
Isolation, Description and Genome Analysis of a Putative Novel Methylobacter Species (‘Ca. Methylobacter coli’) Isolated from the Faeces of a Blackbuck (Indian Antelope)

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Enrichment and Isolation of a Methanotroph
2.3. Morphological Characterization
2.4. DNA Extraction and PCR Amplification
2.5. Biochemical Characterization
2.6. Strain Maintenance
3. Results
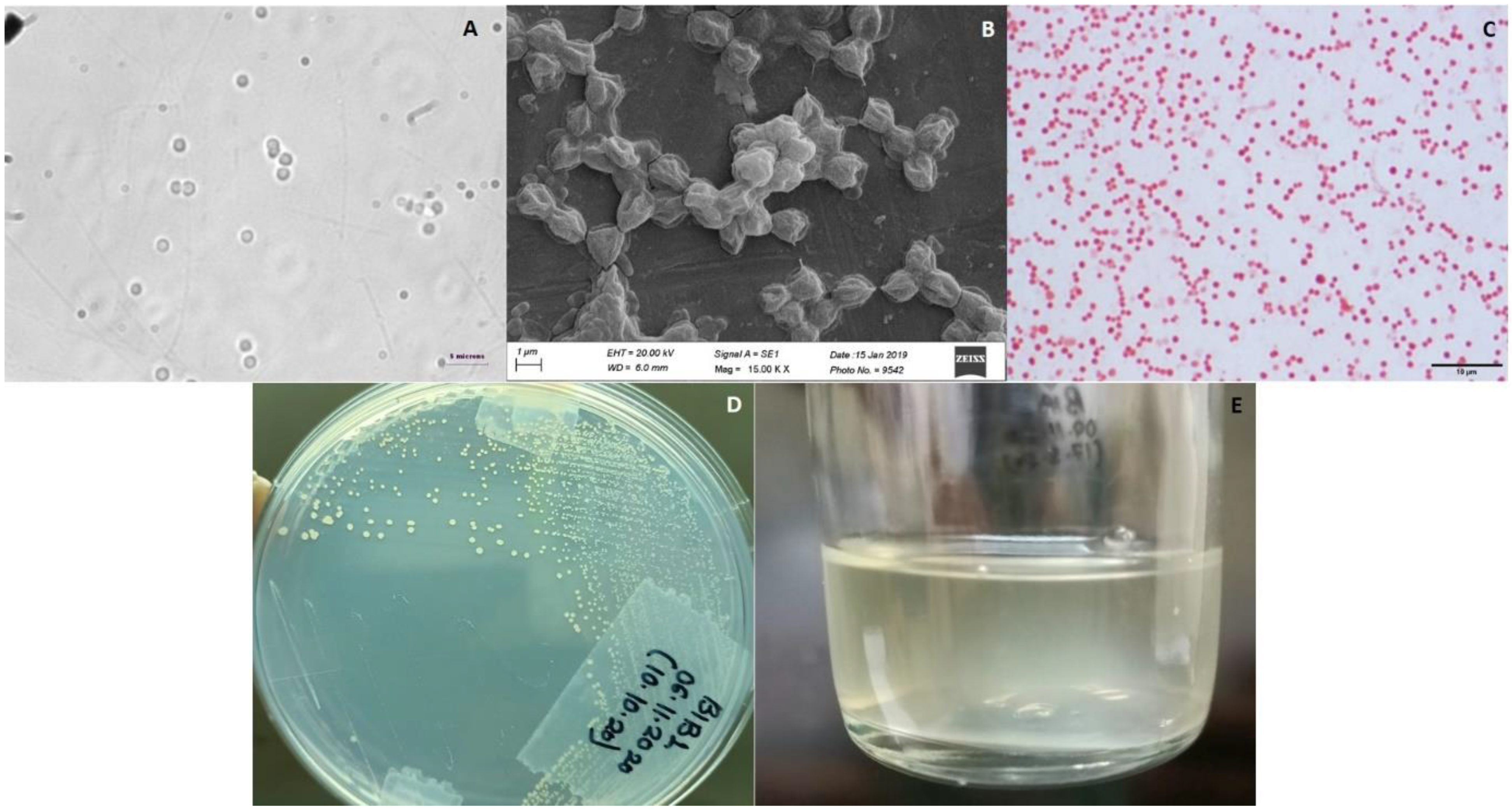
3.1. Enrichment, Isolation, and Identification
3.2. Microscopic Identification
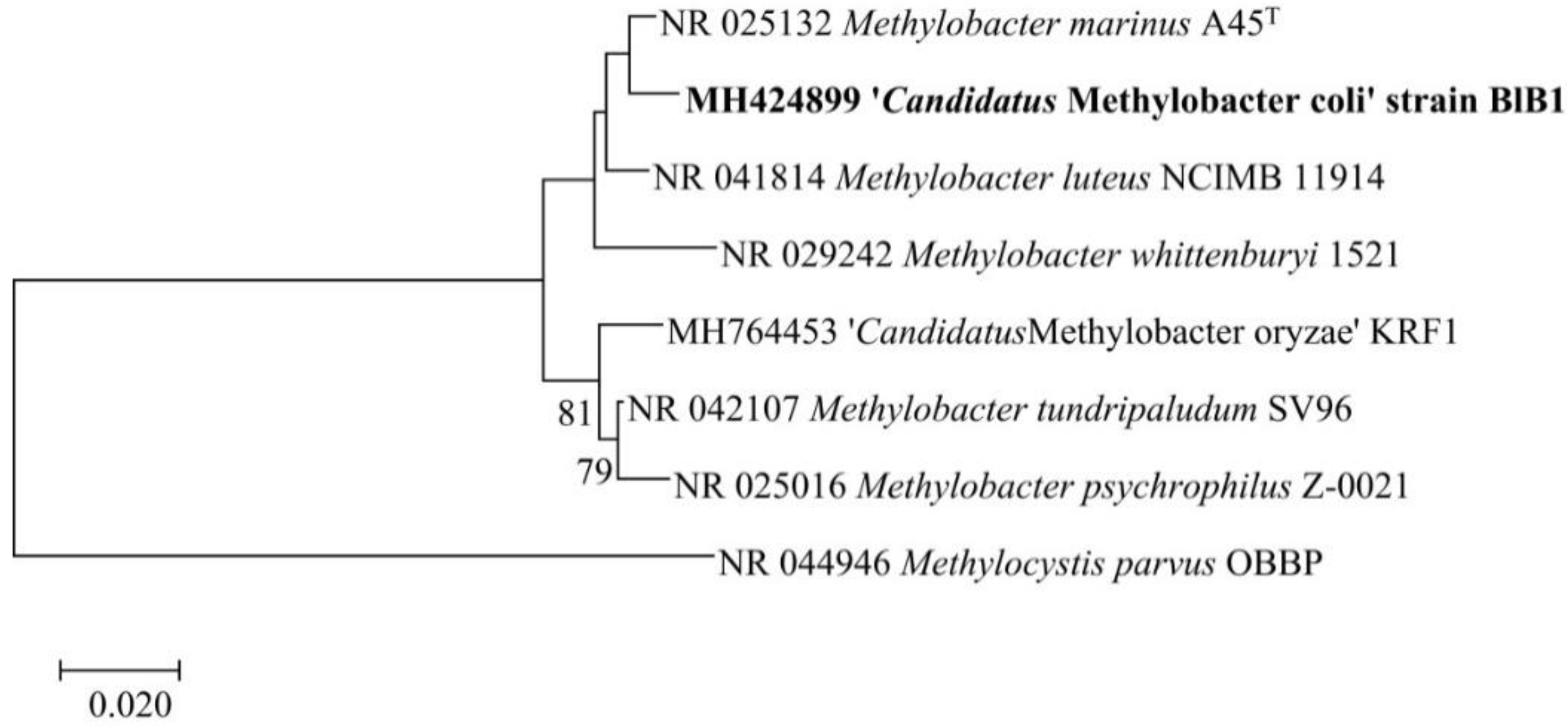
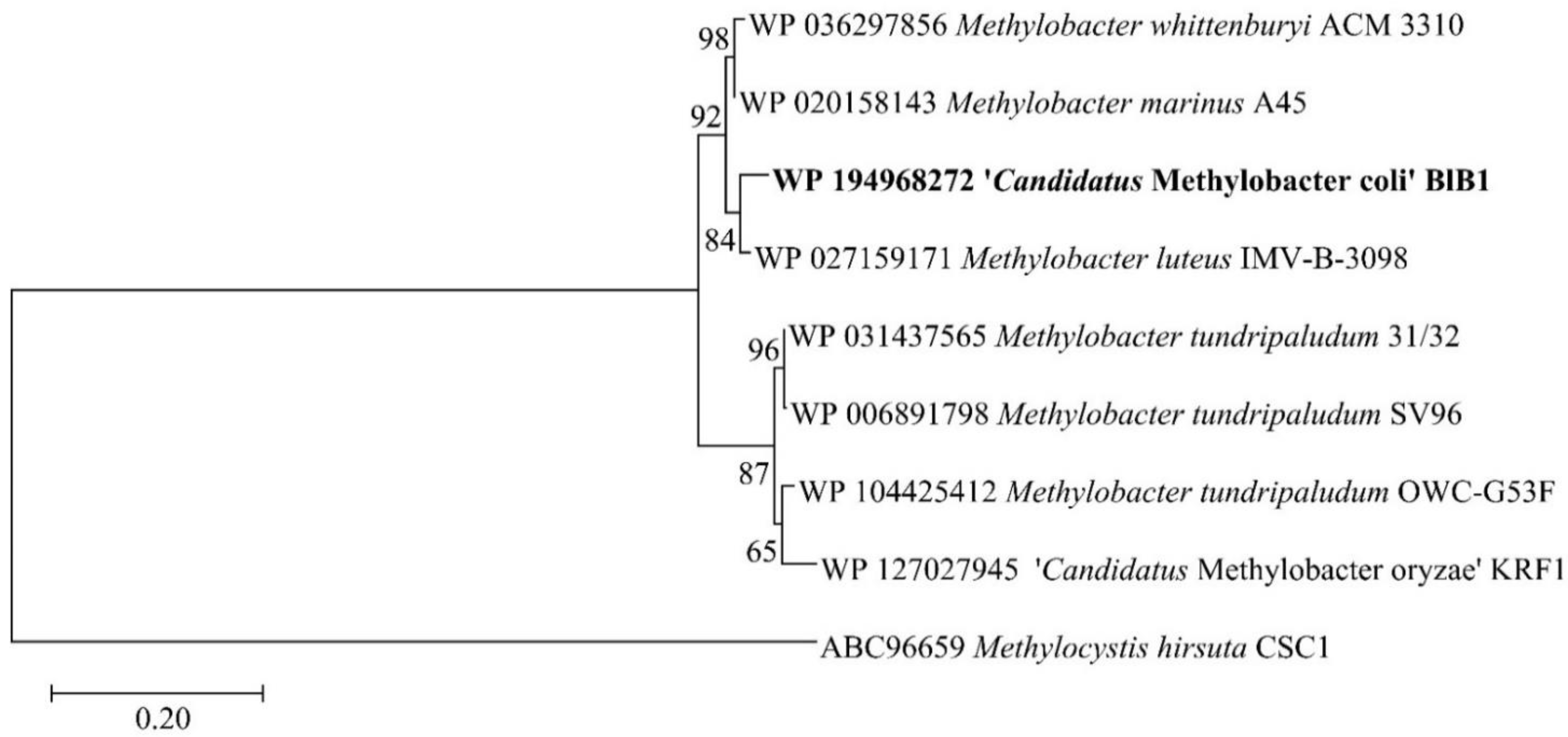
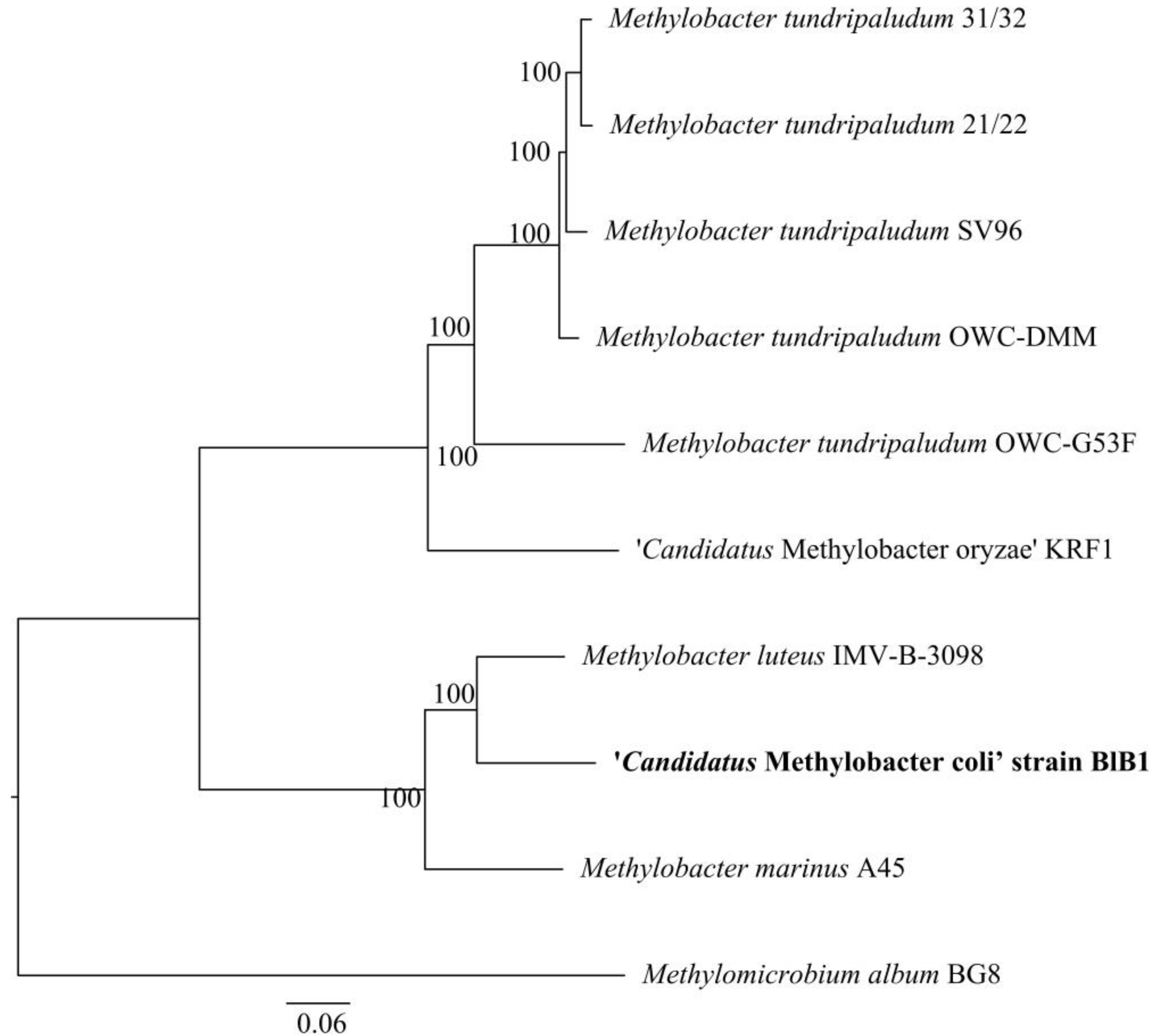
3.3. Phylogenetic and Phylogenomic Affiliation
3.4. Draft Genome Features and Salient Genome Characteristics
3.5. Carbon Metabolism Genes
3.6. Nitrogen Metabolism Genes
3.7. DDH, ANIb, and AAI Indicate That Strain BlB1 Represents a New Species of Methylobacter
3.8. Biochemical Characterisation
3.9. Description of ‘Candidatus Methylobacter coli’ Strain BlB1 Isolated from an Indian Blackbuck
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broucek, J. Production of methane emissions from ruminant husbandry: A review. J. Environ. Prot. 2014, 5, 1482–1493. [Google Scholar] [CrossRef]
- Conrad, R. The global methane cycle: Recent advances in understanding the microbial processes involved. Environ. Microbiol. Rep. 2009, 1, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, A. Enteric fermentation and ruminant eructation: The role (and control?) of methane in the climate change debate. Clim. Chang. 2009, 93, 407–431. [Google Scholar] [CrossRef]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef] [PubMed]
