Abstract
Taxonogenomics combines phenotypic assays and genomic analysis as a means of characterizing novel strains. We used this strategy to study Marseille-P9898T strain, an aerobic, motile, Gram-negative, spore-forming, and rod-shaped bacterium isolated from planarian Schmidtea mediterranea. Marseille-P9898T is catalase-positive and oxidase-negative. The major fatty acids detected are 12-methyl-tetradecanoic acid, 13-methyl-tetradecanoic acid, and hexadecanoic acid. Marseille-P9898T strain shared more than 98% sequence similarity with the Metabacillus niabensis strain 4T19T (98.99%), Metabacillus halosaccharovorans strain E33T (98.75%), Metabacillus malikii strain NCCP-662T (98.19%), and Metabacillus litoralis strain SW-211T (97.15%). Marseille-P9898 strain belongs to Metabacillus genus. Genomic analysis revealed the highest similarities with Ortho-ANI and dDDH, 85.76% with Metabacillus halosaccharovorans, and 34.20% with Bacillus acidicola, respectively. These results show that the Marseille-P9898T strain is a novel bacterial species from Metabacillus genus, for which we propose the name of Metabacillus schmidteae sp. nov. (Type strain Marseille-P9898T = CSUR P9898T = DSM 111480T).
1. Introduction
The birth of genomics, followed by the development of Next Generation Sequencing (NGS) methods, has allowed the characterization, classification, and nomenclature of many prokaryotic species using a taxonogenomics strategy combining phenotypic assays and genome sequencing [1,2,3]. Phenotypic assays are based on the analysis of morphological, physiological, chemical, and biochemical features of an organism [4]. Genotypic characterization, i.e., the analysis of the genetic material, is essential for species description and sheds light on the evolutionary relationships between various lineages. For the genotypic characterization, first, the gene 16S rRNA sequences are used to determine sequence similarity and for phylogenetic analysis [5]. Then Digital DNA-DNA hybridization values (dDDH) that evaluate the degree of genetic similarity between two genomes have been used for bacterial species demarcation by providing a constant numerical threshold (dDDH value > 70%) for species boundary [6].
Planarian Schmidtea mediterranea is an invertebrate living in freshwater and an excellent organism model to investigate the human host-pathogen relationship [7,8,9]. To understand the role of planarian S. mediterranea microbiota in the immune response, we studied its composition and isolated by culturomics [10], a bacterial strain (Marseille-P9898).
Analysis of Marseille-P9898 strain by Matrix-Assisted Laser Desorption-Ionization Time of Flight Mass Spectrometry (MALDI-TOF MS) did not allow identifying this strain, but it revealed that it belongs to the Bacillus genus (now Metabacillus [11]). Bacteria belonging to the Bacillus genus [12] are obligate aerobic, endospore-forming bacilli. This genus was first described by Cohn F. in 1872 [13]. The type strain is Bacillus subtilis [14]. Recently, phylogenomic and comparative genomic analyses showed six novel genera of Bacillus species: Peribacillus, Cytobacillus, Mesobacillus, Neobacillus, Alkalihalobacillus, and Metabacillus [11]. According to its first characteristics, Marseille-P9898 strain could be clustered among Metabacillus (Me.ta.ba.cil’lus. Gr. adv. meta besides; L. masc. n. bacillus a small staff or rod, and Bacillus, a bacterial genus; N.L. masc. n. Metabacillus a genus besides Bacillus), which was approved in the family Bacillaceae [15], belonging to the phylum Firmicutes [16]. The majority of bacteria from this genus are frequently isolated from hypersaline and saline environments, mainly saline soils, and saline aquatic habitats [17,18,19,20]. These bacteria are obligate aerobic or facultative anaerobic [12] and moderately halophilic or halotolerant [21].
In this study, we have characterized the taxonogenomic properties of this strain, designated Marseille-P9898 (=CSUR P9898T = DSM 111480T), which is a novel bacterial species named Metabacillus schmidteae sp. nov.
2. Materials and Methods
2.1. Planarian Schmidtea Mediterranea Culture
S. mediterranea flatworms (asexual clonal line ClW4) were kept in tap water filtered at 19 °C. Filtered water was obtained by filtration through a filter containing charcoal and ceramics (Fairey Industrial Ceramics limited, Suffolk, England), and a 0.2 µm membrane (Thermo Scientific Nalgene filtration Products, Mexico City, Mexico). The sterility of filtered water was analyzed before any utilization. Microbiological analysis of filtered water was performed by inoculation of filtered water (25, 50, 75, and 100 µL) in 5 % sheep blood-enriched Columbia agar plate (bioMérieux, Marcy l’étoile, France) and by incubation at various temperatures (5, 10, 19, 28, 37 and 45 °C). After four days, the absence of bacterial colonies was assessed.
