
Probiotic Potential of Lactic Acid Bacterial Strains Isolated from Human Oral Microbiome

Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Oral Samples
2.2. Identification of Lactic Acid Bacteria
2.3. Enzymatic Activity
2.4. Antimicrobial Activity against Test Pathogens
2.5. Survival Ability in Simulated Oral Conditions
2.6. Survival Ability in the Presence of Pepsin
2.7. Growth Dynamics in the Presence of Stress Factors
2.8. Autoaggregation and Mucin Binding Capability
2.9. Data Analyses
3. Results and Discussion
3.1. Isolation and Identification of Lactic Acid Bacteria
3.2. Enzymatic Activity
3.3. Antimicrobial Activity against Test Pathogens
3.4. Survival Ability in Simulated Oral Conditions
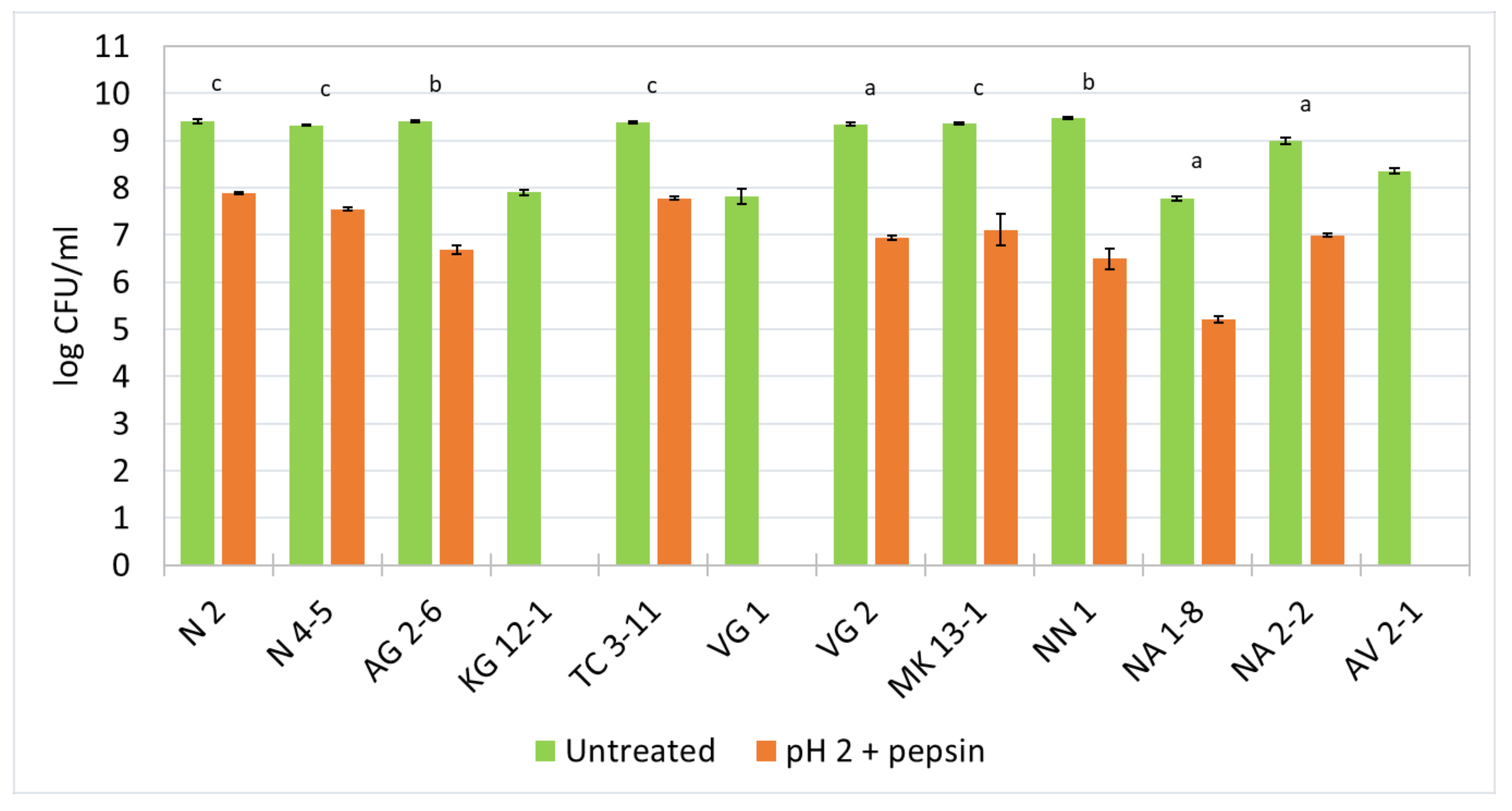
3.5. Survival Ability in Low pH and the Presence of Pepsin
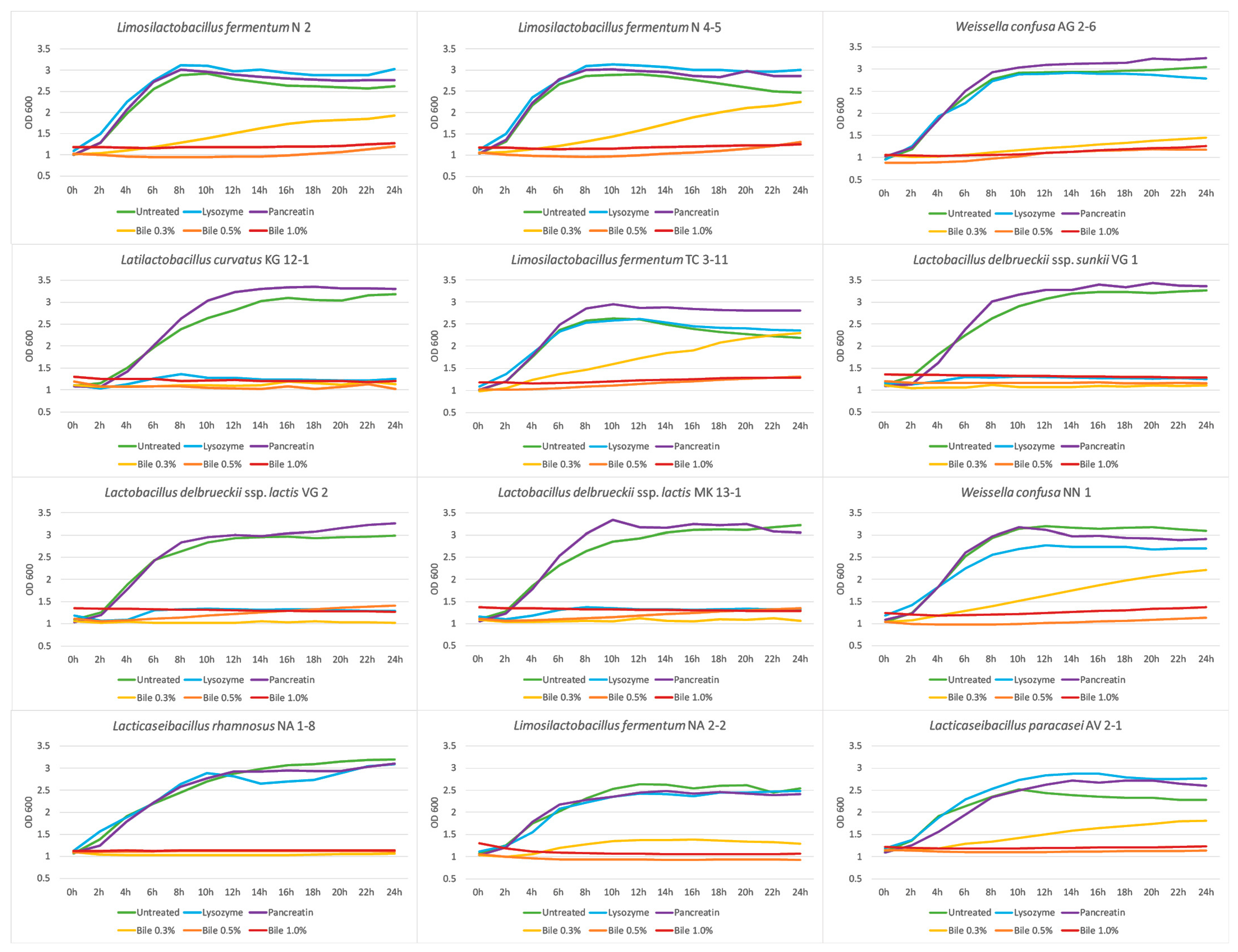
3.6. Growth Dynamics in the Presence of GIT Stress Factors
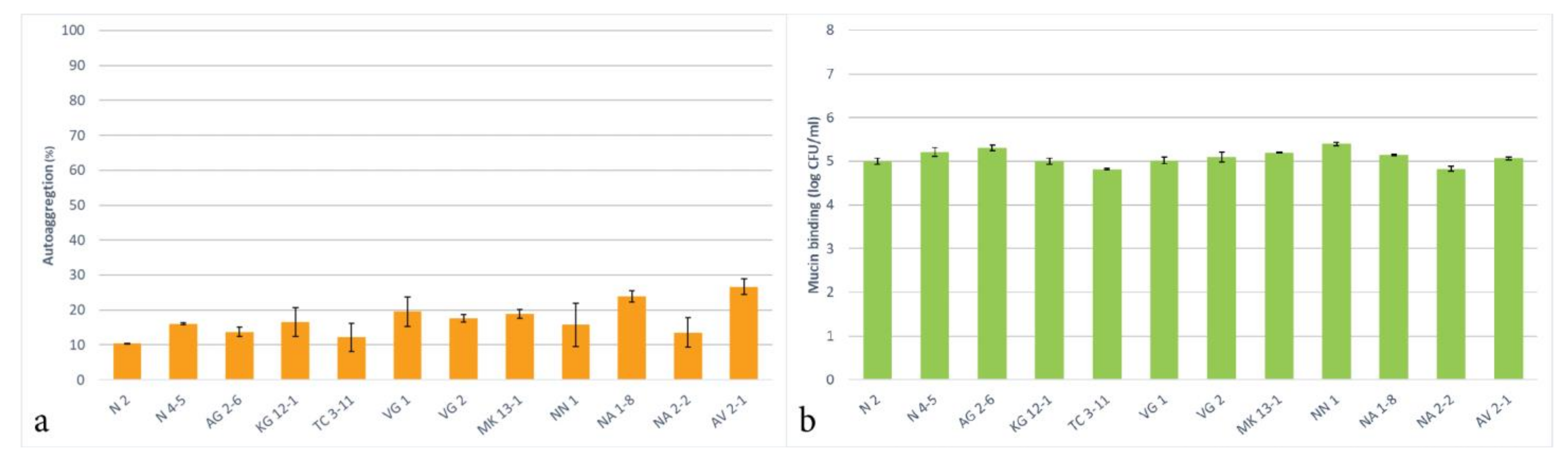
3.7. Autoaggregation and Mucin Binding Capability
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, L.; Xu, T.; Huang, G.; Jian, S.; Gu, Y.; Chen, F. Oral microbiomes: More and more importance in oral cavity and whole body. Protein Cell 2018, 9, 488–500. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations/World Health Organization FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food: London, ON, Canada, 2002. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Ayivi, R.D.; Ibrahim, S.A. Lactic acid bacteria: An essential probiotic and starter culture for the production of yoghurt. Int. J. Food Sci. Technol. 2022, 57, 7008–7025. [Google Scholar] [CrossRef]
- Sanders, M.E. Probiotics: Definition, sources, selection, and uses. Clin. Infect. Dis. 2008, 46, 58–61. [Google Scholar] [CrossRef]
- Parker, C.T.; Tindall, B.J.; Garrity, G.M. International Code of Nomenclature of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2019, 69, 1–111. [Google Scholar] [CrossRef]
- Gad, G.F.; Abdel-Hamid, A.M.; Farag, Z.S. Antibiotic resistance in lactic acid bacteria isolated from some pharmaceutical and dairy products. Braz. J. Microbiol. 2014, 45, 25–33. [Google Scholar] [CrossRef]
- Pimentel, T.C.; da Costa, W.K.A.; Delfino, T.P.C.; de Oliveira, S.M.L.; Sivieri, K.; Magnani, M. Chapter 5—Foods and supplements as probiotic delivery vehicles. In Probiotics for Human Nutrition in Health and Disease; de Souza, E.L., de Brito Alves, J.L., Fusco, V., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 115–142. [Google Scholar] [CrossRef]
- Un-Nisa, A.; Khan, A.; Zakria, M.; Siraj, S.; Ullah, S.; Tipu, M.K.; Ikram, M.; Kim, M.O. Updates on the Role of Probiotics against Different Health Issues: Focus on Lactobacillus. Int. J. Mol. Sci. 2023, 24, 142. [Google Scholar] [CrossRef]
