
In Vitro Inhibition of Zika Virus Replication with Amantadine and Rimantadine Hydrochlorides


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Antivirals
2.3. Antiviral Activity Assay
2.4. Cytotoxicity Assay
2.5. Statistics
3. Results
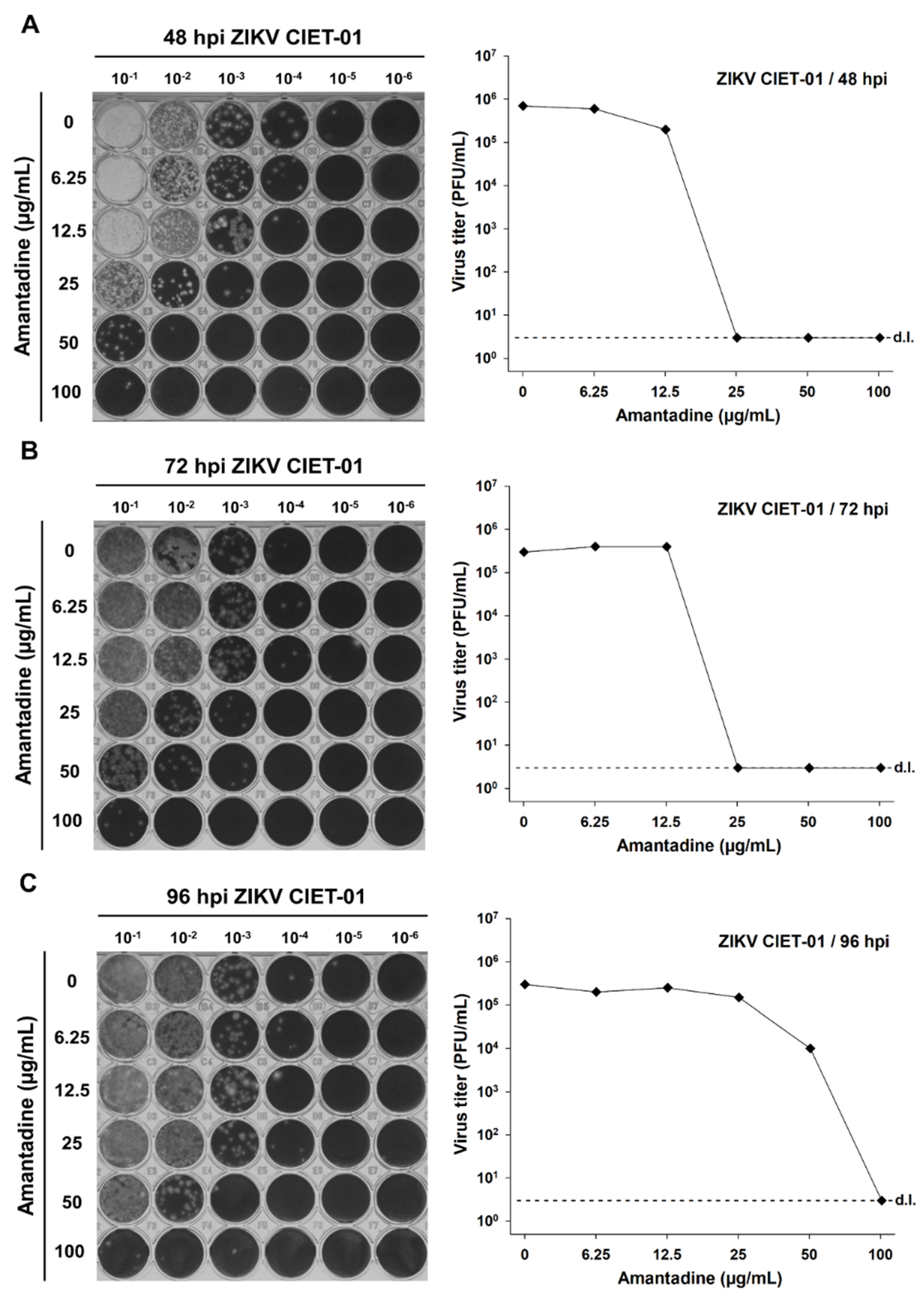
3.1. Amantadine and Rimantadine Hydrochlorides Have Antiviral Activity against a ZIKV Clinical Isolate at Different Times Post-Infection
3.2. Antiviral Activity against ZIKV Is Not Explained by a Cytotoxic Effect Induced by the Tested Concentrations of Adamantanes
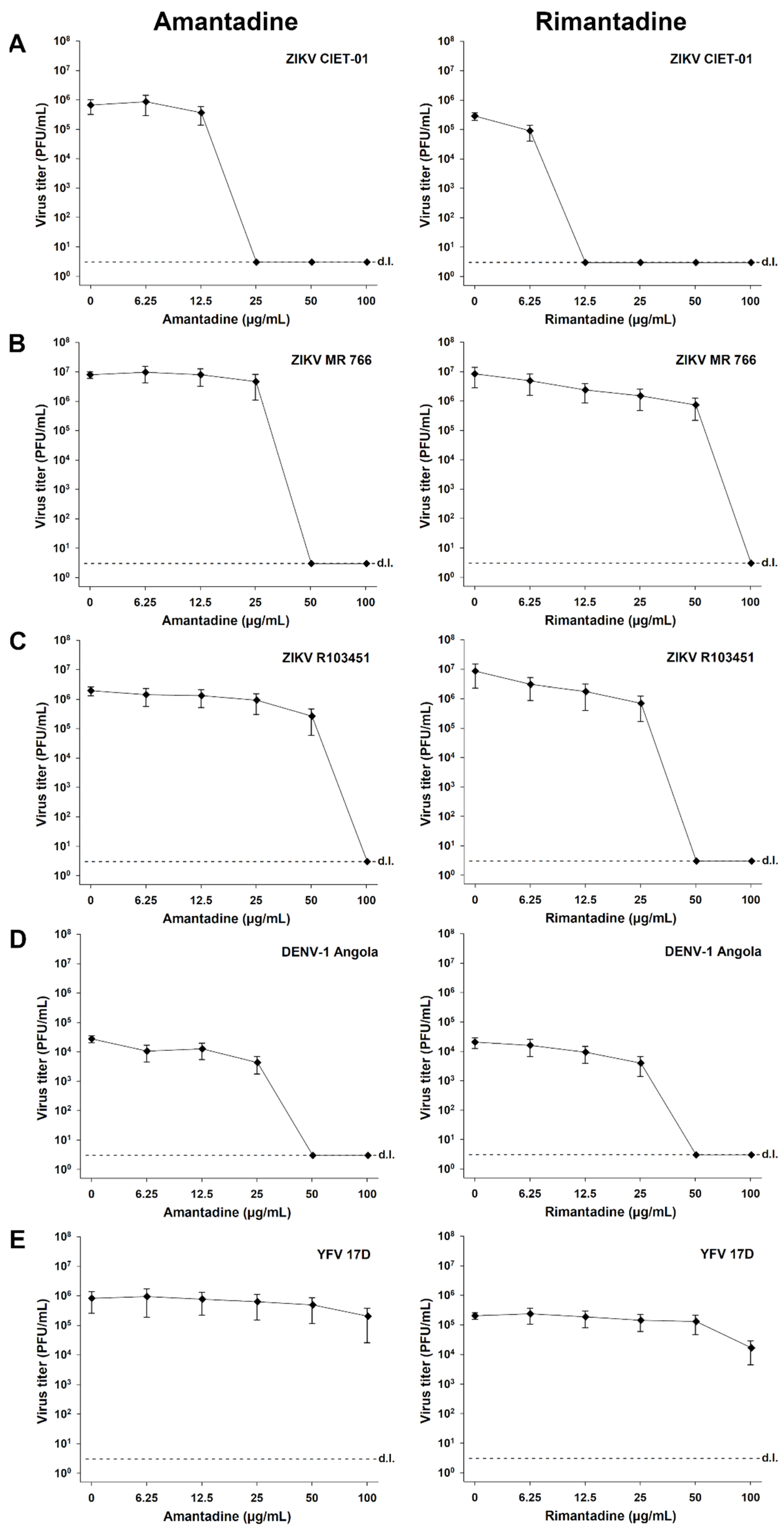
3.3. The Antiviral Activity of Adamantanes Is Also Observed for Other ZIKV Strains and DENV-1, though Only Slightly for YFV
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dick, G.W.A.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Boyer, S.; Calvez, E.; Chouin-Carneiro, T.; Diallo, D.; Failloux, A.B. An overview of mosquito vectors of Zika virus. Microbes Infect. 2018, 20, 646–660. [Google Scholar] [CrossRef]
- Li, Y.; He, L.; He, R.L.; Yau, S.S.T. Zika and flaviviruses phylogeny based on the alignment-free natural vector method. DNA Cell Biol. 2017, 36, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Macnamara, F.N. Zika virus: A report on three cases of human infection during an epidemic of jaundice in Nigeria. Trans. R. Soc. Trop. Med. Hyg. 1954, 48, 139–145. [Google Scholar] [CrossRef]
- Duffy, M.R.; Chen, T.H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; Dubray, C.; et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef] [PubMed]
- Musso, D.; Bossin, H.; Mallet, H.P.; Besnard, M.; Broult, J.; Baudouin, L.; Levi, J.E.; Sabino, E.C.; Ghawche, F.; Lanteri, M.C.; et al. Zika virus in French Polynesia 2013–14: Anatomy of a completed outbreak. Lancet Infect. Dis. 2018, 18, e172–e182. [Google Scholar] [CrossRef]
- Lowe, R.; Barcellos, C.; Brasil, P.; Cruz, O.G.; Honório, N.A.; Kuper, H.; Carvalho, M.S. The zika virus epidemic in brazil: From discovery to future implications. Int. J. Environ. Res. Public Health 2018, 15, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Styczynski, A.R.; Malta, J.M.A.S.; Krow-Lucal, E.R.; Percio, J.; Nóbrega, M.E.; Vargas, A.; Lanzieri, T.M.; Leite, P.L.; Staples, J.E.; Fischer, M.X.; et al. Increased rates of Guillain-Barré syndrome associated with Zika virus outbreak in the Salvador metropolitan area, Brazil. PLoS Negl. Trop. Dis. 2017, 11, e0005869. [Google Scholar] [CrossRef]