- Dedysh, S.N.; Knief, C. Diversity and Phylogeny of Described Aerobic Methanotrophs. In Methane Biocatalysis: Paving the Way to Sustainability; Springer: Berlin/Heidelberg, Germany, 2018; pp. 17–42. [Google Scholar]
- Finn, D.; Ouwerkerk, D.; Klieve, A. Methanotrophs from Natural Ecosystems as Biocontrol Agents for Ruminant Methane Emissions; Project Report; Meat & Livestock Australia Limited: North Sydney, Australia, February 2012. [Google Scholar]
- Oswald, K.; Milucka, J.; Brand, A.; Hach, P.; Littmann, S.; Wehrli, B.; Kuypers, M.M.; Schubert, C.J. Aerobic gammaproteobacterial methanotrophs mitigate methane emissions from oxic and anoxic lake waters. Limnol. Oceanogr. 2016, 61, S101–S118. [Google Scholar] [CrossRef]
- Stocks, P.K.; McCleskey, C. Morphology and physiology of Methanomonas methanooxidans. J. Bacteriol. 1964, 88, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Whittenbury, R.; Phillips, K.; Wilkinson, J. Enrichment, isolation and some properties of methane-utilizing bacteria. Microbiology 1970, 61, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Sauer, C.; Bertelsen, M.F.; Hammer, S.; Lund, P.; Weisbjerg, M.R.; Clauss, M. Macroscopic digestive tract anatomy of two small antelopes, the blackbuck (Antilope cervicapra) and the Arabian sand gazelle (Gazella subgutturosa marica). Anat. Histol. Embryol. 2016, 45, 392–398. [Google Scholar] [CrossRef]
- Pandit, P.S.; Rahalkar, M.C.; Dhakephalkar, P.K.; Ranade, D.R.; Pore, S.; Arora, P.; Kapse, N. Deciphering community structure of methanotrophs dwelling in rice rhizospheres of an Indian rice field using cultivation and cultivation-independent approaches. Microb. Ecol. 2016, 71, 634–644. [Google Scholar] [CrossRef]