- Ahmed, S.; Singh, S.; Singh, V.; Roberts, K.D.; Zaidi, A.; Rodriguez-Palacios, A. The Weissella Genus: Clinically Treatable Bacteria with Antimicrobial/Probiotic Effects on Inflammation and Cancer. Microorganisms 2022, 10, 2427. [Google Scholar] [CrossRef]
- Tekce, M.; Ince, G.; Gursoy, H.; Dirikan Ipci, S.; Cakar, G.; Kadir, T.; Yilmaz, S. Clinical and microbiological effects of probiotic lozenges in the treatment of chronic periodontitis: A 1-year follow-up study. J. Clin. Periodontol. 2015, 42, 363–372. [Google Scholar] [CrossRef]
- Sabatini, S.; Lauritano, D.; Candotto, V.; Silvestre, F.J.; Nardi, G.M. Oral probiotics in the management of gingivitis in diabetic patients: A double blinded randomized controlled study. J. Biol. Regul. Homeost. Agents 2017, 31, 197–202. [Google Scholar]
- Keller, M.K.; Brandsborg, E.; Holmstrom, K.; Twetman, S. Effect of tablets containing probiotic candidate strains on gingival inflammation and composition of the salivary microbiome: A randomised controlled trial. Benef. Microbes 2018, 9, 487–494. [Google Scholar] [CrossRef]
- Nadkerny, P.V.; Ravishankar, P.L.; Pramod, V.; Agarwal, L.A.; Bhandari, S. A comparative evaluation of the efficacy of probiotic and chlorhexidine mouthrinses on clinical inflammatory parameters of gingivitis: A randomized controlled clinical study. J. Indian Soc. Periodontol. 2015, 19, 633–639. [Google Scholar] [CrossRef]
- Grusovin, M.G.; Bossini, S.; Calza, S.; Cappa, V.; Garzetti, G.; Scotti, E.; Gherlone, E.F.; Mensi, M. Clinical efficacy of Lactobacillus reuteri-containing lozenges in the supportive therapy of generalized periodontitis stage III and IV, grade C: 1-year results of a double-blind randomized placebo-controlled pilot study. Clin. Oral Investig. 2020, 24, 2015–2024. [Google Scholar] [CrossRef]
- Parahitiyawa, N.; Scully, C.; Leung, W.; Yam, W.; Jin, L.; Samaranayake, L. Exploring the oral bacterial flora: Current status and future directions. Oral Dis. 2010, 16, 136–145. [Google Scholar] [CrossRef]
- Meurman, J.H.; Stamatova, I. Probiotics: Evidence of Oral Health Implications. Folia Med. 2018, 60, 21–29. [Google Scholar] [CrossRef]
- Bosch, J.A.; Turkenburg, M.; Nazmi, K.; Veerman, C.I.; de Geus, J.C.; Nieuw Amerongen, A.V. Stress as a determinant of saliva-mediated adherence and coadherence of oral and nonoral microorganisms. Psychosom. Med. 2003, 65, 604–612. [Google Scholar] [CrossRef]
- Stamatova, I.; Kari, K.; Vladimirov, S.; Meurman, J.H. In vitro evaluation of yoghurt starter lactobacilli and Lactobacillus rhamnosus GG adhesion to saliva-coated surfaces. Oral Microbiol. Immunol. 2009, 24, 218–223. [Google Scholar] [CrossRef]
- Kilian, M.; Chapple, I.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Ellison, R.T., III; Giehl, T.J. Killing of gram-negative bacteria by lactoferrin and lysozyme. J. Clin. Investig. 1991, 88, 1080–1091. [Google Scholar] [CrossRef]