- Campos Coelho, A.V.; Crovella, S. Microcephaly prevalence in infants born to zika virus-infectedwomen: A systematic review and meta-analysis. Int. J. Mol. Sci. 2017, 18, 1714. [Google Scholar] [CrossRef] [Green Version]
- Pierson, T.C.; Diamond, M.S. The emergence of Zika virus and its new clinical syndromes. Nature 2018, 560, 573–581. [Google Scholar] [CrossRef]
- Sacramento, C.Q.; De Melo, G.R.; De Freitas, C.S.; Rocha, N.; Hoelz, L.V.B.; Miranda, M.; Fintelman-Rodrigues, N.; Marttorelli, A.; Ferreira, A.C.; Barbosa-Lima, G.; et al. The clinically approved antiviral drug sofosbuvir inhibits Zika virus replication. Sci. Rep. 2017, 7, 40920. [Google Scholar] [CrossRef]
- Gorshkov, K.; Shiryaev, S.A.; Fertel, S.; Lin, Y.W.; Huang, C.T.; Pinto, A.; Farhy, C.; Strongin, A.Y.; Zheng, W.; Terskikh, A.V. Zika virus: Origins, pathological action, and treatment strategies. Front. Microbiol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Yan, J.; Gao, G.F. Monoclonal antibodies against Zika virus: Therapeutics and their implications for vaccine design. J. Virol. 2017, 91, e01049-17. [Google Scholar] [CrossRef] [Green Version]
- Saiz, J.C.; Martín-Acebes, M.A. The race to find antivirals for zika virus. Antimicrob. Agents Chemother. 2017, 61, e00411-17. [Google Scholar] [CrossRef] [Green Version]
- Bleidner, W.E.; Harmon, J.B.; Hewes, W.E.; Lynes, T.E.; Hermann, E.C. Absorption, distribution and excretion of amantadine hydrochloride. J. Pharmacol. Exp. Ther. 1965, 150, 484–490. [Google Scholar] [PubMed]
- Hay, A.J.; Wolstenholme, A.J.; Skehel, J.J.; Smith, M.H. The molecular basis of the specific anti-influenza action of amantadine. EMBO J. 1985, 4, 3021–3024. [Google Scholar] [CrossRef] [PubMed]
- Wintermeyer, S.M.; Nahata, M.C. Rimantadine: A clinical perspective. Ann. Pharmacother. 1995, 29, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Drakopoulos, A.; Tzitzoglaki, C.; Ma, C.; Freudenberger, K.; Hoffmann, A.; Hu, Y.; Gauglitz, G.; Schmidtke, M.; Wang, J.; Kolocouris, A. Affinity of rimantadine enantiomers against influenza A/M2 protein revisited. ACS Med. Chem. Lett. 2017, 8, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.B.; Purcell, R.H.; Chanock, R.M. Effect of amantadine hydrochloride on parainfluenza type 1 virus infections in adult volunteers. Am. Rev. Respir. Dis. 1967, 95, 689–690. [Google Scholar] [CrossRef]