2.2. Culture Conditions from Schmidtea mediterranea and Strain Isolation
Worms were starved for 2 weeks, washed in filter-sterilized water. One worm was smashed in a tube with a pestle in 500 µL of PBS, then 100 µL of the crushed mixture was inoculated in 5% sheep blood-enriched Columbia agar (bioMérieux, Marcy l’étoile, France) and incubated at 28 °C. Individual bacterial colonies were harvested and identified by MALDI-TOF-MS (Microflex spectrometer; Bruker Daltonics, Bremen, Germany) [22]. The MALDI Biotyper RTC software was used to interpret results according to obtained score values. The database used was the MaldiBiotyperDBUpdate_V9. 8468 MSP from the Bruker Daltonics. A bacterial colony is likely identified at the species level for a score ≥ 2.0; probably identified for a score between 1.99 and 1.7, but not identified for a score < 1.7. Profile spectrum of the whole cell was obtained after analysis as previously described [22].
2.3. DNA Extraction, Sequencing, Assembly, and Annotation
Genomic DNA extraction (gDNA) from the Marseille-P9898 strain was performed using an EZ1 BioRobot and EZ1 DNA tissue kit (Cat No./ID: 953034, Qiagen, Hilden, Germany). Then gDNA was quantified by a Qubit assay (Life Technologies, Carlsbad, CA, USA). Here, we used two technologies for Marseille-P9898 strain sequencing. First, gDNA was normalized at 0.2 ng/µL, then prepared and sequenced using the Mate-Pair strategy with a Miseq sequencer (Illumina, San Diego, CA, USA), as previously described [23]. Second, gDNA was normalized at 1.5 ng/µl and sequenced using nanopore strategy with a Minion sequencer (Oxford Nanopore MinION™) [24,25]. The reads of Miseq and MinION run were examined using FastQC 0.11.8 to evaluate quality [26]. Two sequencing reads were assembled using Spades [27] genome assembler software for regular and single-cell projects (Galaxy 3.12.0+galaxy1). The “conservative” option was used to reduce mismatches number and short indels. Default parameters were applied for each software. Genomic annotation was obtained using Prokka [28] software (Galaxy version 1.14.5+galaxy1).
2.4. Phylogenetic Analysis
For the taxonomic assignment, we used the nr database (Standard databases) for the BLASTn search. A sequence similarity threshold of 98.65% by comparison with the phylogenetically closest species with standing in nomenclature was used to delineate a putative novel species [29]. Phylogenetic relationships were inferred from the comparison of 16S rRNA sequences using MEGAX 10.1 software [30,31]. The method for estimating phylogenetic trees was maximum likelihood. Sequences were aligned using the MUSCLE algorithm with default parameters. Numbers shown at the nodes are percentages of bootstrap values obtained by repeating the analysis 1000 times to generate a majority consensus tree. Only bootstrap values ≥ 50% were retained. Scale bare indicates a 0.01% sequence divergence.
2.5. Genomic Comparison
The degree of genomic similarity was evaluated using GGDC [32] (http://ggdc.dsmz.de/ggdc.php, GGDC Genome-to-Genome Distance Calculator 2.1 and Orthologous Average Nucleotide Identity [33] (https://www.ezbiocloud.net/tools/orthoani, accessed on 1 January 2021, OrthoANI Tool version 0.93.1) software. Comparison of COG functional categories was carried out using BLASTP (E-value 10−3, coverage 0.7 and identity percent 30%) against the clusters of Metabacillus orthologous groups (COG) database.