- Lynge Pedersen, A.M.; Belstrøm, D. The role of natural salivary defences in maintaining a healthy oral microbiota. J. Dent. 2019, 80, 3–12. [Google Scholar] [CrossRef]
- Shah, N.P. Probiotic bacteria: Selective enumeration and survival in dairy foods. J. Dairy Sci. 2000, 83, 894–907. [Google Scholar] [CrossRef]
- Pitino, I.; Randazzo, C.L.; Mandalari, G.; Lo Curto, A.; Faulks, R.M.; Le Marc, Y.; Bisignano, C.; Caggia, C.; Wickham, M.S.J. Survival of Lactobacillus rhamnosus strains in the upper gastrointestinal tract. Food Microbiol. 2010, 27, 1121–1127. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rodríguez-Aparicio, L.B.; Rúa, J.; Martínez-Blanco, H.; Navasa, N.; García-Armesto, M.R.; Ferrero, M.Á. In vitro evaluation of physiological probiotic properties of different lactic acid bacteria strains of dairy and human origin. J. Funct. Foods 2012, 4, 531–541. [Google Scholar] [CrossRef]
- Quinto, E.J.; Jiménez, P.; Caro, I.; Tejero, J.; Mateo, J.; Girbés, T. Probiotic lactic acid bacteria: A review. Food Nutr. Sci. 2014, 5, 1765. [Google Scholar] [CrossRef]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef]
- Nishiyama, K.; Sugiyama, M.; Mukai, T. Adhesion Properties of Lactic Acid Bacteria on Intestinal Mucin. Microorganisms 2016, 4, 34. [Google Scholar] [CrossRef]
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva, R.C.d.; Ibrahim, S.A. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 202–232. [Google Scholar] [CrossRef]
- Van Tassell, M.L.; Miller, M.J. Lactobacillus adhesion to mucus. Nutrients 2011, 3, 613–636. [Google Scholar] [CrossRef]
- Bartholomew, J.W.; Mittwer, T. The Gram stain. Bacteriol. Rev. 1952, 16, 1–29. [Google Scholar] [CrossRef]
- Duke, P.B.; Jarvis, J.D. The catalase test—A cautionary tale. J. Med. Lab. Technol. 1972, 29, 203–204. [Google Scholar]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Marteau, P.; Minekus, M.; Havenaar, R.; Huis, J.H.J. Survival of Lactic Acid Bacteria in a Dynamic Model of the Stomach and Small Intestine: Validation and the Effects of Bile. J. Dairy Sci. 1997, 80, 1031–1037. [Google Scholar] [CrossRef]
- Bove, P.; Gallone, A.; Russo, P.; Capozzi, V.; Albenzio, M.; Spano, G.; Fiocco, D. Probiotic features of Lactobacillus plantarum mutant strains. Appl. Microbiol. Biotechnol. 2012, 96, 431–441. [Google Scholar] [CrossRef]
- Damodharan, K.; Lee, Y.; Palaniyandi, S.A.; Yang, S.; Suh, J.W. Preliminary probiotic and technological characterization of Pediococcus pentosaceus strain KID7 and in vivo assessment of its cholesterol-lowering activity. Front. Microbiol. 2015, 6, 768. [Google Scholar] [CrossRef]
