
Observations on the Potential of an Endophytic Fungus Associated with Cacao Leaves against Phytophthora palmivora


Abstract
:1. Introduction
2. Materials and Methods
2.1. Endophyte Isolation and Pathogenicity
2.2. Phytophthora Isolation and Pathogenicity
2.3. Cultural, Morphological and Molecular Characterization
2.4. Anti-Pathogen Assay
2.4.1. Dual Culture Test
2.4.2. Test for Antibiosis
2.4.3. Statistical Design and Data Analyses
3. Results
3.1. Pathogenicity Test
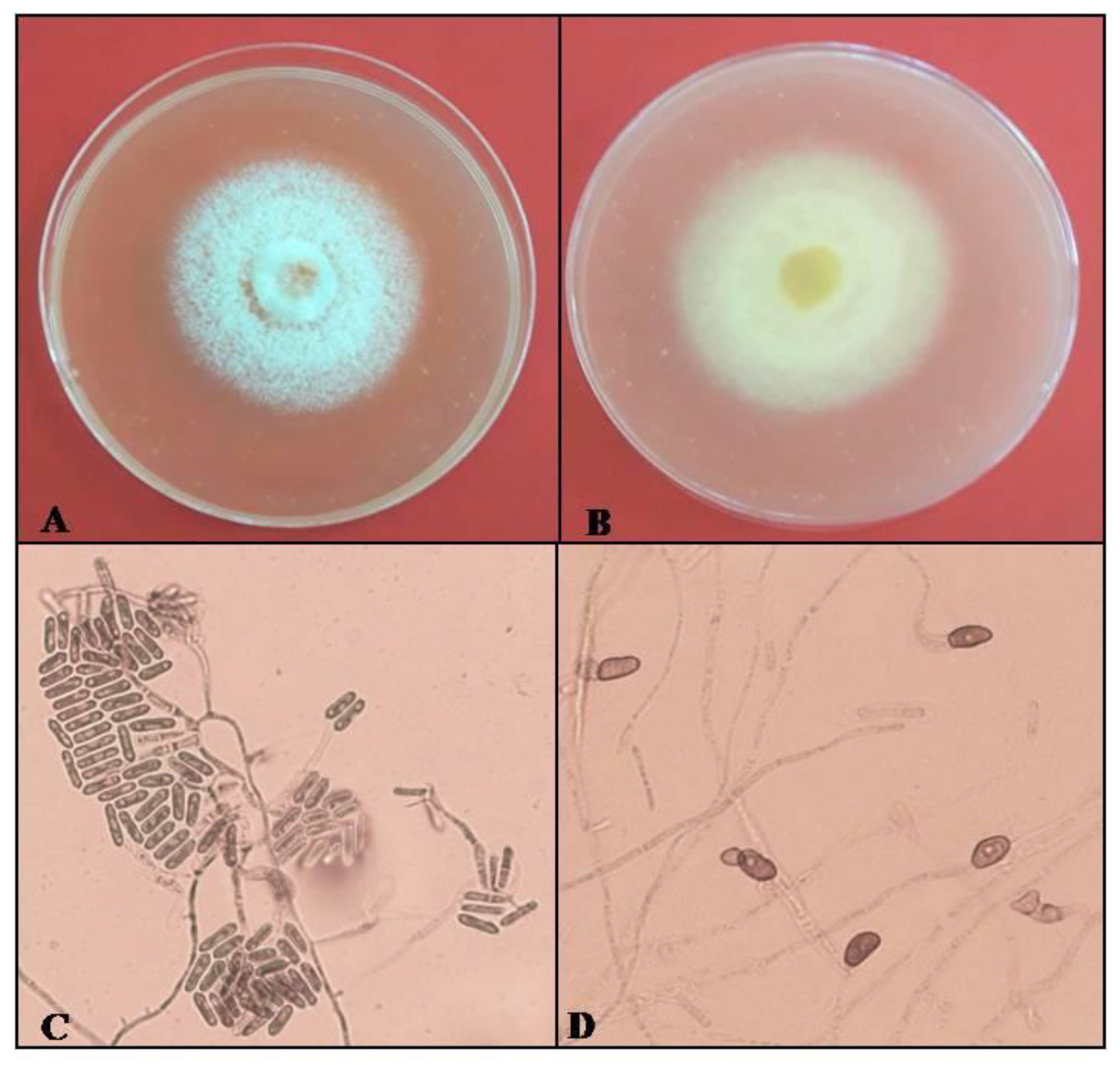
3.2. Cultural, Morphological and Molecular Identification
3.3. Anti-Pathogen Assay
