
New Strains of Wolbachia Unveiling the Complexity of This Symbiotic Interaction in Solenopsis (Hymenoptera: Formicidae)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ant Samples and Wolbachia Isolation
2.2. DNA Sequencing
2.3. Sequence Editing, Allele Identification and Sequence Typing
2.4. Phylogenetic Reconstruction and Mantel Test
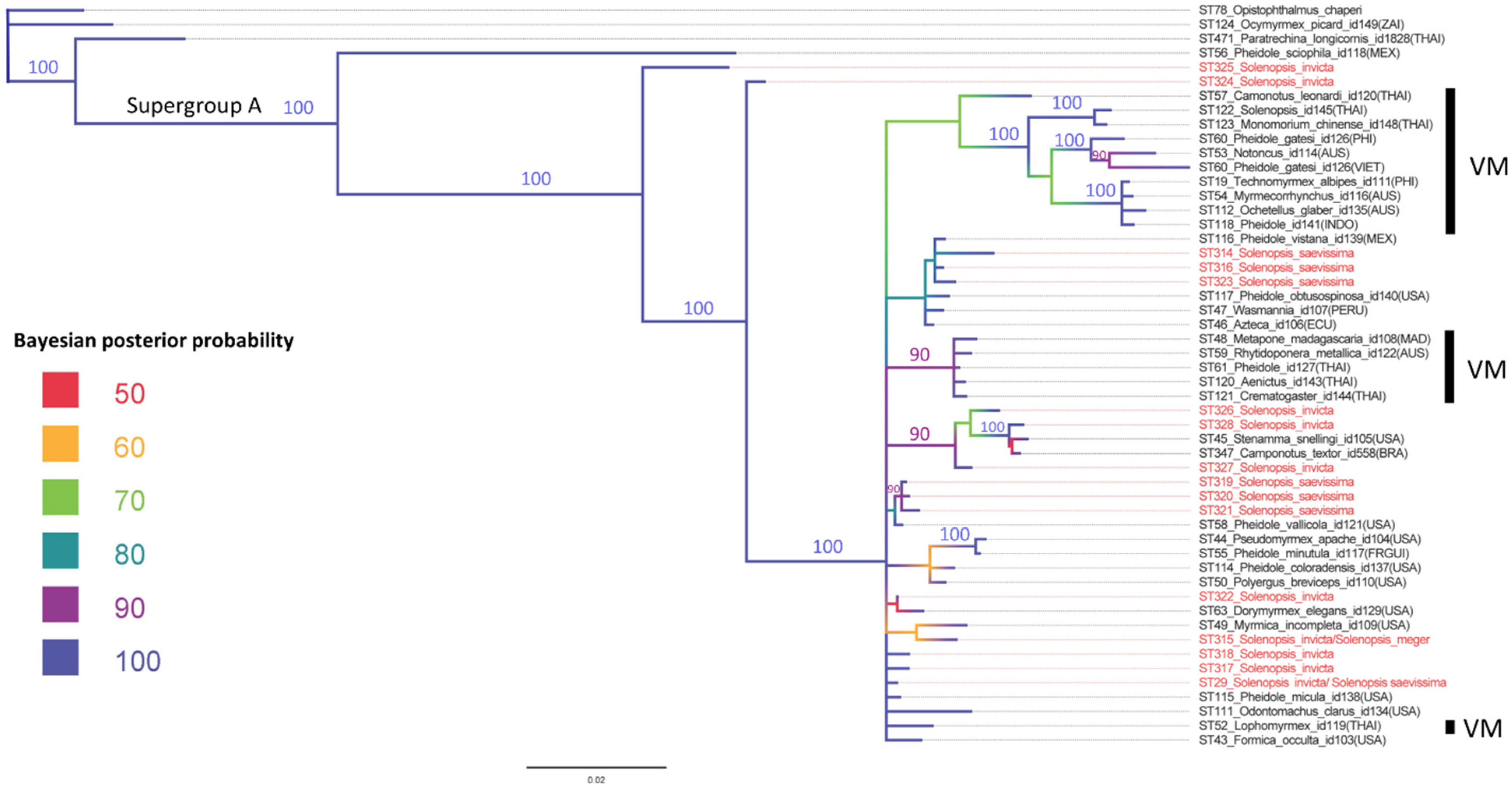
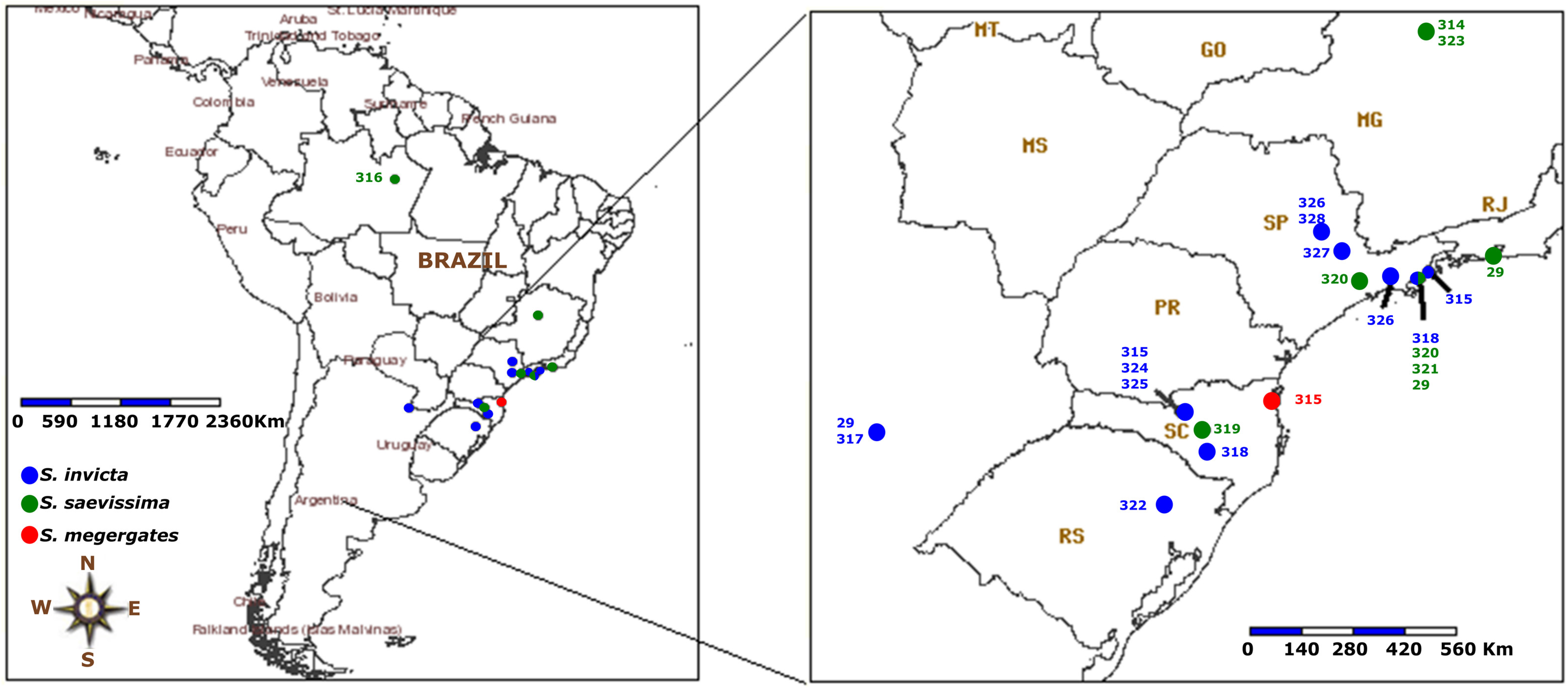
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef]
- Russell, J.A. The ants (Hymenoptera: Formicidae) are unique and enigmatic hosts of prevalent Wolbachia (Alphaproteobacteria) symbionts. Myrmecol. News 2012, 16, 7–23. [Google Scholar]
- Russell, J.A.; Funaro, C.F.; Giraldo, Y.M.; Goldman-Huertas, B.; Suh, D.; Kronauer, D.J.C.; Moreau, C.S.; Pierce, N.E. A veritable menagerie of heritable bacteria from ants, butterflies, and beyond: Broad molecular surveys and a systematic review. PLoS ONE 2012, 7, e51027. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Chen, S.; Huang, Y.; Pierce, N.E.; Riegler, M.; Yang, F.; Zeng, L.; Lu, Y.; Liang, G.; Xu, Y. Symbiotic microbiota may reflect host adaptation by resident to invasive ant species. PLoS Pathog. 2019, 15, e1007942. [Google Scholar] [CrossRef]