- Levin, M.J. Treatment and prevention options for respiratory syncytial virus infections. J. Pediatr. 1994, 124, S22–S27. [Google Scholar] [CrossRef]
- Kim, M.C.; Lee, N.P. How to treat influenza and colds. West. J. Med. 2000, 172, 260–264. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Chan, K.H.; Jiang, Y.; Kao, R.Y.T.; Lu, H.T.; Fan, K.W.; Cheng, V.C.C.; Tsui, W.H.W.; Hung, I.F.N.; Lee, T.S.W.; et al. In Vitro susceptibility of 10 clinical isolates of SARS coronavirus to selected antiviral compounds. J. Clin. Virol. 2004, 31, 69–75. [Google Scholar] [CrossRef]
- Tanner, J.A.; Zheng, B.J.; Zhou, J.; Watt, R.M.; Jiang, J.Q.; Wong, K.L.; Lin, Y.P.; Lu, L.Y.; He, M.L.; Kung, H.F.; et al. The adamantane-derived bananins are potent inhibitors of the helicase activities and replication of SARS coronavirus. Chem. Biol. 2005, 12, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimolai, N. Potentially repurposing adamantanes for COVID-19. J. Med. Virol. 2020, 92, 531–532. [Google Scholar] [CrossRef] [PubMed]
- Staničová, J.; Miškovský, P.; Šutiak, V. Amantadine: An antiviral and antiparkinsonian agent. Vet. Med. 2001, 46, 244–256. [Google Scholar] [CrossRef] [Green Version]
- Martín, J.; Navas, S.; Fernández, M.; Rico, M.; Pardo, M.; Quiroga, J.A.; Zahm, F.; Carreño, V. In vitro effect of amantadine and interferon α-2a on hepatitis C virus markers in cultured peripheral blood mononuclear cells from hepatitis C virus-infected patients. Antivir. Res. 1999, 42, 59–70. [Google Scholar] [CrossRef]
- Blázquez, A.B.; Martín-Acebes, M.A.; Saiz, J.C. Inhibition of West Nile virus multiplication in cell culture by anti-parkinsonian drugs. Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koff, W.C.; Elm, J.R.; Halstead, S.B. Inhibition of dengue virus replication by amantadine hydrochloride. Antimicrob. Agents Chemother. 1980, 18, 125–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koff, W.C.; Elm, J.R.; Halstead, S.B. Suppression of dengue vitus replication in vitro by rimantadine hydrochloride. Am. J. Trop. Med. Hyg. 1981, 30, 184–189. [Google Scholar] [CrossRef]