- Pandit, P.S.; Hoppert, M.; Rahalkar, M.C. Description of ‘Candidatus Methylocucumis oryzae’, a novel Type I methanotroph with large cells and pale pink colour, isolated from an Indian rice field. Antonie Van Leeuwenhoek 2018, 111, 2473–2484. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Zuckerkandl, E.; Pauling, L. Evolutionary Divergence and Convergence in Proteins. In Evolving Genes and Proteins; Elsevier: Amsterdam, The Netherlands, 1965; pp. 97–166. [Google Scholar]
- Rossi-Tamisier, M.; Benamar, S.; Raoult, D.; Fournier, P.-E. Cautionary tale of using 16S rRNA gene sequence similarity values in identification of human-associated bacterial species. Int. J. Syst. Evol. Microbiol. 2015, 65, 1929–1934. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Yang, S.; Liebner, S. Evaluation and update of cutoff values for methanotrophic pmoA gene sequences. Arch. Microbiol. 2016, 198, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Kozliak, E.I.; Fuchs, J.A.; Guilloton, M.B.; Anderson, P.M. Role of bicarbonate/CO2 in the inhibition of Escherichia coli growth by cyanate. J. Bacteriol. 1995, 177, 3213–3219. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.A.; Otwinowski, Z.; Perrakis, A.; Anderson, P.M.; Joachimiak, A. Structure of cyanase reveals that a novel dimeric and decameric arrangement of subunits is required for formation of the enzyme active site. Structure 2000, 8, 505–514. [Google Scholar] [CrossRef][Green Version]
