
Intramuscular Boosting with hIFN-Alpha 2b Enhances BCGphipps-Induced Protection in a Murine Model of Leprosy

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Microorganisms
2.3. Immunizations
2.4. M. lepraemurium Challenge
2.5. Enzyme-Linked Immunosorbent Assays (ELISA)
2.6. Measurements of NO in the Sera of Mice
2.7. Immunofluorescence
2.8. Statistical Analysis
3. Results
3.1. BCGphipps (BCGph)-Priming Followed by Successive Intramuscular hIFN-α 2b Boosting Protects Balb/c Mice from Intradermal Challenge with M. lepraemurium
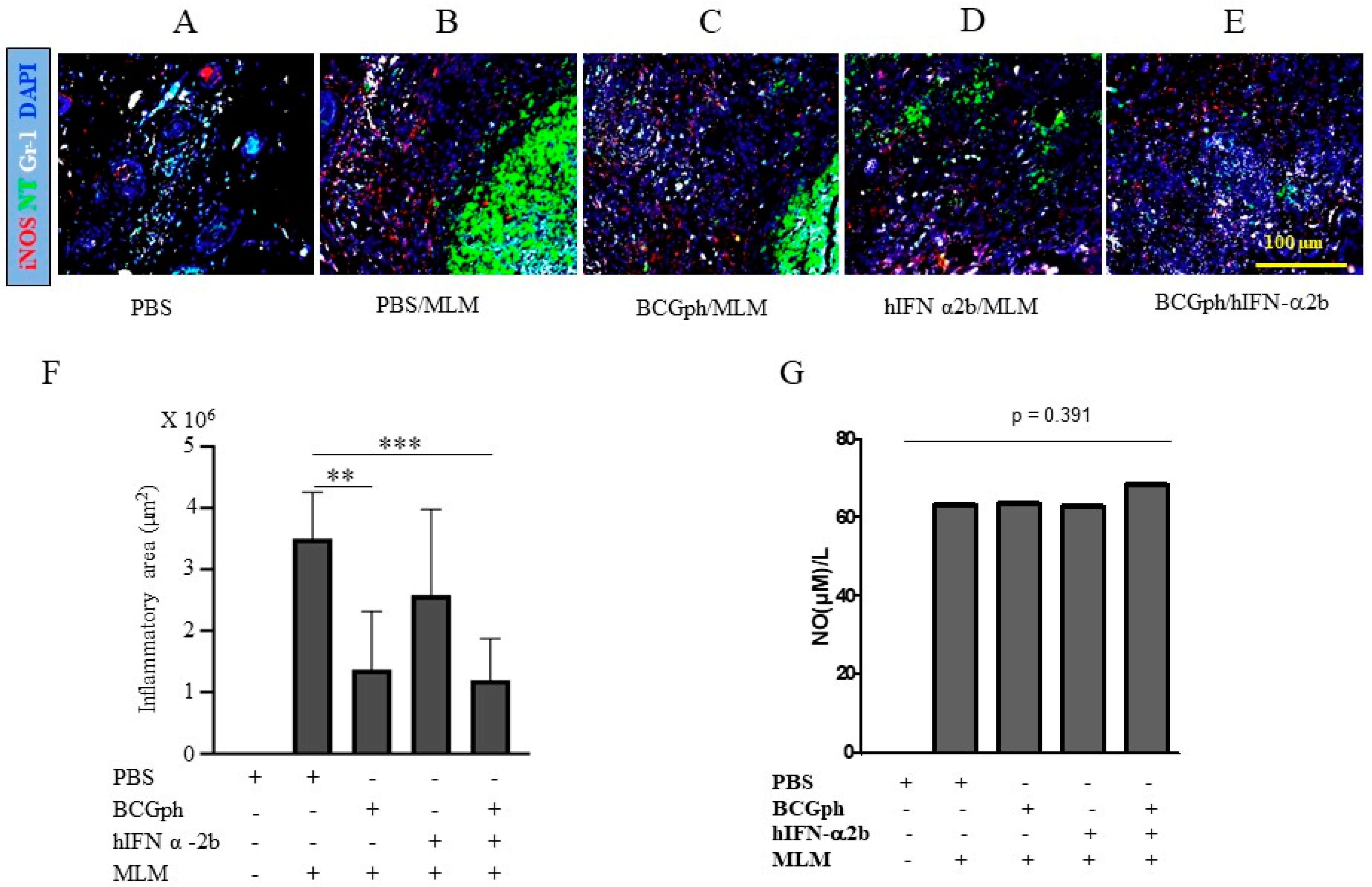
3.2. Minimal Skin Pathology in Mice Primed with BCGphipps and Boosted with hIFNα 2b Correlates with Decreased Nitrotyrosine Accumulation and Impaired Neutrophil Infiltration
3.3. Anti-Inflammatory Cytokines Are Increased in the Serum of Mice Primed with BCGphipps and Boosted with hIFN-α 2b
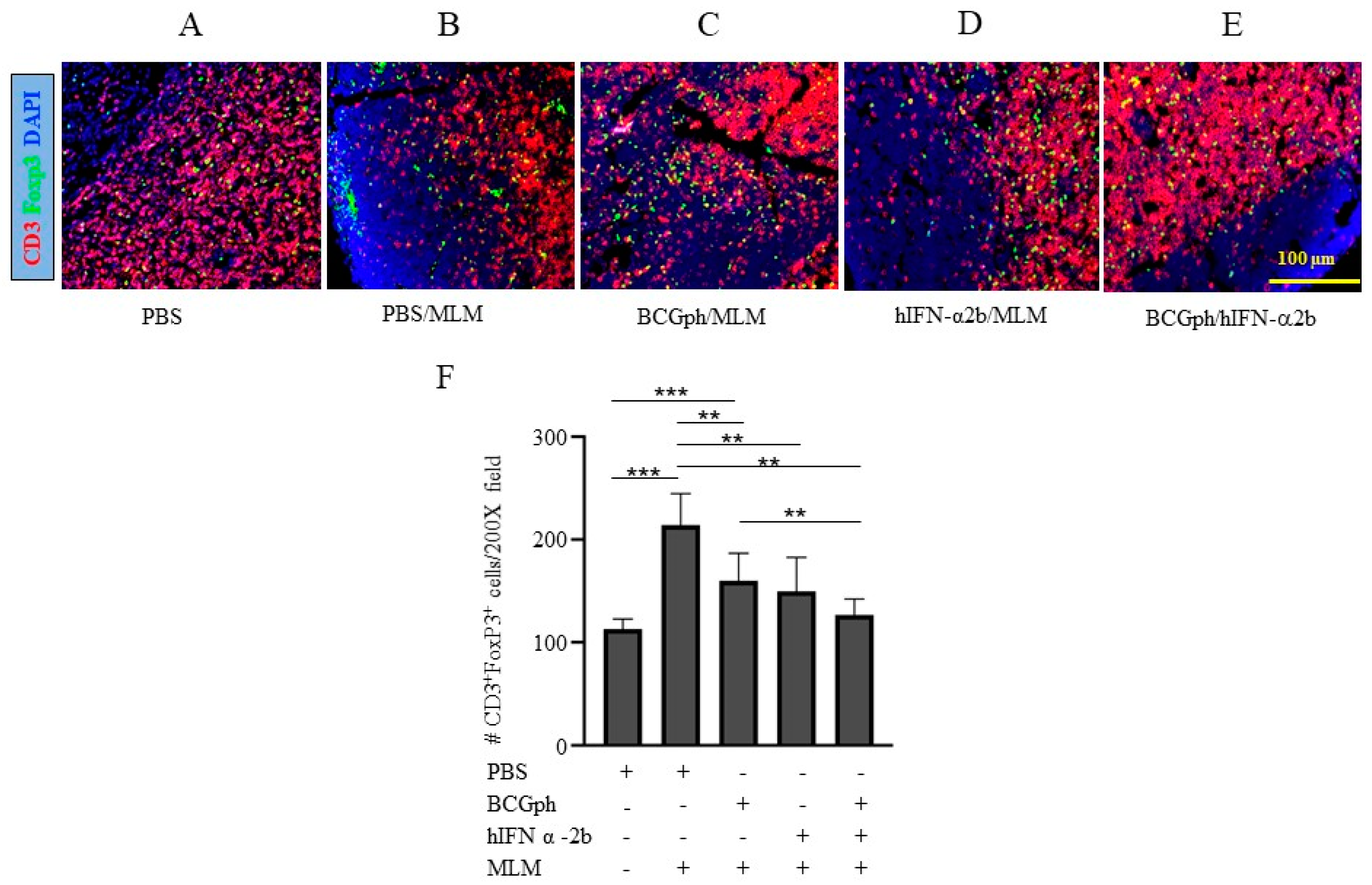
3.4. Impaired Treg Accumulation in the Draining Lymph Nodes (DLNs) of Mice Primed with BCGphipps or Boosted with hIFN-α 2b