- Arias-Arias, J.L.; MacPherson, D.J.; Hill, M.E.; Hardy, J.A.; Mora-Rodríguez, R. A fluorescence-activatable reporter of flavivirus NS2B–NS3 protease activity enables live imaging of infection in single cells and viral plaques. J. Biol. Chem. 2020, 295, 2212–2226. [Google Scholar] [CrossRef] [PubMed]
- Arias-Arias, J.L.; Vega-Aguilar, F.; Corrales-Aguilar, E.; Hun, L.; Loría, G.D.; Mora-Rodríguez, R. Dengue virus infection of primary human smooth muscle cells. Am. J. Trop. Med. Hyg. 2018, 99, 1451–1457. [Google Scholar] [CrossRef] [Green Version]
- Medina, F.; Medina, J.F.; Colon, C.; Vergne, E.; Santiago, G.A.; Munoz-Jordan, J.L. Dengue virus: Isolation, propagation, quantification, and storage. In Current Protocols in Microbiology; Wiley: Hoboken, NJ, USA, 2012; pp. 15D.2.1–15D.2.24. ISBN 9780471729259. [Google Scholar]
- Morens, D.M.; Halstead, S.B.; Repik, P.M.; Putvatana, R.; Raybourne, N. Simplified plaque reduction neutralization assay for dengue viruses by semimicro methods in BHK-21 cells: Comparison of the BHK suspension test with standard plaque reduction neutralization. J. Clin. Microbiol. 1985, 22, 250–254. [Google Scholar] [CrossRef] [Green Version]
- Baz, M.; Boivin, G. Antiviral agents in development for zika virus infections. Pharmaceuticals 2019, 12, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendel, H.A.; Snyder, M.T.; Pell, S. Trial of amantadine in epidemic influenza. Clin. Pharmacol. Ther. 1965, 7, 38–43. [Google Scholar] [CrossRef]
- Dolin, R.; Reichman, R.C.; Madore, P.; Maynard, R.; Linton, P.N.; Webber-Jones, R.N. A controlled trial of amantadine and rimantadine in the prophylaxis of influenza A infection. N. Engl. J. Med. 1982, 307, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Khazaei, M.; Karevan, A.; Taheri, M.; Ghafouri-Fard, S. Comparison of the effects of amantadine and ondansetron in treatment of fatigue in patients with multiple sclerosis. Clin. Transl. Med. 2019, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Raupp-Barcaro, I.F.; Vital, M.A.; Galduróz, J.C.; Andreatini, R. Potential antidepressant effect of amantadine: A review of preclinical studies and clinical trials. Rev. Bras. Psiquiatr. 