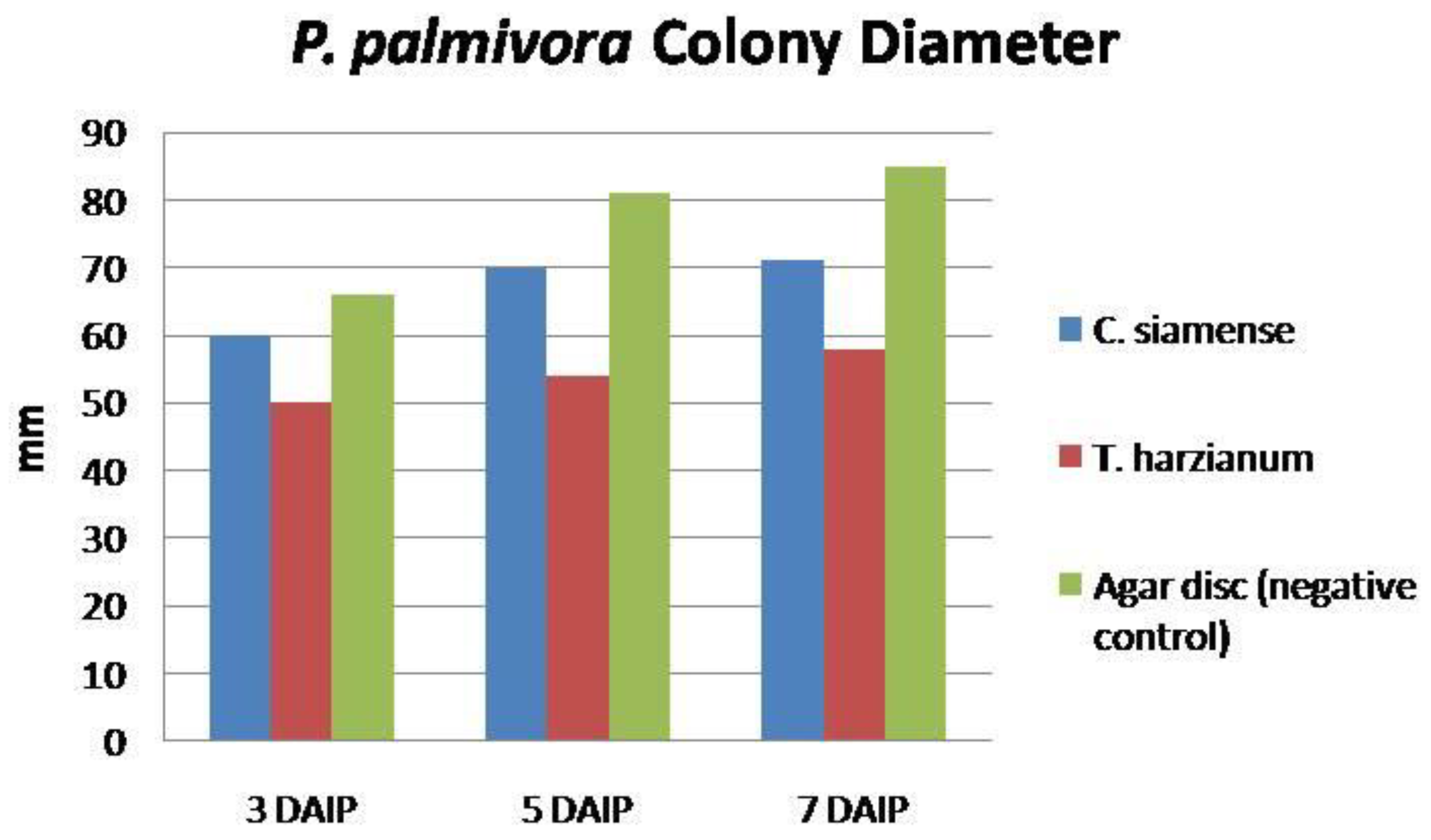
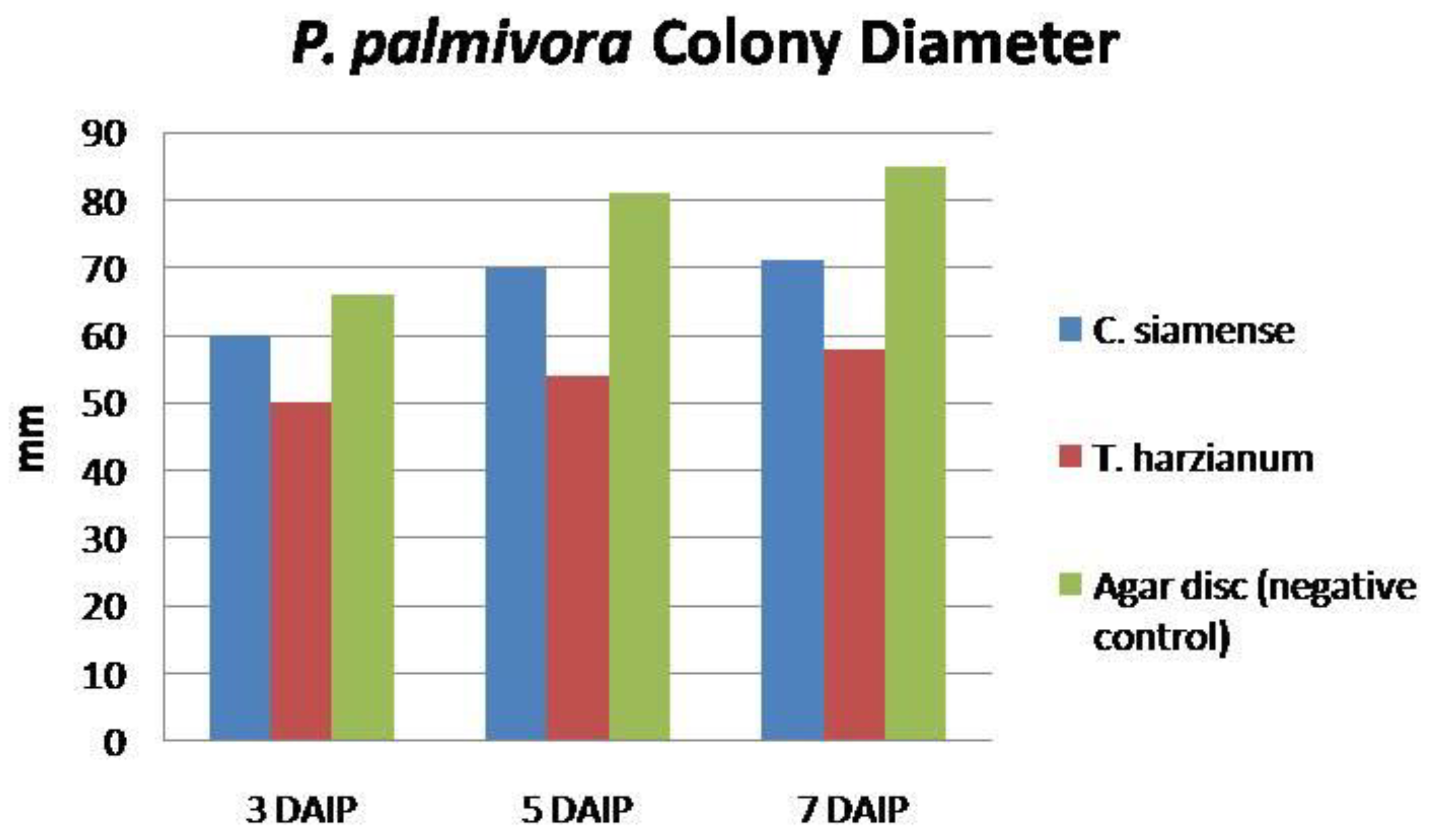
3.3.1. Competition
3.3.2. Antibiosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International Cocoa Organization (ICCO). Available online: https://www.icco.org (accessed on 25 January 2018).