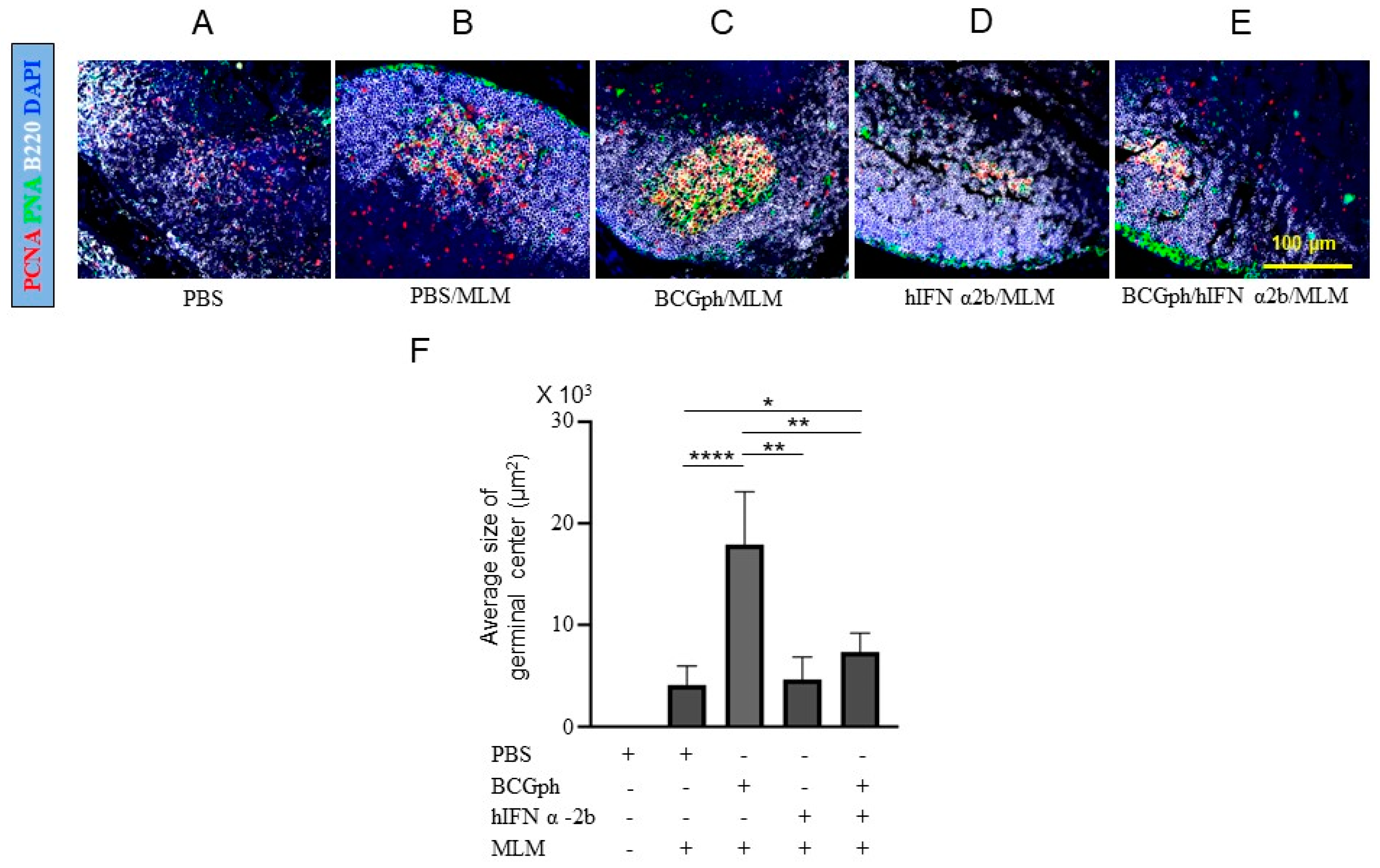
3.5. BCGphipps Priming and hIFN-α 2b Boosting Influences GCs Responses in Mouse DLNs
3.6. Comparable Levels of Glycolipid-Specific Antibodies in Mice Challenged with M. lepraemurium after BCGphipps Priming and hIFN-α 2b Boosting
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Health Organization: Tuberculosis—Global Facts 2011/2102. Geneva: WHO Stop TB Dep. 2012. Available online: http://www.who.int/tb/publications/2011/factsheet_tb_2011.pdf (accessed on 1 February 2021).