- Singh, R.; Linksvayer, T.A. Wolbachia-infected ant colonies have increased reproductive investment and an accelerated life cycle. J. Exp. Biol. 2020, 223, jeb220079. [Google Scholar] [CrossRef]
- Kautz, S.; Rubin, B.E.R.; Moreau, C.S. Bacterial infections across the ants: Frequency and prevalence of Wolbachia, Spiroplasma, and Asaia. Psyche (Stuttg.) 2013. [Google Scholar] [CrossRef]
- Ramalho, M.O.; Kim, Z.; Wang, S.; Moreau, C.S. Wolbachia across social insects: Patterns and implications. Ann. Entomol. Soc. Am. 2021. [Google Scholar] [CrossRef]
- Baldo, L.; Lo, N.; Werren, J.H. Mosaic nature of the Wolbachia surface protein. J. Bacteriol. 2005, 187, 5406–5418. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Dunning Hotopp, J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef]
- Frost, C.L.; FernÁndez-MarÍn, H.; Smith, J.E.; Hughes, W.O.H. Multiple gains and losses of Wolbachia symbionts across a tribe of fungus-growing ants. Mol. Ecol. 2010, 19, 4077–4085. [Google Scholar] [CrossRef]
- Kelly, M.; Price, S.L.; Ramalho, M.O.; Moreau, C.S. Diversity of Wolbachia associated with the giant turtle ant, Cephalotes atratus. Curr. Microbiol. 2019, 76, 1330–1337. [Google Scholar] [CrossRef]
- Ramalho, M.O.; Martins, C.; Silva, L.M.R.; Martins, V.G.; Bueno, O.C. Intracellular symbiotic bacteria of Camponotus textor, Forel (Hymenoptera, Formicidae). Curr. Microbiol. 2017, 74, 589–597. [Google Scholar] [CrossRef]
- Reeves, D.D.; Price, S.L.; Ramalho, M.O.; Moreau, C.S. The diversity and distribution of Wolbachia, Rhizobiales, and Ophiocordyceps within the widespread neotropical turtle ant, Cephalotes atratus (Hymenoptera: Formicidae). Neotrop. Entomol. 2020, 49, 52–60. [Google Scholar] [CrossRef]
- Russell, J.A.; Goldman-Huertas, B.; Moreau, C.S.; Baldo, L.; Stahlhut, J.K.; Werren, J.H.; Pierce, N.E. Specialization and geographic isolation among Wolbachia symbionts from ants and lycaenid butterflies. Evolution (N. Y.) 2009, 63, 624–640. [Google Scholar] [CrossRef]
- Bolton, B. An Online Catalog of the Ants of the World. Available online: http://antcat.org (accessed on 3 April 2021).