- McMahon, P.; Purwantara, A. Phytophthora on Cocoa. In Diversity and Management of Phytophthora in Southeast Asia; Drenth, A., Guest, D.D., Eds.; ACIAR: Canberra, Australia, 2004; pp. 104–115. [Google Scholar]
- Guest, D. Black pod: Diverse pathogens with a global impact on cocoa yield. Phytopathology 2007, 97, 1650–1653. [Google Scholar] [CrossRef] [Green Version]
- Guest, D.; Anderson, R.D.; Foard, H.J.; Phillips, D.; Worboys, S.; Middleton, R.M. Long-term control of Phytophthora diseases of cocoa using trunk-injected phosphonate. Plant Pathol. 1994, 43, 479–492. [Google Scholar] [CrossRef]
- Drenth, A.; Guest, D. Phytophthora in the tropics. In Diversity and Management of Phytophthora in Southeast Asia; Drenth, A., Guest, D.A., Eds.; ACIAR: Canberra, Australia, 2004; pp. 30–41. [Google Scholar]
- Cohen, Y.; Coffey, M.D. Systemic fungicides and the control of Oomycetes. Ann. Rev. Phytopathol. 1986, 24, 311–338. [Google Scholar] [CrossRef]
- Kamil, M.J.A.; Fuddin, S.S.; Bong, C.L. Biological control of black pod disease on cocoa in Malaysia. In Diversity and Management of Phytophthora in Southeast Asia; Drenth, A., Guest, D.D., Eds.; ACIAR: Canberra, Australia, 2004; pp. 174–177. [Google Scholar]
- Pal, K.K.; McSpadden Gardener, B. Biological Control of Plant Pathogens. The Plant Health Instructor. Available online: https://www.apsnet.org (accessed on 20 February 2020).
- Petrini, O. Fungal endophytes of tree leaves. In Microbial Ecology of Leaves; Andrews, J.H., Hirano, S.S., Eds.; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- Christian, N.; Herre, E.A.; Mejia, L.C.; Clay, K. Exposure to the leaf litter microbiome of healthy adults protects seedlings from pathogen damage. Proc. R. Soc. B 2017, 20170641. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejia, L.C.; Kylo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [Green Version]
- Mejia, L.C.; Rojas, E.I.; Maynard, Z.; Bael, S.V.; Arnold, A.E.; Hebbar, P.; Samuels, G.J.; Robbins, N.; Herre, E.A. Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Hanada, R.E.; Souza, T.J.; Pomella, A.W.V.; Hebbar, K.P.; Pereira, J.O.; Ismaiel, A.; Samuels, G.J. Trichoderma martiale sp. nov., a new endophyte from sapwood of Theobroma cacao with a potential for biological control. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Hanada, R.E.; Pomella, A.W.V.; Costa, H.S.; Bezerra, J.L.; Loguercio, L.L.; Pereira, J.O. Endophytic fungal diversity in Theobroma cacao (cacao) and T. grandiflorum (cupuacu) trees and their potential for growth promotion and biocontrol of black-pod disease. Fungal Biol. 2010, 114, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.I.; Mead, A.J.; Cooke, A.W.; Dean, J.R. Mango stem end rot pathogens—Fruit infection by endophytic colonisation of the inflorescence and pedicel. Ann. Appl. Biol. 1992, 120, 225–234. [Google Scholar] [CrossRef]
- Mejia, L.C.; Herre, E.A.; Sparks, J.P.; Winter, K.; Garcia, M.N.; Van Bael, S.A.; Stitt, J.; Shi, Z.; Zhang, Y.; Guiltinan, M.J.; et al. Pervasive effects of an endophytic fungus on host genetic and phenotypic expression in a tropical tree. Front Microbiol. 2014, 5, 479. [Google Scholar] [CrossRef] [PubMed]