2018, 40, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Giacino, J.T.; Whyte, J.; Bagiella, E.; Kalmar, K.; Childs, N.; Khademi, A.; Eifert, B.; Long, D.; Katz, D.I.; Cho, S.; et al. Placebo-controlled trial of amantadine for severe traumatic brain injury. N. Engl. J. Med. 2012, 366, 819–826. [Google Scholar] [CrossRef] [Green Version]
- Sawada, H.; Oeda, T.; Kuno, S.; Nomoto, M.; Yamamoto, K.; Yamamoto, M.; Hisanaga, K.; Kawamura, T. Amantadine for dyskinesias in parkinson’s disease: A randomized controlled trial. PLoS ONE 2010, 5, e15298. [Google Scholar] [CrossRef]
- Maynard, M.; Pradat, P.; Bailly, F.; Rozier, F.; Nemoz, C.; Si Ahmed, S.N.; Adeleine, P.; Trépo, C. Amantadine triple therapy for non-responder hepatitis C patients. Clues for controversies (ANRS HC 03 BITRI). J. Hepatol. 2006, 44, 484–490. [Google Scholar] [CrossRef]
- Lin, C.C.; Chen, W.C. Treatment effectiveness of amantadine against dengue virus infection. Am. J. Case Rep. 2016, 17, 921–924. [Google Scholar] [CrossRef] [Green Version]
- Vernier, V.G.; Harmon, J.B.; Stump, J.M.; Lynes, T.E.; Marvel, J.P.; Smith, D.H. The toxicologic and pharmacologic properties of amantadine hydrochloride. Toxicol. Appl. Pharmacol. 1969, 15, 642–665. [Google Scholar] [CrossRef]
- Levy, M.; Pastuszak, A.; Koren, G. Fetal outcome following intrauterine amantadine exposure. Reprod. Toxicol. 1991, 5, 79–81. [Google Scholar] [CrossRef]
- Kirshon, B.; Faro, S.; Zurawin, R.K.; Samo, T.C.; Carpenter, R.J. Favorable outcome after treatment with amantadine and ribavirin in a pregnancy complicated by influenza pneumonia. A case report. J. Reprod. Med. Obstet. Gynecol. 1988, 33, 399–401. [Google Scholar]
- Tomar, P.P.S.; Oren, R.; Krugliak, M.; Arkin, I.T. Potential viroporin candidates from pathogenic viruses using bacteria-based bioassays. Viruses 2019, 11, 632. [Google Scholar] [CrossRef] [Green Version]
- Griffin, S.D.C. Plugging the holes in hepatitis C virus antiviral therapy. Proc. Natl. Acad. Sci. USA 2009, 106, 12567–12568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, J.; Maheswari, U.; Parthasarathy, K.; Ng, L.; Liu, D.X.; Gong, X. Conductance and amantadine binding of a pore formed by a lysine-flanked transmembrane domain of SARS coronavirus envelope protein. Protein Sci. 2007, 16, 2065–2071. [Google Scholar] [CrossRef]