- Pitts, J.P.; Camacho, G.P.; Gotzek, D.; Mchugh, J.V.; Ross, K.G. Revision of the fire ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae). Proc. Entomol. Soc. Wash. 2018, 120, 308–411. [Google Scholar] [CrossRef]
- Ahrens, M.E.; Ross, K.G.; Shoemaker, D.D. Phylogenographic structure of the fire ant Solenopsis invicta in its native South American range: Roles of natural barriers and habitat connectivity. Evolution (N. Y.) 2005, 59, 1733–1743. [Google Scholar] [CrossRef]
- Dedeine, F.; Ahrens, M.; Calcaterra, L.; Shoemaker, D.D. Social parasitism in fire ants (Solenopsis spp.): A potential mechanism for interspecies transfer of Wolbachia. Mol. Ecol. 2005, 14, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.F.; Ramalho, J.D.S.; Morini, M.S.C.; Wolff, J.L.C.; Araújo, R.C.; Mascara, D. Identification and characterization of Wolbachia in Solenopsis saevissima fire ants (Hymenoptera: Formicidae) in southeastern Brazil. Curr. Microbiol. 2009, 58, 189–194. [Google Scholar] [CrossRef]
- Martins, C.; Souza, R.F.; Bueno, O.C. Presence and distribution of the endosymbiont Wolbachia among Solenopsis spp. (Hymenoptera: Formicidae) from Brazil and its evolutionary history. J. Invertebr. Pathol. 2012, 109, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, D.D.; Ahrens, M.; Sheill, L.; Mescher, M.; Keller, L.; Ross, K.G. Distribution and prevalence of Wolbachia infections in native populations of the fire ant Solenopsis invicta (Hymenoptera: Formicidae). Environ. Entomol. 2003, 32, 1329–1336. [Google Scholar] [CrossRef]
- Shoemaker, D.D.; Ross, K.G.; Keller, L.; Vargo, E.L.; Werren, J.H. Wolbachia infections in native and introduced populations of fire ants (Solenopsis spp.). Insect Mol. Biol. 2000, 9, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.F.; Martins, C.; Pereira, R.; Bueno, O.C. Analysis of the hypervariable regions (HVRs) of the wsp gene of Wolbachia from Solenopsis invicta ants in southeastern Brazil. Adv. Entomol. 2014, 2, 135–143. [Google Scholar] [CrossRef][Green Version]
- Bleidorn, C.; Gerth, M. A critical re-evaluation of multilocus sequence typing (MLST) efforts in Wolbachia. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, T.M.; Bruzzese, D.J.; Klasson, L.; Corretto, E.; Lečić, S.; Stauffer, C.; Feder, J.L.; Schuler, H. Comparative genome sequencing reveals insights into the dynamics of Wolbachia in native and invasive cherry fruit flies. Mol. Ecol. 2021. [Google Scholar] [CrossRef]
- Scholz, M.; Albanese, D.; Tuohy, K.; Donati, C.; Segata, N.; Rota-Stabelli, O. Large scale genome reconstructions illuminate Wolbachia evolution. Nat. Commun. 2020, 11, 5235. [Google Scholar] [CrossRef] [PubMed]
- Dhaygude, K.; Nair, A.; Johansson, H.; Wurm, Y.; Sundström, L. The first draft genomes of the ant Formica exsecta, and its Wolbachia endosymbiont reveal extensive gene transfer from endosymbiont to host. BMC Genom. 2019, 20, 301. [Google Scholar] [CrossRef] [PubMed]
- Braig, H.R.; Zhou, W.; Dobson, S.L.; O’Neill, S.L. Cloning and characterization of a gene encoding the major surface protein of the bacterial endosymbiont Wolbachia pipientis. J. Bacteriol. 1998, 180, 2373–2378. [Google Scholar] [CrossRef]
- Pitts, J.P.; McHugh, J.V.; Ross, K.G. Cladistic analysis of the fire ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae). Zool. Scr. 2005, 34, 493–505. [Google Scholar] [CrossRef]
- Trager, J.C. A Revision of the fire ants, Solenopsis geminata group (Hymenoptera: Formicidae: Myrmicinae). J. N. Y. Entomol. Soc. 1991, 99, 141–198. [Google Scholar]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Higgins, D.G.; Bleasby, A.J.; Fuchs, R. Clustal V: Improved software for multiple sequence alignment. Comput. Appl. Biosci. 1992, 8, 189–191. [Google Scholar] [CrossRef]
- Martins, C.; Souza, R.F.; Bueno, O.C. Molecular characterization of fire ants, Solenopsis spp., from Brazil based on analysis of mtDNA gene cytochrome oxidase I. J. Insect Sci. 2014, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Portals. Available online: http://www.phylo.org/ (accessed on 15 January 2020).
- Ramalho, M.O.; Moreau, C.S. The evolution and biogeography of Wolbachia in ants (Hymenoptera: Formicidae). Diversity 2020, 12, 426. [Google Scholar] [CrossRef]
- R Development Core Team R: A Language and Environment for Statistical Computing. 2017. Available online: https://www.R-project.org/ (accessed on 16 October 2017).
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Bivand, R.; Keitt, T.; Rowlingson, B. rgdal: Bindings for the Geospatial Data Abstraction Library. 2013. Available online: https://CRAN.R-project.org/package=rgdal (accessed on 16 October 2017).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. R Package Version 2.4-5 2017. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 16 October 2017).