- Fine, P.E. Leprosy: What is being “eliminated”? Bull. World Health Organ. 2007, 85, 1–2. [Google Scholar] [CrossRef]
- Rambukkana, A. How does Mycobacterium leprae target the peripheral nervous system? Trends Microbiol. 2000, 8, 23–28. [Google Scholar] [CrossRef]
- Rodriguez, J.I.A.; Triellato, G.L.; Meirelles, N.M.L.; Teixeira, C.C.; Lyon, S.; Esteves, A.R.M. Leprosy classification methods: A comparative study in a referral character in Brazil. Int. J. Infect. Dis. 2016, 45, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Geluk, A.; van der Ploeg, J.; Teles, O.B.R.; Kees, L.; Franken, M.C.; Prins, C.; Wouter, D.J.; Sarno, N.E.; Sampaio, E.P.; Otenhoff, T.H.M. Rational combination of peptides derived from different Mycobacterium leprae proteins improves sensitivity for immunodiagnosis of M. leprae infection. Clin. Vacc. Immunol. 2008, 15, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Geluk, A.; Bobosha, K.; Schip, J.J.V.D.P.-V.; Spencer, J.S.; Banu, S.; Martins, M.V.S.B.; Cho, S.N.; Franken, K.L.M.C.; Kim, H.J.; Bekele, Y.; et al. New Biomarkers with Relevance to Leprosy Diagnosis Applicable in Areas Hyperendemic for Leprosy. J. Immunol. 2012, 188, 4782–4791. [Google Scholar] [CrossRef]
- Geluk, A. Biomarkers for Leprosy: Would you prefer T (cells)? Lepr. Rev. 2013, 84, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Alcaïs, A.; Alter, A.; Antoni, G.; Orlova, M.; Nguyen, V.T.; Singh, M.; Vanderborght, P.R.; Katoch, K.; Mira, M.T.; Vu, H.T.; et al. Stepwise replication identifies a low-producing lymphotoxin-α allele as a major risk factor for early-onset leprosy. Nat. Genet. 2007, 39, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Avanzi, C.; Singh, P.; Truman, R.W.; Suffys, P.N. Molecular epidemiology of leprosy: An update. Infect. Genet. Evol. 2020, 86, 104581. [Google Scholar] [CrossRef]
- Lastoria, J.C.; Abreu, M.A. Leprosy review of the epidemiological clinical and ethnopathogenic aspects-part 1. Bras. Dermatol. 2014, 89, 205–218. [Google Scholar] [CrossRef]
- Rojas-Espinosa, O. Current Topics on the Profiles of Host Immunological Response to Mycobacterial Infections; Review Book; Tomioka, H., Ed.; Research Signpost: Trivandrum, India, 2009; pp. 97–140. [Google Scholar]
- Adams, L.B.; Pena, T.M.; Sharma, R.; Hagge, A.D.; Schurr, E.; Truman, R.W. Insights from animal models on the immunogenetics of leprosy. A Review. Mem. Inst. Oswaldo. Cruz. 2012, 107, 197–208. [Google Scholar] [CrossRef]
- Rojas-Espinosa, O.; Becerril-Villanueva, E.; Wek-Rodriguez, K.; Arce-Paredes, P.; Reyes-Maldonado, E. Palsy of the rear limbs in Mycobacterium lepraemurium-infected mice results from bone damage and not from nerve involvement. Clin. Exp. Immunol. 2005, 140, 436–442. [Google Scholar] [CrossRef]
- Modlin, L.R. The innate immune response in leprosy. Curr. Opin. Immunol. 2010, 22, 48–54. [Google Scholar] [CrossRef]
- Oliveira, R.B.; Ochoa, M.T.; Sieling, P.A.; Rea, T.H.; Rambukkana, A.; Sarno, E.N.; Modlin, R.L. Expression of Toll-Like Receptor 2 on Human Schwann Cells: A Mechanism of Nerve Damage in Leprosy. Infect. Immun. 2003, 71, 1427–1433. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Evans, T.; Buttery, L.; Carpenter, A.; Springall, D.R.; Polak, J.; Cohen, J. Cytokine-Treated Human Neutrophils Contain Inducible Nitric Oxide Synthase That Produces Nitration of Ingested Bacteria. Proc. Natl. Acad. Sci. USA 1996, 93, 9553–9554. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, A.B.; Simon, M.D.; Cazzaniga, R.A.; de Moura, T.R.; de Almeida, R.P.; Duthie, M.S.; Reed, S.G.; de Jesus, A.R. The influence of innate and adaptative immune responses on the differential clinical outcomes of leprosy. Infect. Dis. Poverty 2017, 6, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Sadhu, S.; Khaitan, B.K.; Joshi, B.; Sengupta, U.; Nautiyal, A.K.; Mitra, D.K. Reciprocity between Regulatory T Cells and Th17 Cells: Relevance to Polarized Immunity in Leprosy. PLoS Negl. Trop. Dis. 2016, 10, e0004338. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Espinosa, O.; Wek-Rodríguez, K.; Arce-Paredes, P. The effect of exogenous peroxidase on the evolution of murine leprosy. Int. J. Lepr. Other Mycobact. Dis. 2002, 70, 191–200. [Google Scholar]