- Kao, W.-C.; Wang, V.C.-C.; Huang, Y.-C.; Yu, S.S.-F.; Chang, T.-C.; Chan, S.I. Isolation, purification and characterization of hemerythrin from Methylococcus capsulatus (Bath). J. Inorg. Biochem. 2008, 102, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.O.; Rosenzweig, A.C. A tale of two methane monooxygenases. J. Biol. Inorg. Chem. 2017, 22, 307–319. [Google Scholar] [CrossRef]
- Khatri, K.; Mohite, J.A.; Pandit, P.S.; Bahulikar, R.; Rahalkar, M.C. Description of ‘Ca. Methylobacter oryzae’ KRF1, a novel species from the environmentally important Methylobacter clade 2. Antonie Van Leeuwenhoek 2020, 113, 729–735. [Google Scholar] [CrossRef]
- Rahalkar, M.C.; Khatri, K.; Pandit, P.S.; Dhakephalkar, P.K. A putative novel Methylobacter member (KRF1) from the globally important Methylobacter clade 2: Cultivation and salient draft genome features. Antonie Van Leeuwenhoek 2019, 112, 1399–1408. [Google Scholar] [CrossRef]
- Deutzmann, J.S.; Hoppert, M.; Schink, B. Characterization and phylogeny of a novel methanotroph, Methyloglobulus morosus gen. nov., spec. nov. Syst. Appl. Microbiol. 2014, 37, 165–169. [Google Scholar] [CrossRef]
- Rahalkar, M.; Bussmann, I.; Schink, B. Methylosoma difficile gen. nov., sp. nov., a novel methanotroph enriched by gradient cultivation from littoral sediment of Lake Constance. Int. J. Syst. Evol. Microbiol. 2007, 57, 1073–1080. [Google Scholar] [CrossRef]
- Schink, B.; Rahalkar, M. Genus Methylosoma. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley Publications: Hoboken, NJ, USA, 2016; pp. 1–4. [Google Scholar]