- Verdiá-Báguena, C.; Nieto-Torres, J.L.; Alcaraz, A.; DeDiego, M.L.; Torres, J.; Aguilella, V.M.; Enjuanes, L. Coronavirus E protein forms ion channels with functionally and structurally-involved membrane lipids. Virology 2012, 432, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlton, F.W.; Pearson, H.M.; Hover, S.; Lippiat, J.D.; Fontana, J.; Barr, J.N.; Mankouri, J. Ion Channels as therapeutic targets for viral infections: Further discoveries and future perspectives. Viruses 2020, 12, 844. [Google Scholar] [CrossRef]
- Lamb, R.A.; Choppin, P.W. Identification of a second protein (M2) encoded by RNA segment 7 of influenza virus. Virology 1981, 112, 729–737. [Google Scholar] [CrossRef]
- Pinto, L.H.; Holsinger, L.J.; Lamb, R.A. Influenza virus M2 protein has ion channel activity. Cell 1992, 69, 517–528. [Google Scholar] [CrossRef]
- Mihm, U.; Grigorian, N.; Welsch, C.; Herrmann, E.; Kronenberger, B.; Teuber, G.; Von Wagner, M.; Hofmann, W.P.; Albrecht, M.; Lengauer, T.; et al. Amino acid variations in hepatitis C virus p7 and sensitivity to antiviral combination therapy with amantadine in chronic hepatitis C. Antivir. Ther. 2006, 11, 507–519. [Google Scholar] [PubMed]
- Foster, T.L.; Verow, M.; Wozniak, A.L.; Bentham, M.J.; Thompson, J.; Atkins, E.; Weinman, S.A.; Fishwick, C.; Foster, R.; Harris, M.; et al. Resistance mutations define specific antiviral effects for inhibitors of the hepatitis C virus p7 ion channel. Hepatology 2011, 54, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, A.; Horan, C.R.; Gage, P.W. Dengue virus M protein C-terminal peptide (DVM-C) forms ion channels. J. Membr. Biol. 2005, 204, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, G.; García-Cordero, J.; León-Juárez, M.; Oza, G.; Tapia-Ramírez, J.; Villegas-Sepulveda, N.; Cedillo-Barrón, L. NS2A comprises a putative viroporin of Dengue virus 2. Virulence 2017, 8, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- León-Juárez, M.; Martínez-Castillo, M.; Shrivastava, G.; García-Cordero, J.; Villegas-Sepulveda, N.; Mondragón-Castelán, M.; Mondragón-Flores, R.; Cedillo-Barrón, L. Recombinant