- Ischiropoulos, H. Biological Tyrosine Nitration: A Pathophysiological Function of Nitric Oxide and Reactive Oxygen Species. Arch. Biochem. Biophys. 1998, 356, 1–11. [Google Scholar] [CrossRef]
- Young, D.B. Leprosy lipid provides the key to Schwann cell entry. Trends Microbiol. 2001, 9, 52–54. [Google Scholar] [CrossRef]
- Bannard, O.; Cyster, J.G. Germinal centers: Programmed for affinity maturation and antibody diversification. Curr. Opin. Immunol. 2017, 45, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Victora, D.G.; Nussenweig, M.C. Germinal centers. Annu. Rev. Immunol. 2012, 30, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, K.M.; Crotty, S.H. Germinal center enhancement by extended antigen availability. Curr. Opin. Immunol. 2017, 47, 64–69. [Google Scholar] [CrossRef]
- Good-Jacobson, K.L.; Szumilas, C.G.; Chen, L.; Sharpe, A.H.; Tomayko, M.M.; Shlomchik, M.J. PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells. Nat. Immunol. 2010, 11, 535–542. [Google Scholar] [CrossRef]
- Wollenberg, I.; Agua-Doce, A.; Hernández, A.; Almeida, C.; Oliveira, V.G.; Faro, J.; Graca, L. Regulation of the Germinal Center Reaction by Foxp3+ Follicular Regulatory T Cells. J. Immunol. 2011, 187, 4553–4560. [Google Scholar] [CrossRef] [PubMed]
- Litterman, M.A.; Piersone, W.; Lee, S.K.; Kallies, A.; Kawamot, S.; Rayner, T.F.; Srivastava, M.; Divekar, D.P.; Beaton, L.; Hogan, J.J.; et al. FOXP3+ follicular regulatory T cells control the germinal center response. Nat. Med. 2011, 17, 975–982. [Google Scholar] [CrossRef]
- Miles, B.; Connick, E. Control of the Germinal Center by Follicular Regulatory T Cells During Infection. Front. Immunol. 2018, 9, 2704–3710. [Google Scholar] [CrossRef]
- Fu, W.; Liu, X.; Lin, X.; Feng, H.; Sun, L.; Li, S.; Chen, H.; Tang, H.; Lu, L.; Jin, W.; et al. Deficiency in T follicular regulatory cells promotes autoimmunity. J. Exp. Med. 2018, 215, 815–825. [Google Scholar] [CrossRef]
- Pitzalis, C.; Jones, G.W.; Bombardieri, M.; Jones, S.A. Ectopic lymphoid-like structures in infection, cancer and autoimmunity. Nat. Rev. Immunol. 2014, 14, 447–462. [Google Scholar] [CrossRef]
- Aloulou, M.; Carr, E.J.; Gador, M.; Bignon, A.; Liblau, R.S.; Fazilleau, N.; Linterman, M.A. Follicular regulatory T cells can be specific for the immunizing antigen and derive from naive T cells. Nat. Commun. 2016, 7, 10579–10589. [Google Scholar] [CrossRef]
- Vinuesa, C.G.; Sanz, I.; Cook, M.C. Dysregulation of germinal centers in autoimmune disease. Nat. Rev. Immunol. 2009, 9, 845–857. [Google Scholar] [CrossRef]
- Cucak, H.; Yrlid, U.; Reizis, B.; Kalinke, U.; Johansson-Lindborn, B. Type I Interferon signaling in Dendritic cells stimulates the development of Lymph Node-Resident T follicular helper cells. Immunity 2009, 31, 491–501. [Google Scholar] [CrossRef]
- Desvignes, L.; Wolf, A.J.; Ernst, J.D. Dynamic Roles of Type I and Type II IFNs in Early Infection with Mycobacterium tuberculosis. J. Immunol. 2012, 188, 6205–6215. [Google Scholar] [CrossRef]
- Teles, M.B.R.; Graeber, G.T.; Krutzik, R.S.; Montoya, D.; Schenk, M.; Lee, J.D.; Komisopoulou, E.; Kelly-Scumpia, K.; Chun, R.; Iyer, S.S.; et al. Type I Interferon suppresses Type II Interferon-Triggered human Anti-Mycobacterial Responses. Science 2013, 339, 1448–1453. [Google Scholar] [CrossRef]
- Ji, D.X.; Yamashiro, L.H.; Chen, K.J.; Mukaida, N.; Kramnik, I.; Darwin, K.H.; Vance, R.E. Type I interferon-driven susceptibility to Mycobacterium tuberculosis is mediated by IL-1Ra. Nat. Microbiol. 2019, 4, 2128–2135. [Google Scholar] [CrossRef]
- Manca, C.; Tsenova, L.; Bergtold, A.; Freeman, S.H.; Tovey, M.; Musser, M.J.; Barry, E.C., 3rd; Freedman, H.V.; Kaplan, G. Virulence of a Mycobacterium tuberculosis clinical isolate in mice is determined by failure to induce Th1 type immunity and is associated with induction of IFN-α/β. Proc. Natl. Acad. Sci. USA 2007, 98, 5752–5757. [Google Scholar] [CrossRef] [PubMed]