- Oshkin, I.Y.; Miroshnikov, K.K.; Danilova, O.V.; Hakobyan, A.; Liesack, W.; Dedysh, S.N. Thriving in wetlands: Ecophysiology of the spiral-shaped methanotroph Methylospira mobilis as revealed by the complete genome sequence. Microorganisms 2019, 7, 683. [Google Scholar] [CrossRef]
- Collins, D.A.; Akberdin, I.R.; Kalyuzhnaya, M. Genus Methylobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley Publications: Hoboken, NJ, USA, 2017. [Google Scholar]
- Whitman, W.B.; Sutcliffe, I.C.; Rossello-Mora, R. Proposal for changes in the International Code of Nomenclature of Prokaryotes: Granting priority to Candidatus names. Int. J. Syst. Evol. Microbiol. 2019, 69, 2174–2175. [Google Scholar] [CrossRef]
- Niwiska, B. Digestion in Ruminants. In Carbohydrates-Comprehensive Studies on Glycobiology and Glycotechnology; Chang, C.-F., Ed.; InTech: Rijeka, Croatia, 2012; pp. 245–258. [Google Scholar]
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.; Yarlett, N.; Hillman, K.; Williams, A.; Lloyd, D.; Williams, T. The presence of oxygen in rumen liquor and its effects on methanogenesis. J. Appl. Bacteriol. 1983, 55, 143–149. [Google Scholar] [CrossRef]
- Dobson, A. Blood flow and absorption from the rumen. Q. J. Exp. Physiol. Transl. Integr. 1984, 69, 599–606. [Google Scholar] [CrossRef]
- Wallace, R.J.; Rooke, J.A.; McKain, N.; Duthie, C.-A.; Hyslop, J.J.; Ross, D.W.; Waterhouse, A.; Watson, M.; Roehe, R. The rumen microbial metagenome associated with high methane production in cattle. BMC Genom. 2015, 16, 452. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Closely Related Members | ANIb Value | AAI Value | DDH Value |
|---|---|---|---|
| Methylobacter sp. BlB1 | - | - | - |
| Methylobacter marinus A45 | 83.07 | 83.96% | 27.40% [25–29.9%] |