- Plessas, S.; Kiousi, D.E.; Rathosi, M.; Alexopoulos, A.; Kourkoutas, Y.; Mantzourani, I.; Galanis, A.; Bezirtzoglou, E. Isolation of a Lactobacillus paracasei Strain with Probiotic Attributes from Kefir Grains. Biomedicines 2020, 8, 594. [Google Scholar] [CrossRef]
- Jia, G.C.; Che, N.; Xia, Y.J.; Lai, P.F.; Xiong, Z.Q.; Wang, G.Q.; Zhang, H.; Ai, L.Z. Adhesion to pharyngeal epithelium and modulation of immune response: Lactobacillus salivarius AR809, a potential probiotic strain isolated from the human oral cavity. J. Dairy Sci. 2019, 102, 6738–6749. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Du, M.; Yi, H.; Guo, C.; Tuo, Y.; Han, X.; Li, J.; Zhang, L.; Yang, L. Antimicrobial activity against Shigella sonnei and probiotic properties of wild lactobacilli from fermented food. Microbiol. Res. 2011, 167, 27–31. [Google Scholar] [CrossRef]
- Unban, K.; Chaichana, W.; Baipong, S.; Abdullahi, A.D.; Kanpiengjai, A.; Shetty, K.; Khanongnuch, C. Probiotic and Antioxidant Properties of Lactic Acid Bacteria Isolated from Indigenous Fermented Tea Leaves (Miang) of North Thailand and Promising Application in Synbiotic Formulation. Fermentation 2021, 7, 195. [Google Scholar] [CrossRef]
- Monteiro, C.R.A.V.; do Carmo, M.S.; Melo, B.O.; Alves, M.S.; dos Santos, C.I.; Monteiro, S.G.; Bomfim, M.R.Q.; Fernandes, E.S.; Monteiro-Neto, V. In Vitro Antimicrobial Activity and Probiotic Potential of Bifidobacterium and Lactobacillus against Species of Clostridium. Nutrients 2019, 11, 448. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Jussiaux, F.; Miot-Sertier, C.; Nguyen-Lopez, D.; Badet, C.; Samot, J. Reliability of MALDI-TOF mass spectrometry to identify oral isolates of Streptococcus salivarius and Lactobacillus spp. Arch. Oral Biol. 2021, 121, 104983. [Google Scholar] [CrossRef]
- Kim, H.; Jeong, Y.; Kim, J.E.; Kim, Y.; Paek, N.S.; Kang, C.H. Anti-obesity Potential of Lactobacillus spp. Isolated from Infant Feces. Biotechnol. Bioprocess E 2021, 26, 575–585. [Google Scholar] [CrossRef]
- Mudryk, Z.J.; Podgórska, B. Enzymatic Activity of Bacterial Strains Isolated from Marine Beach Sediments. Pol. J. Environ. Stud. 2006, 15, 441–448. [Google Scholar]
- Song, M.W.; Chung, Y.; Kim, K.T.; Hong, W.S.; Chang, H.J.; Paik, H.D. Probiotic characteristics of Lactobacillus brevis B13-2 isolated from kimchi and investigation of antioxidant and immune-modulating abilities of its heat-killed cells. LWT 2020, 128, 109452. [Google Scholar] [CrossRef]
- Gao, Z.; Daliri, E.B.M.; Wang, J.; Liu, D.; Chen, S.; Ye, X.; Ding, T. Inhibitory effect of lactic acid bacteria on foodborne pathogens: A review. J. Food Prot. 2019, 82, 441–453. [Google Scholar] [CrossRef]
- Ren, D.; Zhu, J.; Gong, S.; Liu, H.; Yu, H. Antimicrobial Characteristics of Lactic Acid Bacteria Isolated from Homemade Fermented Foods. Biomed. Res. Int. 2018, 2018, 5416725. [Google Scholar] [CrossRef]