- Telesca, C.; Angelico, M.; Piccolo, P.; Nosotti, L.; Morrone, A.; Longhi, C.; Carbone, M.; Baiocchi, L. Interferon-alpha treatment of hepatitis D induces tuberculosis exacerbation in an immigrant. J. Infect. 2007, 54, e223–e226. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, E.; Remoli, M.E.; Gafa, V.; Pardini, M.; Fattorini, L.; Coccia, E.M. IFN-β improves BCG immunogenicity by acting on DC maturation. J. Leukoc. Biol. 2008, 85, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santiago, C.E.; Guerrero, G.G. IFN-α Boosting of Mycobacterium bovis Bacillus Calmette Güerin-Vaccine Promoted Th1 Type Cellular Response and Protection against M. tuberculosis Infection. BioMed Res. Int. 2017, 2017, 6760. [Google Scholar] [CrossRef]
- Guerrero, G.G.; Rangel-Moreno, J.; Islas-Trujillo, S.; Rojas-Espinosa, O. Successive Intramuscular Boosting with IFN-Alpha Protects Mycobacterium bovis BCG-Vaccinated Mice against M. lepraemurium infection. BioMed Med. Res. Int. 2015, 2015, 414027. [Google Scholar]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferon in infectious disease. Nat. Rev. Immunol. 2015, 15, 67–103. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Visconti, R. Type I IFNs and regulation of TH1 responses: Enigmas both resolved and emerged. Nat. Immunol. 2000, 1, 17–19. [Google Scholar] [CrossRef]
- González-Navajas, J.M.; Lee, J.; David, M.; Raz, E. Immunomodulatory functions of type I interferons. Nat. Rev. Immunol. 2012, 12, 125–135. [Google Scholar] [CrossRef]
- Le Bon, A.; Tough, D.F. Type I interferon as a stimulus for cross-priming. Cytokine Growth Fact Rev. 2008, 19, 33–40. [Google Scholar] [CrossRef]
- Bracci, L.; Canini, I.; Puzelli, S.; Sestili, P.; Venditti, M.; Spada, M.; Donatelli, I.; Belardelli, F.; Proietti, E. Type I IFN is a powerful mucosal adjuvant for a selective intranasal vaccination against influenza virus in mice and affects antigen capture at mucosal level. Vaccine 2005, 23, 2994–3004. [Google Scholar] [CrossRef]
- Couch, R.B.; Atmar, R.L.; Cate, T.R.; Quarles, J.M.; Keitel, W.A.; Arden, N.H.; Wells, J.; Niño, D.; Wyde, P.R. Contrasting effects of type I interferon as a mucosal adjuvant for influenza vaccine in mice and humans. Vaccine 2009, 27, 5344–5348. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Denton, A.E.; Innocentin, S.; Carr, E.J.; Bradford, B.M.; Lafouresse, F.; Mabbott, N.A.; Mörbe, U.; Ludewig, B.; Groom, J.R.; Good-Jacobson, K.L.; et al. Type I interferon induces CXCL13 to support ectopic germinal center formation. J. Exp. Med. 2019, 216, 621–637. [Google Scholar] [CrossRef]
- Toledo, P.T.G.; Batista-Silva, L.R.; Medeiros, R.C.A.; Lara, F.A.; Moraes, M.O. Type I Interferons, Autophagy and Host Metabolism in Leprosy. Front. Immunol. 2018, 23, 806–817. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Y.; Zheng, H.; Pan, Y.; Liu, H.; Du, P.; Wan, L.; Liu, J.; Zhu, B.; Zhag, G.; et al. Genome sequencing and analysis of BCG vaccine strains. PLoS ONE 2013, 8, e071243. [Google Scholar] [CrossRef]
- Wek-Rodriguez, K.; Silva-Miranda, M.; Arce-Paredes, P.; Rojas-Espinosa, O. Effect of reactive oxygen intermediaries on the viability and infectivity of Mycobacterium lepraemurium. Int. J. Exp. Pathol. 2007, 88, 137–145. [Google Scholar] [CrossRef]
- Kim, S.H.; Cohen, B.; Novick, D.; Rubinstein, M. Mammalian Type I Interferon Receptors Consists of Two Subunits: IFN-αR1 and IFN-αR2. Gene 1997, 196, 279–286. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, X.; Han, R.A.; O’Donnell, M. Recombinant bacille Calmette-Guérin (BCG) expressing human interferon-alpha 2B demonstrates enhanced immunogenicity. Clin. Exp. Immunol. 2001, 123, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Nian, X.; Liu, C.; Fan, X.; Han, R. Construction of recombinant human IFNα-2b BCG and its antitumor effects on bladder cancer cells in vitro. Genet. Mol. Res. 2015, 14, 3436–3449. [Google Scholar] [CrossRef] [PubMed]