- Species Mapper. Available online: http://splink.cria.org.br/mapper?criaLANG=en (accessed on 5 May 2017).
- Baldo, L.; Bordenstein, S.; Wernegreen, J.J.; Werren, J.H. Widespread recombination throughout Wolbachia genomes. Mol. Biol. Evol. 2006, 23, 437–449. [Google Scholar] [CrossRef]
- Reuter, M.; Keller, L. High levels of multiple Wolbachia infection and recombination in the ant Formica exsecta. Mol. Biol. Evol. 2003, 20, 748–753. [Google Scholar] [CrossRef][Green Version]
- Van Borm, S.; Wenseleers, T.; Billen, J.; Boomsma, J.J. Cloning and sequencing of wsp encoding gene fragments reveals a diversity of co-infecting Wolbachia strains in Acromyrmex leafcutter ants. Mol. Phylogenet. Evol. 2003, 26, 102–109. [Google Scholar] [CrossRef]
- Ahrens, M.E.; Shoemaker, D. Evolutionary history of Wolbachia infections in the fire ant Solenopsis invicta. BMC Evol. Biol. 2005, 5, 35. [Google Scholar] [CrossRef]
- Viljakainen, L.; Reuter, M.; Pamilo, P. Wolbachia transmission dynamics in Formica wood ants. BMC Evol. Biol. 2008, 8, 55. [Google Scholar] [CrossRef]
- Ramalho, M.O.; Bueno, O.C.; Moreau, C.S. Species-specific signatures of the microbiome from Camponotus and Colobopsis ants across developmental stages. PLoS ONE 2017, 12, e0187461. [Google Scholar] [CrossRef]
- Ramalho, M.O.; Bueno, O.C.; Moreau, C.S. Microbial composition of spiny ants (Hymenoptera: Formicidae: Polyrhachis) across their geographic range. BMC Evol. Biol. 2017. [Google Scholar] [CrossRef]
- Zhang, Y.-K.; Zhang, K.-J.; Sun, J.-T.; Yang, X.-M.; Ge, C.; Hong, X.-Y. Diversity of Wolbachia in natural populations of spider mites (genus Tetranychus): Evidence for complex infection history and disequilibrium distribution. Microb. Ecol. 2013, 65, 731–739. [Google Scholar] [CrossRef]
- Ramalho, M.O.; Martins, C.; Campos, T.; Nondillo, A.; Botton, M.; Bueno, O.C. Occurrence of 15 haplotypes of Linepithema micans (Hymenoptera: Formicidae) in southern Brazil. J. Econ. Entomol. 2017, 110, 1841–1846. [Google Scholar] [CrossRef]
- Yun, Y.; Lei, C.; Peng, Y.; Liu, F.; Chen, J.; Chen, L. Wolbachia strains typing in different geographic population spider, Hylyphantes graminicola (Linyphiidae). Curr. Microbiol. 2011, 62, 139–145. [Google Scholar] [CrossRef]
- Baldo, L.; Ayoub, N.A.; Hayashi, C.Y.; Russell, J.A.; Stahlhut, J.K.; Werren, J.H. Insight into the routes of Wolbachia invasion: High levels of horizontal transfer in the spider genus Agelenopsis revealed by Wolbachia strain and mitochondrial DNA diversity. Mol. Ecol. 2008, 17, 557–569. [Google Scholar] [CrossRef]
- Souza, R.F. Aspectos Bioecológicos e Genéticos de Solenopsis spp. No Mosaico de Paisagens do Alto Tietê Cabeceiras, São Paulo. Ph.D. Thesis, Universidade de Mogi das Cruzes, São Paulo, Brazil, 2019. [Google Scholar]