- Matevosyan, L.; Bazukyan, I.; Trchounian, A. Antifungal and antibacterial effects of newly created lactic acid bacteria associations depending on cultivation media and duration of cultivation. BMC Microbiol. 2019, 19, 102. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Donelli, G. Probiotics to counteract biofilm-associated infections: Promising and conflicting data. Int. J. Oral Sci. 2014, 6, 189–194. [Google Scholar] [CrossRef]
- Sookkhee, S.; Chulasiri, M.; Prachyabrued, W. Lactic acid bacteria from healthy oral cavity of Thai volunteers: Inhibition of oral pathogens. J. Appl. Microbiol. 2001, 90, 172–179. [Google Scholar] [CrossRef]
- Strahinic, I.; Busarcevic, M.; Pavlica, D.; Milasin, J.; Golic, N.; Topisirovic, L. Molecular and biochemical characterizations of human oral lactobacilli as putative probiotic candidates. Oral Microbiol. Immunol. 2007, 22, 111–117. [Google Scholar] [CrossRef]
- Itapary dos Santos, C.; Ramos França, Y.; Duarte Lima Campos, C.; Quaresma Bomfim, M.R.; Oliveira Melo, B.; Assunção Holanda, R.; Santos, V.L.; Gomes Monteiro, S.; Buozzi Moffa, E.; Souza Monteiro, A.; et al. Antifungal and Antivirulence Activity of Vaginal Lactobacillus Spp. Products against Candida Vaginal Isolates. Pathogens 2019, 8, 150. [Google Scholar] [CrossRef]
- Denkova, R.; Yanakieva, V.; Denkova, Z.; Nikolova, V.; Radeva, V. In vitro inhibitory activity of Bifidobacterium and Lactobacillus strains against Candida albicans. Bulg. J. Vet. Med. 2013, 16, 186–197. [Google Scholar]
- Liao, H.; Liu, S.; Wang, H.; Su, H.; Liu, Z. Enhanced antifungal activity of bovine lactoferrin-producing probiotic Lactobacillus casei in the murine model of vulvovaginal candidiasis. BMC Microbiol. 2019, 19, 7. [Google Scholar] [CrossRef]
- Ariningsih, I.; Ramona, Y.; Antara, N. Isolation, Screening, and Characterization of Probiotics (Lactic Acid Bacteria) Antagonistic Against Candida albicans. Metamorf. J. Biol. Sci. 2017, 4, 263. [Google Scholar] [CrossRef]
- Haukioja, A.; Yli-Knuuttila, H.; Loimaranta, V.; Kari, K.; Ouwehand, A.C.; Meurman, J.H.; Tenovuo, J. Oral adhesion and survival of probiotic and other lactobacilli and bifidobacteria in vitro. Oral Microbiol. Immunol. 2006, 21, 326–332. [Google Scholar] [CrossRef]
- Haukioja, A.; Loimaranta, V.; Tenovuo, J. Probiotic bacteria affect the composition of salivary pellicle and streptococcal adhesion in vitro. Oral Microbiol. Immunol. 2008, 23, 336–343. [Google Scholar] [CrossRef]
- Mantzourani, I.; Chondrou, P.; Bontsidis, C.; Karolidou, K.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Galanis, A.; Plessas, S. Assessment of the probiotic potential of lactic acid bacteria isolated from kefir grains: Evaluation of adhesion and antiproliferative properties in in vitro experimental systems. Ann. Microbiol. 2019, 69, 751–763. [Google Scholar] [CrossRef]