- Mejia, L.C.; Guiltinan, M.J.; Shi, Z.; Landherr, L.; Maximova, S.N. Expression of designed antimicrobial peptides in Theobroma cacao L. trees reduces leaf necrosis caused by Phytophthora spp. In Small Wonders; Peptides for Disease Control; Rajasekan, K., Cary, J.W., Jaynes, J.M., Montesinos, E., Eds.; American Chemical Society: Washington, DC, USA, 2012; pp. 379–395. [Google Scholar]
- Arnold, A.E.; Herre, E.A. Canopy cover and leaf age affect colonization by tropical fungal endophytes: Ecological pattern and process in Theobroma cacao (Malvaceae). Mycologia 2003, 95, 388–398. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylo-genetics. PCR Protoc. 1990, 315–322. [Google Scholar] [CrossRef]
- Bell, D.K.; Wells, D.H.; Markham, R.C. In vitro antagonism of Trichoderma species against six fungal plant pathogens. Phytopathology 1982, 72, 379–382. [Google Scholar] [CrossRef]
- Soytong, K. Identification of Species of Chaetomium in the Philippines and Screening for Their Biocontrol Properties against Seedborne Fungi of Rice. Ph.D. Thesis, University of the Philippines Los Baños, Laguna, Philippines, 1988. [Google Scholar]
- Asman, A.; Amin, A.; Abdullah, T. Endophytic fungi associated with cacao branch and their potential for biocontrol vascular streak dieback disease on cacao seedling. IOP Conf. Ser. Earth Environ. Sci. 2018, 157, 012039. [Google Scholar] [CrossRef] [Green Version]
- Rojas, E.I.; Rehner, S.A.; Samuels, G.J.; Van Bael, S.A.; Herre, E.A.; Cannon, P.; Chen, R.; Pang, J.; Wang, R.; Zhang, Y.; et al. Colletotrichum gloeosporioides s.l. associated with Theobroma cacao and other plants in Panama: Multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 2010, 102, 1318–1338. [Google Scholar] [CrossRef] [Green Version]
- James, R.S.; Ray, J.; Tan, Y.P.; Shivas, R.G. Colletotrichum siamense, C. theobromicola and C. queenslandicum from several plant species and the identification of C. asianum in the Northern Territory, Australia. Australas. Plant Dis. Notes 2014, 9, 138. [Google Scholar] [CrossRef] [Green Version]
- Crozier, J.; Thomas, S.E.; Aime, M.C.; Evans, H.C.; Holmes, K.A. Molecular characterization of fungal endophytic morpho species isolated from stems and pods of Theobroma cacao. Plant Pathol. 2006, 55, 783–791. [Google Scholar] [CrossRef]
- Rubini, M.R.; Silva-Ribeiro, R.T.; Pomella, A.W.V.; Maki, C.S.; Araujo, W.L.; Dos Santos, D.R.; Azevedo, J.L. Diversity of endophytic fungal community of cacao (Theobroma cacao L.) and biological control of Crinipellis perniciosa, causal agent of witches’ broom disease. Int. J. Biol. Sci. 2005, 1, 24–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashizume, Y.; Fukuda, K.; Sahashi, N. Effects of summer temperature on fungal endophyte assemblages in Japanese beech (Fagus crenata) leaves in pure beech stands. Botany 2010, 88, 266–274. [Google Scholar] [CrossRef]
- Kiewnick, S. Effect of temperature on growth, germination, germ tube extension and survival of Paecilomyceslilaci nus strain 251. Biocontrol Sci. Techn. 2006, 16, 535–546. [Google Scholar] [CrossRef]
- Elad, Y.; Kirshner, B. Survival in the phylloplane of an introduced biocontrol agent (Trichoderma harzianum) and populations of the plant pathogen Botrytis cinerea as modified by abiotic conditions. Phytoparasitica 1993, 21, 303–313. [Google Scholar] [CrossRef]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in Northern Thailand. Fungal Div. 2009, 39, 89–109. [Google Scholar]