- Ramalho, M.O.; Vieira, A.S.; Pereira, M.C.; Moreau, C.S.; Bueno, O.C. Transovarian transmission of Blochmannia and Wolbachia endosymbionts in the neotropical weaver ant Camponotus textor (Hymenoptera, Formicidae). Curr. Microbiol. 2018, 75, 866–873. [Google Scholar] [CrossRef]
- Werren, J.H. Biology of Wolbachia. Annu. Rev. Entomol. 1997, 42, 587–609. [Google Scholar] [CrossRef]
- Schilthuizen, M.O.; Stouthamer, R. Horizontal transmission of parthenogenesis–inducing microbes in Trichogramma wasps. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1997, 264, 361–366. [Google Scholar] [CrossRef]
- Tolley, S.J.A.; Nonacs, P.; Sapountzis, P. Wolbachia horizontal transmission events in ants: What do we know and what can we learn? Front. Microbiol. 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Raychoudhury, R.; Baldo, L.; Oliveira, D.C.S.G.; Werren, J.H. Modes of acquisition of Wolbachia: Horizontal transfer, hybrid introgression, and codivergence in the Nasonia species complex. Evolution (N. Y.) 2009, 63, 165–183. [Google Scholar] [CrossRef]
- Watanabe, M.; Tagami, Y.; Miura, K.; Kageyama, D.; Stouthamer, R. Distribution patterns of Wolbachia endosymbionts in the closely related flower bugs of the genus Orius: Implications for coevolution and horizontal transfer. Microb. Ecol. 2012, 64, 537–545. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ant Species and Collection Code | Location | Geographic Coordinates | wsp | HVR1 | HVR2 | HVR3 | HVR4 | Id | ST | COI | gatB | coxA | hcpA | ftsZ | fbpA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. saevissima E1714 | Buritizeiro, MG, Brazil | S17°5′20″ W44°56′54″ | 28 | 21 | 21 | 25 | 21 | 516 | 314 * | JN808797 | 75 | 184 * | 45 | 37 | 46 |
| S. saevissima E1821 | Manaus, AM, Brazil | S03°06′25″ W60°01′34″ | 28 | 21 | 21 | 25 | 21 | 518 | 316 * | JN808830 | 75 | 20 | 45 | 17 | 252 * |
| S. saevissima E1792 | S. Cristovão do Sul, SC, Brazil | S27°15′32″ W50°26′50″ | p | 42 | 43 | 9 | 528 | 319 * | JN808815 | 19 | 20 | 207 * | 43 | 253 * | |
| S. saevissima E1738 | Rio de Janeiro, RJ, Brazil | S22°58′51″ W43°16′75″ | 28 | 21 | 21 | 25 | 21 | 533 | 29 | JN808818 | 19 | 20 | 22 | 17 | 20 |
| S. saevissima E1740 | Rio de Janeiro, RJ, Brazil | S22°58′51″ W43°16′75″ | 28 | 21 | 21 | 25 | 21 | 534 | 29 | JN808818 | 19 | 20 | 22 | 17 | 20 |
| S. saevissima E1742 | São Paulo, SP, Brazil | S23°32′53″ W46°38′11″ | 50 | 42 | 43 | 9 | 269 | 540 | 320 * | JN808781 | 19 | 20 | 207 * | 37 | 253 * |
| S. saevissima E1743 | Ubatuba, SP, Brazil | S23°30′21″ W45°07′55″ | 50 | 42 | 43 | 9 | 269 | 541 | 320 * | JN808781 | 19 | 20 | 207 * | 37 | 253 * |
| S. saevissima E1751 | Ubatuba, SP, Brazil | S23°30′21″ W45°07′55″ | p | 42 | 43 | 9 | 543 | 321 * | JN808807 | 19 | 20 | 208 * | 43 | 253 * | |
| S. saevissima E1713 | Buritizeiro, MG, Brazil | S17°25′20″ W44°56′54″ | 28 | 21 | 21 | 25 | 21 | 529 | 323 * | JN808797 | 75 | 20 | 45 | 17 | 46 |