- Tokatlı, M.; Gülgör, G.; Bağder Elmacı, S.; Arslankoz İşleyen, N.; Özçelik, F. In Vitro Properties of Potential Probiotic Indigenous Lactic Acid Bacteria Originating from Traditional Pickles. BioMed Res. Int. 2015, 2015, 315819. [Google Scholar] [CrossRef]
- Fang, F.; Xu, J.; Li, Q.; Xia, X.; Du, G. Characterization of a Lactobacillus brevis strain with potential oral probiotic properties. BMC Microbiol. 2018, 18, 221. [Google Scholar] [CrossRef]
- Bosch, M.; Nart, J.; Audivert, S.; Bonachera, M.A.; Alemany, A.S.; Fuentes, M.C.; Cuñé, J. Isolation and characterization of probiotic strains for improving oral health. Arch. Oral Biol. 2012, 57, 539–549. [Google Scholar] [CrossRef]
- Khagwal, N.; Sharma, P.K.; Sharma, D.C. Screening and evaluation of Lactobacillus spp. for the development of potential probiotics. Afr. J. Microbiol. Res. 2014, 8, 1573–1579. [Google Scholar] [CrossRef]
- Jose, N.M.; Bunt, C.R.; Hussain, M.A. Comparison of Microbiological and Probiotic Characteristics of Lactobacilli Isolates from Dairy Food Products and Animal Rumen Contents. Microorganisms 2015, 3, 198–212. [Google Scholar] [CrossRef]
- Montville, T.J.; Matthews, K.R. Physiology, growth, and inhibition of microbes in foods. Food Microbiol. Fundam. Front. 2012, 2013, 3–18. [Google Scholar] [CrossRef]
- Alameri, F.; Tarique, M.; Osaili, T.; Obaid, R.; Abdalla, A.; Masad, R.; Al-Sbiei, A.; Fernandez-Cabezudo, M.; Liu, S.Q.; Al-Ramadi, B.; et al. Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects. Microorganisms 2022, 10, 389. [Google Scholar] [CrossRef]
- Aarti, C.; Khusro, A.; Varghese, R.; Arasu, M.V.; Agastian, P.; Al-Dhabi, N.A.; Ilavenil, S.; Choi, K.C. In vitro investigation on probiotic, anti-Candida, and antibiofilm properties of Lactobacillus pentosus strain LAP1. Arch. Oral Biol. 2018, 89, 99–106. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Cui, H.; Li, Y.; Sun, Y.; Qiu, H.J. Characterization of Lactic Acid Bacteria Isolated from the Gastrointestinal Tract of a Wild Boar as Potential Probiotics. Front. Vet. Sci. 2020, 7, 49. [Google Scholar] [CrossRef]
- Tuo, Y.; Yu, H.; Ai, L.; Wu, Z.; Guo, B.; Chen, W. Aggregation and adhesion properties of 22 Lactobacillus strains. J. Dairy Sci. 2013, 96, 4252–4257. [Google Scholar] [CrossRef]
- Campana, R.; van Hemert, S.; Baffone, W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathog. 2017, 9, 12. [Google Scholar] [CrossRef]
- Sharma, S.; Kandasamy, S.; Kavitake, D.; Shetty, P.H. Probiotic characterization and antioxidant properties of Weissella confusa KR780676, isolated from an Indian fermented food. LWT 2018, 97, 53–60. [Google Scholar] [CrossRef]