Dengue virus protein NS2B alters membrane permeability in different membrane models. Virol. J. 2016, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.; Lefteri, D.; Singh, R.; Thompson, R.; Maskell, D.; Bentham, M.; Swinscoe, G.; Merits, A.; Donald, C.; Naransonleedsacuk, N.R.; et al. Inhibitors of the small membrane ( M ) protein viroporin prevent Zika virus infection. bioRxiv 2021, 1–49. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Amantadine (CC50 b > 100 µg/mL) | Rimantadine (CC50 > 100 µg/mL) | ||||
|---|---|---|---|---|---|---|
| IC50 c (µg/mL) | IC90 d (µg/mL) | SI e | IC50 (µg/mL) | IC90 (µg/mL) | SI | |
| ZIKV CIET-01 | 12.25 ± 2.28 | 16.04 ± 1.94 | >8.16 | 5.89 ± 0.22 | 6.96 ± 0.19 | >16.98 |
| ZIKV MR 766 | 26.52 ± 12.79 | 32.62 ± 9.26 | >3.77 | 8.95 ± 3.79 | 36.04 ± 11.19 | >11.17 |
| ZIKV R103451 | 29.05 ± 13.79 | 61.48 ± 19.00 | >3.44 | 4.91 ± 2.38 | 19.60 ± 4.87 | >20.37 |
| DENV-1 Angola | 12.81 ± 3.56 | 35.11 ± 5.06 | >7.81 | 12.26 ± 1.67 | 37.14 ± 5.54 | >8.17 |
| YFV 17D | 50.62 ± 8.51 | >100 | >1.98 | 74.69 ± 13.09 | >100 | >1.34 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arias-Arias, J.L.; Vega-Aguilar, F.; Picado-Soto, D.; Corrales-Aguilar, E.; Loría, G.D. In Vitro Inhibition of Zika Virus Replication with Amantadine and Rimantadine Hydrochlorides. Microbiol. Res. 2021, 12, 727-738. https://doi.org/10.3390/microbiolres12030052
Arias-Arias JL, Vega-Aguilar F, Picado-Soto D, Corrales-Aguilar E, Loría GD. In Vitro Inhibition of Zika Virus Replication with Amantadine and Rimantadine Hydrochlorides. Microbiology Research. 2021; 12(3):727-738. https://doi.org/10.3390/microbiolres12030052
Chicago/Turabian StyleArias-Arias, Jorge L., Francisco Vega-Aguilar, Dihalá Picado-Soto, Eugenia Corrales-Aguilar, and Gilbert D. Loría. 2021. "In Vitro Inhibition of Zika Virus Replication with Amantadine and Rimantadine Hydrochlorides" Microbiology Research 12, no. 3: 727-738. https://doi.org/10.3390/microbiolres12030052
APA StyleArias-Arias, J. L., Vega-Aguilar, F., Picado-Soto, D., Corrales-Aguilar, E., & Loría, G. D. (2021). In Vitro Inhibition of Zika Virus Replication with Amantadine and Rimantadine Hydrochlorides. Microbiology Research, 12(3), 727-738. https://doi.org/10.3390/microbiolres12030052