| S. saevissima E1746 | Ubatuba, SP, Brazil | S23°30′21″ W45°07′55″ | 28 | 21 | 21 | 25 | 21 | 542 | 29 | JN808805 | 19 | 20 | 22 | 17 | 20 |
| S. saevissima E1753 | Ubatuba, SP, Brazil | S23°30′21″ W45°07′55″ | 28 | 21 | 21 | 25 | 21 | 544 | 29 | JN808808 | 19 | 20 | 22 | 17 | 20 |
| S. megergates E1782 | São Francisco, SC, Brazil | S26°33′53″ W48°43′10″ | p | 42 | 43 | 198 | 525 | 315 * | JN808826 | 196 * | 20 | 45 | 37 | 251 * | |
| S. megergates E1643 | Caçador, SC, Brazil | S26°46′32″ W51°00′56″ | 59 | 21 | 40 | 42 | 39 | 538 | 315 * | JN808826 | 196 * | 20 | 45 | 37 | 251 * |
| S. megergates E1644 | Caçador, SC, Brazil | S26°46′32″ W51°00′56″ | 59 | 21 | 40 | 42 | 39 | 539 | 315 * | JN808826 | 196 * | 20 | 45 | 37 | 251 * |
| S. invicta E1805 | Corrientes, Argentina | S27°18′39″ W58°33′44″ | 28 | 21 | 21 | 25 | 21 | 515 | 29 | JN808817 | 19 | 20 | 22 | 17 | 20 |
| S. invicta E1686 | Picinguaba, SP, Brazil | S23°19′02″ W44°54′04″ | 59 | 21 | 40 | 42 | 39 | 517 | 315 * | JN808784 | 196 * | 20 | 45 | 37 | 251 * |
| S. invicta E1801 | Corrientes, Argentina | S27°18′39″ W58°33′44″ | 28 | 21 | 21 | 25 | 21 | 519 | 29 | JN808817 | 19 | 20 | 22 | 17 | 20 |
| S. invicta E1802 | Corrientes, Argentina | S27°18′39″ W58°33′44″ | 28 | 21 | 21 | 25 | 21 | 520 | 29 | JN808817 | 19 | 20 | 22 | 17 | 20 |
| S. invicta E1803 | Corrientes, Argentina | S27°18′39″ W58°33′44″ | 28 | 21 | 21 | 25 | 21 | 521 | 29 | JN808817 | 19 | 20 | 22 | 17 | 20 |
| S. invicta E1807 | Corrientes, Argentina | S27°18′39″ W58°33′44″ | 28 | 21 | 21 | 25 | 21 | 522 | 29 | JN808817 | 19 | 20 | 22 | 17 | 20 |
| S. invicta E1808 | Corrientes, Argentina | S27°18′39″ W58°33′44″ | 28 | 21 | 21 | 25 | 21 | 523 | 317 * | JN808817 | 19 | 20 | 22 | 37 | 20 |
| S. invicta E1810 | Corrientes, Argentina | S27°18′39″ W58°33′44″ | 28 | 21 | 21 | 25 | 21 | 524 | 29 | JN808817 | 19 | 20 | 22 | 17 | 20 |
| S. invicta E1784 | Lages, SC, Brazil | S27°48′57″ W50°22′17″ | 505 | 42 | 43 | 198 | 269 | 526 | 318 * | JN808819 | 19 | 20 | 55 | 37 | 46 |
| S. invicta E1789 | Pinto Bandeira, RS, Brazil | S29°07′21″ W51°26′56″ | p | 42 | 43 | 198 | 527 | 322 * | JN808814 | 19 | 20 | 55 | 160* | 46 | |
| S. invicta E1749 | Ubatuba, SP, Brazil | S23°30′21″ W45°07′55″ | 505 | 42 | 43 | 198 | 269 | 537 | 318 * | JN808783 | 19 | 20 | 55 | 37 | 46 |
| S. invicta E1645_1 | Caçador, SC, Brazil | S26°46′32″ W51°00′56″ | 59 | 21 | 40 | 42 | 39 | 535 | 324 * | JN808837 | 196 * | 183 * | 45 | 17 | 251 * |
| S. invicta E1645_2 | Caçador, SC, Brazil | S26°46′32″ W51°00′56″ | 59 | 21 | 40 | 42 | 39 | 536 | 325 * | JN808837 | 196 * | 183 * | 45 | 17 | 254 * |
| S. invicta Sol128 | Campinas, SP, Brazil | S22°49′21″ W47°03′42″ | p | 42 | 43 | 198 | 546 | 327 * | KJ690243 | 19 | 20 | 210 * | 160 * | 46 | |
| S. invicta Sol106 | Rio Claro, SP, Brazil | S22°23′50″ W47°32′56″ | p | 42 | 43 | 198 | 547 | 328 * | KJ690242 | 3 | 20 | 210 * | 37 | 46 | |