- Manamgoda, D.S.; Udayanga, D.; Cai, L.; Chukeatirote, E.; Hyde, K.D. Endophytic Colletotrichum from tropical grasses with a new species C. endophytica. Fungal Div. 2013, 61, 107–115. [Google Scholar] [CrossRef]
- Sudarma, I.M.; Suniti, N.W.; Darmiati, N.N. Endophytic fungi with potential as biological control agents for black pod rot disease (P. palmivora Butler). Int. J. Curr. Res. Biosci. Plant Biol. 2018, 5, 41–48. [Google Scholar] [CrossRef]
- Sharma, G.; Shenoy, B.D. Colletotrichum fructicola and C. siamense are involved in chilli anthracnose in India. Arch. Phytopathol. Plant Prot. 2014, 47, 1179–1194. [Google Scholar] [CrossRef]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Capobiango, N.P.; Pinho, D.B.; Zambolim, O.L.; Lopes, U.P. Anthracnose on Strawberry Fruits Caused by Colletotrichum siamense in Brazil. The American Phytopathological Society (APS) Publication. Available online: https://www.apsnet.org (accessed on 20 February 2020).
- Chung, P.C.; Wu, H.Y.; Ariyawansa, H.A.; Tzean, S.S.; Chung, C.L. First Report of Anthracnose Crown Rot of Strawberry Caused by Colletotrichum siamense in Taiwan. The American Phytopathological Society (APS) Publications. Available online: https://apsjournals.apsnet.org (accessed on 25 February 2020).
- Ni, H.F.; Huang, C.W.; Wu, C.J.; Yang, H.R.; Lin, C.Y.; Chang, J.Y.; Chang, J.W. First report of pepper spot disease of lychee caused by Colletotrichum siamense in Taiwan. J. Plant Pathol. 2017, 99, 808. [Google Scholar]
- Chang, T.; Hassan, Q.; Jeon, J.Y.; Shin, J.S.; Oh, N.K.; Lim, T.H. First Report of Anthracnose of Persimmon (Diospyros kaki L. f.) Caused by Colletotrichum siamense in Korea. The American Phytopathological Society (APS) Publication. Available online: https://apsjournals.apsnet.org/ (accessed on 25 February 2020).
- Faeth, S.H. Are endophytic fungi defensive plant mutualists? OIKOS 2002, 98, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Loayza, P.; White, J.F., Jr.; Torres, M.S.; Balslev, H.; Kristiansen, T.; Svenning, J.C.; Gil, N. Light converts endosymbiotic fungus to pathogen, influencing seedling survival and niche space filling of a common tropical tree, Iriartea deltoidea. PLoS ONE 2011, 6, e16386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Photita, W.; Lumyong, S.; Lumyong, P.; McKenzie, E.H.C.; Hyde, K.D. Are some endophytes of Musa acuminata latent pathogens? Fungal Div. 2004, 16, 131–140. [Google Scholar]
- Promputtha, I.; Lumyong, S.; Dhanasekaran, V.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. A phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579–590. [Google Scholar] [CrossRef]
- Delaye, L.; Garcia-Guzman, G.; Heil, M. Endophytes versus biotrophic and necrotrophic pathogens–are fungal lifestyles evolutionarily stable traits? Fungal Div. 2013, 60, 125–135. [Google Scholar] [CrossRef]
- Chesters, C.G.C.; Hornby, D. Studies on Colletotrichum coccodes: II. Alternative host tests and tomato fruit inoculations using a typical root isolate. T. Brit. Mycol. Soc. 1965, 48, 583–594. [Google Scholar] [CrossRef]
- Sharma, P.N.; Katoch, A.; Sharma, P.; Sharma, S.K.; Sharma, O.P. First report on association of Colletotrichum coccodes with anthracnose in India (Abstract). Am. Phytopathol. Soc. 2011, 95, 584. [Google Scholar]
