10.3390/microbiolres14030064