| S. invicta Sol158 | Salesópolis, SP, Brazil | S23°32′00″ W45°50′55″ | p | 42 | 43 | 198 | 548 | 326 * | KJ690244 | 3 | 20 | 210 * | 160 * | 46 | |
| S. invicta Sol71 | Cachoeira de Minas, MG, Brazil | S22°23′49″ W45°50′55″ | p | 42 | 43 | 198 | 545 | 326 * | KJ690241 | 3 | 20 | 210 * | 160 * | 46 | |
| S. invicta E1725 | Porto Alegre, RS, Brazil | S29°59′14″ W51°09′580″ | 505 | 42 | 43 | 198 | 269 | 530 | na | JN808802 | 19 | 20 | 55 | 46 | |
| S. invicta E1726 | Porto Alegre, RS, Brazil | S29°59′14″ W51°09′580″ | 505 | 42 | 43 | 198 | 269 | 531 | na | JN808802 | 19 | 20 | 55 | 46 | |
| S. invicta E1727 | Porto Alegre, RS, Brazil | S29°59′14″ W51°09′580″ | 505 | 42 | 43 | 198 | 269 | 532 | na | JN808803 | 19 | 20 | 55 | 46 | |
| S. invicta Sol48 | Mogi das Cruzes, SP, Brazil | S23°25′19″ W46°05′24″ | p | 21 | 40 | 549 | na | KJ690243 | 3 | 210 * | 37 | ||||
| S. invicta E1739 | Rio de Janeiro, RJ, Brazil | S22°58′51″ W43°16′75″ | p | 21 | 40 | 42 | 552 | na | JN808826 | 19 | 20 | 45 | 251 * | ||
| S. invicta E1646_A | Caçador, SC, Brazil | S26°46′32″ W51°00′56″ | 59 | 21 | 40 | 42 | 39 | 553 | na | JN808838 | 20 | 45 | 37 | 251 * | |
| S. invicta E1646_B | Caçador, SC, Brazil | S26°46′32″ W51°00′56″ | 59 | 21 | 40 | 42 | 39 | 554 | na | JN808838 | 20 | 45 | 37 | 254 * | |
| S. geminata E1818 | Manaus, AM, Brazil | S03°06′25″ W60°01′34″ | 28 | 21 | 21 | 25 | 21 | 550 | na | JN808828 | 37 | ||||
| S. geminata E1822 | Manaus, AM, Brazil | S03°06′25″ W60°01′34″ | 28 | 21 | 21 | 25 | 21 | 551 | na | JN808832 | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, C.; Ramalho, M.d.O.; Silva, L.M.R.; Souza, R.F.d.; Bueno, O.C. New Strains of Wolbachia Unveiling the Complexity of This Symbiotic Interaction in Solenopsis (Hymenoptera: Formicidae). Microbiol. Res. 2021, 12, 567-579. https://doi.org/10.3390/microbiolres12030040
Martins C, Ramalho MdO, Silva LMR, Souza RFd, Bueno OC. New Strains of Wolbachia Unveiling the Complexity of This Symbiotic Interaction in Solenopsis (Hymenoptera: Formicidae). Microbiology Research. 2021; 12(3):567-579. https://doi.org/10.3390/microbiolres12030040
Chicago/Turabian StyleMartins, Cintia, Manuela de Oliveira Ramalho, Larissa Marin Rodrigues Silva, Rodrigo Fernando de Souza, and Odair Correa Bueno. 2021. "New Strains of Wolbachia Unveiling the Complexity of This Symbiotic Interaction in Solenopsis (Hymenoptera: Formicidae)" Microbiology Research 12, no. 3: 567-579. https://doi.org/10.3390/microbiolres12030040
APA StyleMartins, C., Ramalho, M. d. O., Silva, L. M. R., Souza, R. F. d., & Bueno, O. C. (2021). New Strains of Wolbachia Unveiling the Complexity of This Symbiotic Interaction in Solenopsis (Hymenoptera: Formicidae). Microbiology Research, 12(3), 567-579. https://doi.org/10.3390/microbiolres12030040