2.6. Phenotypic and Biochemical Features
The growth of the Marseille-P9898 strain was analyzed at different temperatures (4, 19, 28, 30, 37, and 45 °C) in 5% sheep blood-enriched Columbia agar (bioMérieux) under anaerobic and aerobic atmospheres using GasPak™ EZ generators (Becton-Dickinson, Marynland, MD, USA). The capacity to grow in distinct salinity (0, 20, 40, 50, 60, 80 and 100 g of NaCl/l) and pH (5, 5.5, 6, 6.5, 7.5, 8.5, 9, and 10) conditions were tested. Sporulation capacity was tested by thermal shock. For this purpose, bacteria were exposed to 80 °C temperature for 30 min, and then bacterial growth was monitored for 4 days. Gram staining and motility of fresh colonies were observed using a DM1000 light microscope (Leica Microsystems, Nanterre, France) with 100× lens and oil immersion. To evaluate bacterial structures, we used scanning electron microscopy (Hitachi SU5000) (Hitachi High-Technologies Corporation, Tokyo, Japan). Catalase and oxidase activities were tested using a BBL DrySlide according to the manufacturer’s instructions (Becton Dickinson, Le Pont de Claix, France). API strips (API ZYM [34,35,36], API 20NE [37,38], API 20E [39,40] and API 50CH [41,42,43,44], bioMérieux) were used to study the biochemical characteristics for Marseille-P9898 strain and Metabacillus niabensis strain 4T19T [45].
2.7. Antibiotic Susceptibility
Bacterial susceptibility to benzylpenicillin, amoxicillin, ampicillin, ceftriaxone, imipenem, ciprofloxacin, amikacin, gentamicin, streptomycin, daptomycin, doxycyclin, metronidazole, rifampicin, fosfomycin, vancomycin, and tigecycline was assessed using E-tests. For this purpose, we used a 0.5 McFarland concentration of Marseille-P9898 strain and Metabacillus niabensis strain 4T19T.
2.8. Chemotaxonomic Analysis
Cellular fatty acid methyl ester (FAME) analysis of Marseille-P9898 strain and Metabacillus niabensis strain 4T19T was performed by GC/MS. Fatty acid methyl esters were prepared as published by Sasser [46] and GC/MS analysis was realized as previously described [47]. Briefly, fatty acid methyl esters were separated using an Elite 5-MS column and monitored by mass spectrometry (Clarus 500-SQ 8 S, Perkin Elmer, Courtaboeuf, France). Spectral database search was performed using MS Search 2.0 operated with the Standard Reference Database 1A (NIST, Gaithersburg, USA) and the FAMEs mass spectral database (Wiley, Chichester, UK).
3. Results
3.1. Phenotypic and Biochemical Characteristics
The Marseille-P9898 strain was isolated on 5% Columbia agar enriched with sheep blood (bioMérieux) after 2 days of growth at 28 °C in an aerobic atmosphere. This strain grows at a temperature ranging from 19 to 50 °C, and under pH from 7.5 to 10 (alkaline). Marseille-P9898 strain can grow at NaCl concentration lower than 10 g/L (Table 1). After 4 days of incubation at 28 °C on blood-enriched Columbia agar, Marseille-P9898 strain colonies were white, round, and smooth, with a 3–5 mm diameter and a convex shape (Supplementary data Figure S1). Electron microscopy revealed that Marseille-P9898 strain cells are rod-shaped and have a 2.70 µm mean length, a 0.42 µm mean width, and a polar flagellum (Figure 1). Bacterial cells are Gram-negative (Figure 2), motile, and spore-forming bacilli (Supplementary data Figure S2). The endospore formation occurs in the terminal position. Marseille-P9898 strain is catalase positive and oxidase negative. Bacterial metabolism was assessed using API 50CHB/E, API 20NE, API Zym, and API 20E strips (Table 2). Marseille-P9898 strain differs from Metab. halosaccharovorans, Metab. niabensis, Metab. malikii and Metab. litoralis compared to α-glucosidase, L-rhamnose, methyl-αD-glucopyranoside, glycogen, potassium gluconate, natriumpyruvat, glucose, trisodium citrate, because Marseille-P9898 strain needs these substrates to grow. Interestingly, identification by MALDI-TOF-MS of Marseille-P9898 strain showed a score of 1.77, matching with Metabacillus niabensis (spectrum in supplementary data Figure S3). Because this value was less than 2, the Marseille-P9898 strain cannot be identified as Metabacillus niabensis. This score only indicates that the Marseille-P9898 strain belongs to the Metabacillus genus. To define the species type, we performed a genomic approach.

Figure 1.
Transmission electron microscopy of Marseille-P9898 strain. Bacteria are rod-shaped and exhibit a polar flagellum (black arrow). Scale bar = 10.0 µm.
Figure 2.
Gram staining of Marseille-P9898 strain.