- Lakra, A.K.; Domdi, L.; Hanjon, G.; Tilwani, Y.M.; Arul, V. Some probiotic potential of Weissella confusa MD1 and Weissella cibaria MD2 isolated from fermented batter. LWT 2020, 125, 125–109261. [Google Scholar] [CrossRef]
- Moslem, P.; Hossein, N.; Mahdi, R.; Seyed, N.H.; Seyed, A.S. Lactobacillus rhamnosus Gorbach-Goldin (GG): A top well-researched probiotic strain. J. Med. Microbiol. 2017, 5, 46–59. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Cell Morphology | Gram-Stain | Peroxidase and Catalase | 16S rDNA Gene Sequence | % Identity | MALDI-TOF MS | Score |
|---|---|---|---|---|---|---|---|
| N 2 | Short rods | + | - | Limosilactobacillus fermentum | 99.7 | Limosilactobacillus fermentum | 1586 |
| N 4-5 | Short rods | + | - | Limosilactobacillus fermentum | 99.6 | Limosilactobacillus fermentum | 1822 |
| AG 2-6 | Short rods | + | - | Weissella confusa | 99.4 | Weissella confusa | 1837 |
| KG 12-1 | Short rods | + | - | Latilactobacillus curvatus | 99.6 | Latilactobacillus curvatus | 1189 |
| TC 3-11 | Short rods | + | - | Limosilactobacillus fermentum | 99.6 | Limosilactobacillus fermentum | 1234 |
| VG 1 | Long rods | + | - | Lactobacillus delbrueckii subsp. sunkii | 98.9 | Lactobacillus delbrueckii | 1324 |
| VG 2 | Long rods | + | - | Lactobacillus delbrueckii subsp. lactis | 98.7 | Lactobacillus delbrueckii | 1676 |
| MK 13-1 | Long rods | + | - | Lactobacillus delbrueckii subsp. lactis | 99.1 | Lactobacillus delbrueckii | 1193 |
| NN 1 | Short rods | + | - | Weissella confusa | 99.5 | Weissella confusa | 1490 |
| NA 1-8 | Short rods | + | - | Lacticaseibacillus rhamnosus | 99.9 | Lacticaseibacillus rhamnosus | 1324 |
| NA 2-2 | Short rods | + | - | Limosilactobacillus fermentum | 99.1 | Limosilactobacillus fermentum | 2774 |
| AV 2-1 | Short rods | + | - | Lacticaseibacillus paracasei | 99.8 | Lacticaseibacillus paracasei | 5533 |
| Enzyme Assayed for | N 2 | N 4-5 | AG 2-6 | KG 12-1 | TC 3-11 | VG 1 | VG 2 | MK 13-1 | NN 1 | NA 1-8 | NA 2-2 | AV 2-1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alkaline phosphatase | 1 | 1 | 2 | 2 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 3 |
| Esterase (C4) | 2 | 3 | 1 | 3 | 2 | 1 | 2 | 2 | 3 | 3 | 3 | 2 |
| Esterase lipase (C8) | 2 | 3 | 1 | 2 | 2 | 1 | 1 | 2 | 2 | 2 | 2 | 2 |
| Lipase (C14) | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 |
| Leucine arylamidase | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| Valine arylamidase | 3 | 3 | 2 | 5 | 3 | 3 | 2 | 2 | 3 | 5 | 3 | 5 |
| Cystine arylamidase | 3 | 3 | 2 | 2 | 3 | 2 | 2 | 2 | 2 | 3 | 2 | 3 |
| Trypsin | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| α-chymotrypsin | 0 | 0 | 3 | 3 | 0 | 4 | 4 | 4 | 4 | 4 | 4 | 3 |
| Acid phosphatase | 5 | 4 | 5 | 5 | 4 | 3 | 3 | 3 | 4 | 3 | 3 | 4 |
| Naphtol-AS-BIphosphohydrolase | 1 | 1 | 4 | 5 | 1 | 4 | 2 | 3 | 5 | 4 | 4 | 2 |
| α-galactosidase | 4 | 5 | 2 | 1 | 5 | 4 | 3 | 3 | 5 | 3 | 5 | 0 |