- Prabhakaran, K.; Harris, E.B.; Kirchheimer, W.F. Binding of 14C-labeled dopa by Mycobacterium leprae in vitro. Int. J. Lepr. Other Mycobact. Dis. 1976, 44, 58–64. [Google Scholar]
- Draper, P. Purification of Mycobacterium leprae. In Proceedings of the Annex 4 of Report of the Fifth Meeting of the Scientific Working Group on the Immunology of Leprosy (IMMLEP), Geneva, Switzerland, 24–26 June 1980; World Health Organization: Geneva, Switzerland, 1980. [Google Scholar]
- Ischiropoulos, H.; Nelson, J.; Duran, D.; Al-Mehdi, A. Reactions of nitric oxide and peroxynitrite with organic molecules and ferrihorseradish peroxidase: Interference with the determination of hydrogen peroxide. Free Radic. Biol. Med. 1996, 20, 373–381. [Google Scholar] [CrossRef]
- Schön, T.; Hernandez-Pando, R.; Negesse, Y.; Leekassa, R.; Sundqvist, T.; Britton, S. Expression of inducible nitric oxide synthase and nitrotyrosine in borderline leprosy lesions. Br. J. Dermatol. 2001, 145, 809–815. [Google Scholar] [CrossRef]
- Silva-Miranda, M.; WeK-Rodriguez, K.; Martinez, C.E.; Rojas-Espinosa, O. Expression of cyclooxygenase-2, alpha 1-acid-glycoprotein and inducible nitric oxide synthase in the developing lesions of murine leprosy. Int. J. Exp. Path. 2006, 87, 485–494. [Google Scholar] [CrossRef]
- Hernández-Pando, R.; Schön, T.; Orozco, E.H.; Serafín, T.; Estrada-Garcia, I. Expression of inducible nitric oxide synthase and nitrotyrosine during the evolution of experimental pulmonary tuberculosis. Exp. Toxic. Pathol. 2001, 53, 257–265. [Google Scholar] [CrossRef]
- Izcue, A.; Coombes, J.L.; Powrie, F. Regulatory Lymphocytes and Intestinal Inflammation. Annu. Rev. Immunol. 2009, 27, 313–338. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Miyara, M.; Costantino, C.M.; Hafler, D.A. FOXP3+ regulatory T cells in the human immune system. Nat. Rev. Immunol. 2010, 10, 490–500. [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of Protection Induced by Vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, Y.; Liu, H.; Xu, L.-L.; Teuscher, P.; Wang, S.; Lu, S.; Dent, A.L. Follicular regulatory T cells repress cytokine production by follicular helper T cells and optimize IgG responses in mice. Eur. J. Immunol. 2016, 46, 1152–1161. [Google Scholar] [CrossRef] [PubMed]
- Jeyanathan, M.; Damjanovic, D.; Shaler, C.R.; Lai, R.; Wortzman, M.; Yin, C.; Zganiacz, A.; Lichty, D.; Xing, Z. Differentially imprinted innate immunity by mucosal boost vaccination determines antituberculosis immune protective outcomes, independent of T-cell immunity. Mucosal Immunol. 2013, 6, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Jeyanathan, M.; Shao, Z.; Yu, X.; Harkness, R.; Jiang, R.; Li, J.; Xing, Z.; Zhu, T. AdHu5Ag85A Respiratory Mucosal Boost Immunization Enhances Protection against Pulmonary Tuberculosis in BCG-Primed Non-Human Primates. PLoS ONE 2015, 10, e0135009. [Google Scholar] [CrossRef]
- Afhami, S.; Drumond, V.A.; D’Agositnio, M.R.; Jeyanathan, M.; Gillgrass, A.; Xing, Z. Advancing Immunotherapeutic Vaccine Strategies Against Pulmonary Tuberculosis. Front. Immunol. 2020, 11, 557809. [Google Scholar] [CrossRef]
- Torrado, E.; Robinson, R.T.; Cooper, A.M. Cellular response to mycobacteria: Balancing protection and pathology. Trends Immunol. 2011, 32, 66–72. [Google Scholar] [CrossRef]
- Xu, X.; Gao, W.; Cheng, S.; Yin, D.; Li, F.; Wu, Y.; Sun, D.; Zhou, S.; Wang, D.; Zhang, Y.; et al. Anti-inflammatory and immunomodulatory mechanisms of atorvastatin in a murine model of traumatic brain injury. J. Neuroinflamm. 2017, 14, 167–182. [Google Scholar] [CrossRef]
- Azeem, W.; Maukon, B.R.; Appel, S.; Maegrete, A.O.; Kalland, K.H. Dual Pro- and Anti-Inflammatory Features of Monocyte-Derived Dendritic Cells. Front. Immunol. 2020, 11, 438–453. [Google Scholar] [CrossRef]