| Methylobacter tundripaludum OWC-DMM | 74.78 | 71.43% | 21.10% [18.9–23.5%] |
| Methylobacter tundripaludum SV96 | 74.62 | 71.52% | 21.10% [18.8–23.5%] |
| Methylobacter tundripaludum 31/32 | 74.55 | 71.02% | 21.10% [18.8–23.5%] |
| Methylobacter tundripaludum 21/22 | 74.61 | 71.5% | 21.00% [18.7–23.4%] |
| Methylobacter tundripaludum OWC-G53F | 74.95 | 70.83% | 20.50% [18.2–22.9%] |
| Candidatus Methylobacter oryzae KRF1 | 73.71 | 70.43% | 20.20% [18–22.6%] |
| Parameters | Range | Optimum Condition | Final OD600 |
|---|---|---|---|
| Temperature | 30°–39° | 30° (pH 7.0) | 0.16 |
| pH | Buffered 4–9; Unbuffered 6.8–10 | Buffered–6.8; Unbuffered–6.8 | 0.20; 0.19 |
| Salt tolerance | 0.1–0.5% | 0% | 0.16 |
| Methanol tolerance | 0.01–5% | 0.2% | 0.25 |
| Air concentration | 5–80% | 80% | 0.16 |
| Features | Strain BlB1 | Mtb marinus A45T [28] | Mtb luteus NCIMB 11914 [28] | Ca. Mtb oryzae KRF1T [22] | Mtb tundripaludum SV96T [28] | Mtb psychrophilus Z-0021T [28] | Mtb whittenburyi 1521 [28] |
|---|---|---|---|---|---|---|---|
| Cell Morphology | Cocci, diplococcic cells, long chains of cocci | Rods and elliptical rods | Elliptical rods and cocci | Rods | Rods and elliptical rods | Rods and elliptical rods and cocci | Rods and elliptical rods and cocci |
| Cell size | 1.5–2 µm in diameter | 0.8–1.2 × 1.5–2.0 μm | 1.2–1.5 × 1.5–2.0 μm | 0.9–1.2 × 3-5 µm | 0.8–1.3 × 1.9–2.5 μm | 1.0–1.7 μm in diameter | 1.2–1.5 × 1.5–3.0 μm |
| Motility | No | Yes | No | Yes | Variable | No | Variable |
| Pigmentation | Yellow | White, pink, creamy | Yellow | Yellow | Pale pink | White/Buff | Cream, Brown |