3.2. Phylogenetic Analysis
The gene 16S rRNA sequence from the Marseille-P9898 strain had a size of 1548 bp. Using phylogenetic comparison with the GenBank database, we found that Marseille-P9898 strain had similarity at the level of an 16S rRNA gene sequence with Metabacillus niabensis strain 4T19T [45] (98.99%), Metab. halosaccharovorans strain E33T [48] (98.75%), Metab. malikii strain NCCP-662T [49] (98.19%), Metab. litoralis strain SW-211T [50] (97.15%), Bacillus frigoritolerans strain DSM 8801T [51,52] (95.99%), Mesob. foraminis strain CV53T [53] (95.97%), Mesob. subterraneus strain COOI3BT [54] (95.85%), Metab. herbersteinensis strain D-15aT [55] (97.21%), Neob. niacini strain IFO15566T [56] (95.95%), B. acidicola strain 105-2T [57] (95.36%), B. oryzisoli strain 1DS3-10T [58] (96.10%), B. flexus strain IFO15715T [59] (95.69%), Metab. indicus strain Sd/3T [60] (96.21%), Cytob. gottheilii strain WCC 4585T [61] (95.91%), B. horikoshii strain DSM 8719T [62] (95.55%), Perib. simplex DSM 1321T [59] (95.68%), B. tianshenii strain YIM M13235T [63] (95.27%), Metab. fastidiosus strain DSM91T (96.48%), B. aryabhattai B8W22T [64] (95.50%), Metab. endolithicus strain JC267T [65] (98.21%), Neob. bataviensis strain NBRC 102449T [66] (95.94%), Metab. galliciensis strain BFLP-1T [67] (96.18%), Neob.. novalis strain IDA3307T [66] (95.89%), B. licheniformis DSM 13T (94.45%), and Metab. crassostreae JSM 100118T [68] (98.07%). The most closely related species to the Marseille-P9898 strain was the Metab. niabensis strain 4T19T, Metab. halosaccharovorans strain E33T, Metab. malikii strain NCCP-662T and Metab. litoralis strain SW-211T. A phylogenetic tree based on the maximum-likelihood algorithm revealed that the Marseille-P9898 strain is shared in a cluster of members of the Metabacillus genus (Figure 3), belonging from Bacillaceae family and Firmicutes phylum. On the phylogenetic tree, it was clear that the Marseille-P9898 strain presented a distinct taxon separated from other species of genus Bacillus. We selected nine closely related species to make a comparison genomic (Table 3 and Supplementary Data Table S1).

Figure 3.
Phylogenetic tree carried out from a comparative analysis of 16S rRNA gene sequences indicating the relationships between Marseille-P9898 strain and related species. Sequences are aligned using the MUSCLE algorithm with default parameters and phylogenies are inferred by the MEGAX software (version 10.1). Numbers at the nodes are percentages of bootstrap values obtained by repeating the analysis 1000 times to generate a majority consensus tree. Only bootstrap values ≥ 50% are retained. Scale bare indicates a 0.01% sequence divergence.

Table 1.
Characteristics and classification of strain Marseille-P9898.
Table 1.
Characteristics and classification of strain Marseille-P9898.
| Property | Term |
|---|---|
| Current classification | Kingdom: Bacteria [69] |
| Phylum: Firmicutes [16] | |
| Class: Bacilli [70] | |
| Order: Bacillales [71] | |
| Family: Bacillaceae [15] | |
| Genus: Metabacillus [11] | |
| Species name: Schmidteae | |
| Specific epithet: Metabacillus schmidteae | |
| Type strain: Marseille-P9898 | |
| Species status | sp. nov. |
| Gram stain | Negative |
| Cell shape | Rod-shape |
| Motility | Motile |
| Sporulation | Spore-forming |
| Temperature range for growth | 19 to 50 |
| Temperature optimum | 28 |
| pH range for growth | 7.5 to 10 |
| pH optimum | 8 |
| pH category | alkaline |
| Lowest NaCl concentration for growth | 0 |
| Highest NaCl concentration for growth | 12 g/L |
| Salinity optimum | 10 g/L |
| O2 conditions for strain testing | Strict aerobic |
| Catalase | Positive |
| Oxidase | Negative |
| Habitat | Planarians |
| Biotic relationship | Microbiota planarian |
Table 2.