- Lu, H.; Zou, W.X.; Meng, J.C.; Hu, J.; Tan, R.X. New bioactive metabolites produced by Colletotrichum sp., an endophytic fungus in Artemisia annua. Plant Sci. 2000, 151, 67–73. [Google Scholar] [CrossRef]
- Ezhil Arivudainambi, U.S.; Anand, T.D.; Shanmugaiah, V.; Karunakaran, C.; Rajendran, A. Novel bioactive metabolites producing endophytic fungus Colletotrichum gloeosporioides against multidrug-resistant Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 2011, 61, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Rabha, A.J.; Naglot, A.; Sharma, G.D.; Gogo, H.K.; Veer, V. In vitro evaluation of antagonism of endophytic Colletotrichum gloeosporioides against potent fungal pathogens of Camellia sinensis. Indian J. Microbiol. 2014, 54, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Munasinghe, M.V.K.; Kumar, N.S.; Jayasinghe, L.; Fujimoto, Y. Indole-3-acetic acid production by Colletotrichum siamense, an endophytic fungus from Piper nigrum leaves. J. Biol. Act. Prod. Nat. 2017, 7, 475–479. [Google Scholar]
- Refaei, J.; Jones, E.B.G.; Sakayaroj, J.; Santhanam, J. Endophytic fungi from Rafflesia cantleyi: Species diversity and antimicrobial activity. Mycosphere 2011, 2, 429–447. [Google Scholar]
- Zou, X.; Meng, J.C.; Lu, H.; Chen, G.X.; Shi, G.X.; Zhang, T.Y.; Tan, R.X. Metabolites of Colletotrichum gloeosporioides, an endophytic fungus in Artemisia mongolica. J. Nat. Prod. 2000, 63, 1529–1530. [Google Scholar] [CrossRef]
- Serrato-Diaz, L.M.; Mariño, Y.A.; Guadalupe, I.; Bayman, P.; Goenaga, R. Colletotrichum siamense Causing Cacao Pod Rot and First Report of C. tropicale Causing Cacao Pod Rot in Puerto Rico. The American Phytopathological Society (APS) Publications. Available online: https://apsjournal.apsnet.org (accessed on 25 February 2020).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Sample | UF18 Clone | K9 Clone | ||
|---|---|---|---|---|
| 14 DAI | 26 DAI | 10 DAI | 20 DAI | |
| Inoculated | ||||
| 1 | + | + | + | − |
| 2 | − | − | − | − |
| 3 | − | − | + | − |
| 4 | + | + | − | − |
| 5 | − | + | + | − |
| Control | ||||
| 1 | − | − | − | − |
| 2 | − | − | − | − |
| 3 | − | − | − | − |
| 4 | − | − | − | − |
| 5 | − | − | − | − |
| Treatment | PIRG 1 | Bell’s Rating 2 | Antagonistic Activity 3 |
|---|---|---|---|
| C. siamense | 22.0b | 3 | Low |
| T. harzianum (Standard check) | 51.0a | 1 | Moderate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadoral, J.P.; Cumagun, C.J.R. Observations on the Potential of an Endophytic Fungus Associated with Cacao Leaves against Phytophthora palmivora. Microbiol. Res. 2021, 12, 528-538. https://doi.org/10.3390/microbiolres12030037
Sadoral JP, Cumagun CJR. Observations on the Potential of an Endophytic Fungus Associated with Cacao Leaves against Phytophthora palmivora. Microbiology Research. 2021; 12(3):528-538. https://doi.org/10.3390/microbiolres12030037
Chicago/Turabian StyleSadoral, Joan P., and Christian Joseph R. Cumagun. 2021. "Observations on the Potential of an Endophytic Fungus Associated with Cacao Leaves against Phytophthora palmivora" Microbiology Research 12, no. 3: 528-538. https://doi.org/10.3390/microbiolres12030037
APA StyleSadoral, J. P., & Cumagun, C. J. R. (2021). Observations on the Potential of an Endophytic Fungus Associated with Cacao Leaves against Phytophthora palmivora. Microbiology Research, 12(3), 528-538. https://doi.org/10.3390/microbiolres12030037