- Sultana, S.; Bishay, B. Neutralization of TNFR-1 and TNFR-2 modulates S. aureus induced septic arthritis by regulating the levels of pro-inflammatory and anti-inflammatory cytokines during the progression of the disease. Immunol. Lett. 2018, 196, 33–51. [Google Scholar] [CrossRef]
- Panigraphy, D.; Gilligan, M.M.; Huang, S.; Gartung, A.; Cortes-Puch, I.; Sime, P.J.; Phipps, R.P.; Serhan, C.H.N.; Hammock, B.D. Inflammation resolution: A dual-pronged approach to averting cytokine storms in COVID-19? Cancer Metastasis Rev. 2020, 39, 337–340. [Google Scholar] [CrossRef]
- Longhi, M.P.; Trumpfheller, C.; Idoyaga, J.; Caskey, M.; Matos, I.; Kluger, C.; Salazar, A.M.; Colonna, M.; Steinman, R.M. Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant. J. Exp. Med. 2009, 206, 1589–1602. [Google Scholar] [CrossRef]
- Khader, S.A.; Guglani, L.; Rangel-Moreno, J.; Gopal, R.; Junecko, B.A.F.; Fountain, J.J.; Martino, C.; Pearl, J.E.; Tighe, M.; Lin, Y.-Y.; et al. IL-23 Is Required for Long-Term Control of Mycobacterium tuberculosis and B Cell Follicle Formation in the Infected Lung. J. Immunol. 2011, 187, 5402–5407. [Google Scholar] [CrossRef] [PubMed]
- Choreño-Parra, J.A.; Bobba, S.; Rangel-Moreno, J.; Ahmed, M.; Mehra, S.; Rosa, B.; Martin, J.; Mitreva, M.; Kaushal, D.; Zúñiga, J.; et al. Mycobacterium tuberculosis HN878 Infection Induces Human-Like B-Cell Follicles in Mice. J. Infect. Dis. 2020, 221, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.G.; Haribhai, D.; Williams, J.B.; Aggarwal, P.; Jia, S.; Charbonnier, L.M.; Yan, K.; Lorier, R.; Turner, A.; Ziegelbauer, J.; et al. IL-10 produced by iTreg cells controls colitis and pathogenic ex. iTreg cells during immunotherapy. J. Immunol. 2012, 189, 5628–5648. [Google Scholar] [CrossRef]
- Rubtsovo, Y.P.; Rasmussen, J.P.; Chi, Y.; Fontenot, J.; Castelli, I.; Ye, X.; Treuting, P.; Siewe, I.; Roers, A.; Henderson, W.R.J.; et al. Regulatory T cell-derived interleukin-10 limits inflammation at environmental interfaces. Immunity 2018, 28, 546–558. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | IFN-γ | IL-4 | IL-17 | TNF-γ | IL-6 | IL-10 |
|---|---|---|---|---|---|---|
| PBS BCGph hIFN-α 2b S BCGph/hIFN-α 2b S | 625 ± 442 1641 ± 154 2086 ± 609 2062 ± 299 | 1250 ± 173 1437 ± 108 1637 ± 53 1875 ± 352 * | 95 ± 67 367 ± 55 1230 ± 0 421 ± 191 | 697 ± 436 784 ± 62 978 ± 257 859 ± 122 | 68 ± 48 1065 ± 60 1320 ± 322 1270 ± 26 | 1763 ± 124 3201 ± 214 2860 ± 338 4250 ± 271 * |
| Experimental Groups | IgM | IgG | IgG1 | IgG2a | IgG2b | IgG3 |
|---|---|---|---|---|---|---|
| PBS | 0.86 ± 0.30 | 1.05 ± 0.07 | 1.70 ± 0.0 | 1.25 ± 0.21 | 1.35 ± 0.07 | 0.89 ± 0.03 |
| BCGph | 0.99 ± 0.15 | 1.10 ± 0.33 | 1.10 ± 0.41 | 1.40 ± 0.32 | 1.30 ± 0.30 | 0.90 ± 0.28 |
| hIFN-α 2b S | 1.00 ± 0.27 | 1.40 ± 0.13 | 1.80 ± 0.11 | 1.56 ± 0.38 | 1.30 ± 0.08 | 1.10 ± 0.10 |
| BCGph-hIFN-α 2b S | 0.90 ± 0.07 | 1.05 ± 0.28 | 1.50 ± 0.21 | 1.40 ± 0.06 | 1.30 ± 0.19 | 0.93 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero, G.G.; Rangel-Moreno, J.; Islas-Trujillo, S.O.; Rojas-Espinosa, O. Intramuscular Boosting with hIFN-Alpha 2b Enhances BCGphipps-Induced Protection in a Murine Model of Leprosy. Microbiol. Res. 2021, 12, 711-726. https://doi.org/10.3390/microbiolres12030051
Guerrero GG, Rangel-Moreno J, Islas-Trujillo SO, Rojas-Espinosa O. Intramuscular Boosting with hIFN-Alpha 2b Enhances BCGphipps-Induced Protection in a Murine Model of Leprosy. Microbiology Research. 2021; 12(3):711-726. https://doi.org/10.3390/microbiolres12030051
Chicago/Turabian StyleGuerrero, Gloria G., Javier Rangel-Moreno, Sergio O. Islas-Trujillo, and Oscar Rojas-Espinosa. 2021. "Intramuscular Boosting with hIFN-Alpha 2b Enhances BCGphipps-Induced Protection in a Murine Model of Leprosy" Microbiology Research 12, no. 3: 711-726. https://doi.org/10.3390/microbiolres12030051
APA StyleGuerrero, G. G., Rangel-Moreno, J., Islas-Trujillo, S. O., & Rojas-Espinosa, O. (2021). Intramuscular Boosting with hIFN-Alpha 2b Enhances BCGphipps-Induced Protection in a Murine Model of Leprosy. Microbiology Research, 12(3), 711-726. https://doi.org/10.3390/microbiolres12030051