Biochemical characteristics of Marseille-P9898 and phylogenomically related species. Taxa: 1, Marseille-P9898; 2, Metabacillus niabensis 4T19T; 3, Metabacillus halosaccharovorans E33T; 4, Metabacillus litoralis SW-211T. Data of E33T and SW-211T strains were obtained using previously published work [48,50]. Data of Marseille-P9898 and 4T19T strains were obtained in the present study. +, positive; −, negative; NA, data not available.
Table 2.
Biochemical characteristics of Marseille-P9898 and phylogenomically related species. Taxa: 1, Marseille-P9898; 2, Metabacillus niabensis 4T19T; 3, Metabacillus halosaccharovorans E33T; 4, Metabacillus litoralis SW-211T. Data of E33T and SW-211T strains were obtained using previously published work [48,50]. Data of Marseille-P9898 and 4T19T strains were obtained in the present study. +, positive; −, negative; NA, data not available.
| Property | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Gram-staining | − | + | + | − |
| Mobility | Motile | Motile | Motile | Motile |
| Sporulation | + | + | + | + |
| Growth temperature range (°C) | 19–50 | 15–40 | 20–45 | 4–45 |
| Aerobic growth | Strict aerobic | Aerobic | Strict aerobic | Strict aerobic |
| Source | Planarians | Cotton waste | Lake | Tidal flat |
| Colony color | White | Yellowish white | Cream | Yellowish white |
| Catalase | + | + | + | + |
| Oxidase | − | − | + | + |
| ENZYMATIC ACTIVITIES | ||||
| 2-nitrophenyl-βD-galactopyranoside (ONPG) | − | + | NA | NA |
| 4-Nitrophenyl-βD-Galactopyranoside | + | + | NA | NA |
| Acid phosphatase | − | − | NA | + |
| Alkaline phosphatase | + | − | NA | + |
| Cystine arylamidase | − | − | − | − |
| Esculin ferric citrate (hydolyse) | + | + | + | NA |
| Esterase (C4) | + | + | NA | + |
| Esterase lipase (C8) | + | + | NA | + |
| Gelatin | − | − | + | + |
| Indole production | − | − | − | − |
| L-arginin | − | − | − | − |
| Leucine arylamidase | − | − | − | − |
| Lipase (C14) | − | − | NA | − |
| L-lysin | − | − | − | − |
| L-ormithin | − | − | − | − |
| N-acetyl-β-glucosaminidase | − | − | NA | − |
| Naphtol-AS-BI-phosphohydrolase | + | + | NA | + |
| Natriumpyruvat | + | − | NA | − |
| Natriumthiosulfat | − | − | − | − |
| Trinatriumcitrat | − | − | NA | NA |
| Trypsin | − | − | NA | − |
| Urea | − | − | − | − |
| Valine arylamidase | − | − | − | − |
| α-chymotrypsin | − | − | NA | − |
| α-fucosidase | − | − | NA | − |
| α-galactosidase | − | − | NA | − |
| α-glucosidase | + | − | NA | − |
| α-mannosidase | − | − | NA | − |
| β-glucosidase | − | + | NA | − |
| β-glucuronidase | + | − | NA | − |
| CARBOHYDRATE ASSIMILATION | ||||
| Adipic acid | − | − | NA | NA |
| Amygdalin | + | + | NA | NA |
| Arbutin | − | + | NA | NA |
| Capric acid | − | − | NA | NA |
| D-adonitol | − | − | NA | − |
| D-arabinose | − | − | + | NA |
| D-arabitol | + | − | NA | NA |
| D-cellobiose | + | + | + | + |
| D-fructose | + | + | + | + |
| D-fucose | − | − | NA | NA |
| D-galactose | + | + | + | + |
| D-glucose | + | + | + | + |
| D-lactose | + | + | + | + |
| D-lyxose | + | − | NA | NA |
| D-maltose | + | − | + | + |
| D-mannitol | + | − | + | − |
| D-mannose | + | + | + | − |
| D-melezitose | + | − | NA | + |
| D-melibiose | + | + | + | + |
| D-raffinose | + | + | + | + |
| D-ribose | − | − | + | + |
| D-saccharose | + | + | + | + |
| D-sorbitol | − | + | NA | − |
| D-tagatose | − | − | NA | NA |