| Optimum temperature (°C) | 30 | 35 | 30 | 20–25 | 23 | 3.5–10 | 30–35 |
| nifH gene | Yes | Yes | Yes | Yes | Yes | No | Yes |
| G + C Content (%) | 51.3 | 52.3–52.7 | 50.3–51.1 | 49.3 | 49.2–49.7 | 45.6 | 48.7–53.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatri, K.; Mohite, J.; Pandit, P.; Bahulikar, R.A.; Rahalkar, M.C. Isolation, Description and Genome Analysis of a Putative Novel Methylobacter Species (‘Ca. Methylobacter coli’) Isolated from the Faeces of a Blackbuck (Indian Antelope). Microbiol. Res. 2021, 12, 513-523. https://doi.org/10.3390/microbiolres12020035
Khatri K, Mohite J, Pandit P, Bahulikar RA, Rahalkar MC. Isolation, Description and Genome Analysis of a Putative Novel Methylobacter Species (‘Ca. Methylobacter coli’) Isolated from the Faeces of a Blackbuck (Indian Antelope). Microbiology Research. 2021; 12(2):513-523. https://doi.org/10.3390/microbiolres12020035
Chicago/Turabian StyleKhatri, Kumal, Jyoti Mohite, Pranitha Pandit, Rahul A. Bahulikar, and Monali C. Rahalkar. 2021. "Isolation, Description and Genome Analysis of a Putative Novel Methylobacter Species (‘Ca. Methylobacter coli’) Isolated from the Faeces of a Blackbuck (Indian Antelope)" Microbiology Research 12, no. 2: 513-523. https://doi.org/10.3390/microbiolres12020035
APA StyleKhatri, K., Mohite, J., Pandit, P., Bahulikar, R. A., & Rahalkar, M. C. (2021). Isolation, Description and Genome Analysis of a Putative Novel Methylobacter Species (‘Ca. Methylobacter coli’) Isolated from the Faeces of a Blackbuck (Indian Antelope). Microbiology Research, 12(2), 513-523. https://doi.org/10.3390/microbiolres12020035