| β-galactosidase | 5 | 5 | 1 | 5 | 5 | 2 | 2 | 4 | 5 | 3 | 5 | 5 |
| β-glucuronidase | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| α-glucosidase | 0 | 0 | 4 | 5 | 5 | 5 | 5 | 5 | 4 | 5 | 3 | 5 |
| β-glucosidase | 0 | 0 | 1 | 3 | 0 | 3 | 3 | 3 | 4 | 3 | 1 | 3 |
| N-acetyl-β-glucosaminidase | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| α-mannosidase | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| α-fucosidase | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| Strains | Native CFS pH | Zone Diameter (mm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| E. coli | B. subtilis | B. cereus | S. aureus | S. epidermidis | P. aeruginosa | P. acnes | S. mutans | C. albicans | ||
| N 2 | 3.99 | 13.0 ± 0.5 | 12.0 ± 0.3 | 12.0 ± 0.5 | NZ | NZ | NZ | NZ | NZ | NZ |
| N 4-5 | 4.00 | 13.5 ± 0.5 | 11.0 ± 0.2 | 11.0 ± 0.3 | NZ | NZ | NZ | NZ | NZ | NZ |
| AG 2-6 | 3.89 | 12.0 ± 0.2 | NZ | NZ | NZ | 13.0 ± 0.1 | NZ | 12.0 ± 0.2 | 12.0 ± 0.3 | NZ |
| KG 12-1 | 3.90 | NZ | NZ | NZ | NZ | NZ | NZ | NZ | NZ | NZ |
| TC 3-11 | 3.72 | 13.0 ± 0.5 | 11.0 ± 0.3 | 12.0 ± 1.0 | NZ | NZ | 11.0 ± 0.1 | NZ | NZ | NZ |
| VG 1 | 3.42 | 17.0 ± 0.5 | 14.6 ± 0.6 | 14.6 ± 0.6 | 13.5 ± 0.5 | NZ | 11.0 ± 0.2 | NZ | NZ | NZ |
| VG 2 | 3.41 | 17.6 ± 0.4 | 15.3 ± 0.2 | 14.0 ± 0.1 | 13.0 ± 0.1 | NZ | 11.0 ± 0.2 | NZ | NZ | NZ |
| MK 13-1 | 3.54 | 17.0 ± 0.5 | 15.5 ± 0.5 | 14.6 ± 0.4 | 12.0 ± 0.1 | NZ | 11.0 ± 0.2 | NZ | NZ | NZ |
| NN 1 | 3.92 | 13.0 ± 0.2 | NZ | NZ | NZ | 12.0 ± 0.1 | NZ | 12.0 ± 0.1 | 13.0 ± 0.2 | NZ |
| NA 1-8 | 3.60 | 14.0 ± 0.1 | 13.2 ± 0.3 | NZ | NZ | NZ | NZ | NZ | NZ | NZ |
| NA 2-2 | 3.92 | 13.0 ± 0.3 | 12.5 ± 0.5 | NZ | NZ | NZ | 12.0 ± 1.0 | NZ | NZ | NZ |
| AV 2-1 | 3.44 | 13.0 ± 0.1 | NZ | 12.0 ± 0.1 | NZ | NZ | NZ | NZ | NZ | NZ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atanasov, N.; Evstatieva, Y.; Nikolova, D. Probiotic Potential of Lactic Acid Bacterial Strains Isolated from Human Oral Microbiome. Microbiol. Res. 2023, 14, 262-278. https://doi.org/10.3390/microbiolres14010021
Atanasov N, Evstatieva Y, Nikolova D. Probiotic Potential of Lactic Acid Bacterial Strains Isolated from Human Oral Microbiome. Microbiology Research. 2023; 14(1):262-278. https://doi.org/10.3390/microbiolres14010021
Chicago/Turabian StyleAtanasov, Nikola, Yana Evstatieva, and Dilyana Nikolova. 2023. "Probiotic Potential of Lactic Acid Bacterial Strains Isolated from Human Oral Microbiome" Microbiology Research 14, no. 1: 262-278. https://doi.org/10.3390/microbiolres14010021
APA StyleAtanasov, N., Evstatieva, Y., & Nikolova, D. (2023). Probiotic Potential of Lactic Acid Bacterial Strains Isolated from Human Oral Microbiome. Microbiology Research, 14(1), 262-278. https://doi.org/10.3390/microbiolres14010021