| D-trehalose | + | + | + | + |
| D-turanose | − | − | NA | NA |
| Dulcitol | − | − | NA | NA |
| D-xylose | + | + | + | + |
| Erythritol | − | − | NA | NA |
| Gentiobiose | + | + | NA | NA |
| Glycerol | + | + | + | NA |
| Glycogen | − | + | NA | NA |
| Inositol | − | + | NA | − |
| Inulin | + | + | NA | NA |
| L-arabinose | + | - | + | + |
| L-arabitol | − | − | NA | NA |
| L-fucose | − | − | NA | NA |
| L-rhamnose | − | + | NA | + |
| L-sorbose | − | − | NA | NA |
| L-xylose | − | − | + | NA |
| Malic acid | + | − | NA | + |
| Methyl-αD-glucopyranoside | + | − | − | NA |
| Methyl-αD-mannopyranoside | − | − | − | NA |
| Methyl-βD-xylopyranoside | + | + | NA | NA |
| N-Acetyl-glucosamine | + | − | NA | NA |
| Phenylacetic acid | − | − | NA | NA |
| Potassium 2-ketoGluconate | − | − | NA | NA |
| Potassium 5-ketogluconate | − | − | NA | NA |
| Potassium gluconate | + | − | NA | NA |
| Salicin | + | + | + | NA |
| Starch | + | + | + | + |
| Trisodium citrate | + | − | NA | − |
| Xylitol | − | − | NA | NA |
| SUBSTRAT REDUCTION | ||||
| L-tryptophan | + | + | NA | − |
| Potassium nitrate | − | + | − | NA |
Table 3.
Main genomic characteristics of strain Marseille-P9898 and closely related species.
Table 3.
Main genomic characteristics of strain Marseille-P9898 and closely related species.
| Name | Size (bp) | GC% | Contigs | Refseq |
|---|---|---|---|---|
| Bacillus flexus | 3,906,163 | 37.6 | 259 | BCVD01000001.1 |
| Bacillus acidicola | 5,137,992 | 39.4 | 10 | LWJG01000001.1 |
| Mesobacillus foraminis | 5,730,823 | 43.0 | 35 | SLVV01000001.1 |
| Metabacillus halosaccharovorans | 5,399,327 | 36.1 | 8 | MTIR01000001.1 |
| Metabacillus litoralis | 5,230,624 | 35.9 | 1 | NZ_CP033043.1 |
| Metabacillus niabensis | 4,987,608 | 35.5 | 462 | NZ_CADEPK010000000 |
| Neobacillus niacini | 2,201,253 | 38.3 | 143 | JRYQ01000001.1 |
| Metabacillus schmidteae | 5,499,502 | 35.8 | 3 | CAESCH000000000.1 |
| Mesobacillus subterraneus | 4,571,170 | 43.9 | 42 | RSFW01000001.1 |
| Bacillus frigoritolerans | 5,475,560 | 43.9 | 1 | NZ_CP030063.1 |
3.3. Genomic Comparison
Marseille-P9898 strain genome was assembled from three contiguous sequences (N50, 4,279,115; L50, 1; coverage 12x) of 5,499,502 bp with a G+C content of 35.8% (Table 3). Marseille-P9898 strain presents a total of 5318 predicted protein-coding genes, 48 complete rRNAs, 114 tRNAs, and 1 tmRNA.
dDDH hybridization values were obtained using GGDC software and were reported in Table 3. Marseille-P9898 strain shows values ranged from 22.30% with Metab. niabensis to 34.20% with B. acidicola. These values were below the 70% threshold recognized for delineation of distinct species [32]. Ortho-ANI values ranged from 67.64% with Mesob. subterraneus to 85.76 % with Metab. halosaccharovorans, which was lower than the 95% threshold used to discriminate bacterial species [33] (Figure 4 and Table 4). These results were sufficient to separate the Marseille-P9898 strain from Metab. halosaccharovorans strain E33T, Metab. niabensis strain 4T19T, Metab. malikii strain NCCP-662T, Metab. litoralis strain SW-211T, and other most closely related species of genus Metabacillus. The distribution of genes into COG categories was similar among all compared genomes (Figure 5 and Table 5). Taken together, these results confirmed that the Marseille-P9898 strain belongs to a separate Bacillus species.
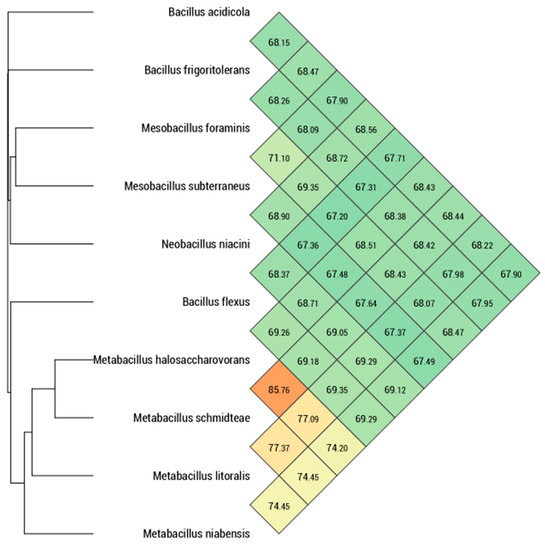
Figure 4.
The numeric map is generated with Ortho-ANI values calculated using OAT software between the Marseille-P9898 strain and related species with nomenclature classification. Color code shows closely (in red) to the farthest (in green) related species. Metabacillus halosaccharovorans, Metabacillus litoralis, and Metabacillus niabensis are closely related to Metabacillus schmidteae.
Table 4.
dDDH values obtained by comparison of related genomes studied using GGDC, formula 2 (DDH Estimates Based on Identities/HSP length). Taxa: 1, Bacillus acidicola; 2, Bacillus flexus 3, Mesobacillus foraminis; 4, Metabacillus halosaccharovorans; 5, Metabacillus litoralis; 6, Metabacillus niabensis; 7, Neobacillus niacini; 8, Metabacillus schmidteae; 9, Mesobacillus subterraneus.
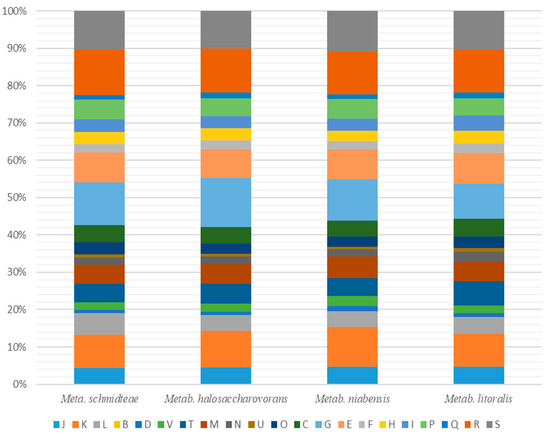
Figure 5.
Comparison of predicted genes according to the COGs of Marseille-P9898 strain and related species. Metabacillus halosaccharovorans, Metabacillus litoralis, and Metabacillus niabensis are closely related to Metabacillus schmidteae. [B] Chromatin structure and dynamics; [C] Energy production and conversion; [D] Cell cycle control, cell division, chromosome partitioning; [E] Amino acid transport and metabolism; [F] Nucleotide transport and metabolism; [G] Carbohydrate transport and metabolism; [H] Coenzyme transport and metabolism; [I] Lipid transport and metabolism; [J] Translation, ribosomal structure, and biogenesis; [K] Transcription; [L] Replication, recombination, and repair; [M] Cell wall/membrane/envelope biogenesis; [N] Cell motility; [O] Posttranslational modification, protein turnover, chaperones; [P] Inorganic ion transport and metabolism; [Q] Secondary metabolites biosynthesis, transport, and catabolism; [R] General function prediction only; [S] Function unknown; [T] Signal transduction mechanisms; [U] Intracellular trafficking, secretion, and vesicular transport; [V] Defense mechanisms; [W] Extracellular structures; [X] Mobilome: prophages, transposons; [Z] Cytoskeleton.
Table 5.
Functional annotation of predicted genes according to the COGs database.
3.4. Chemotaxonomic Analysis
The major fatty acids present in the Marseille-P9898 strain were 12-methyl-tetradecanoic acid (62.6%), 13-methyl-tetradecanoic acid (12.5%), Hexadecanoic acid (7.1%), 15-methyl-Hexadecanoic acid (3.8%), 12-methyl-Tridecanoic acid (3.3%), Pentadecanoic acid (2.7%), 12-methyl-Hexadecanoic acid (2.3%), 7-hexadecenoic acid (3.1%), 11-methyl-dodecanoic acid (1.1%), and 15-methyl-hexadecanoic acid (1.6%) (Table 6). 12-methyl-tetradecanoic acid was the most present in the Marseille-P9898 strain, Metab. halosaccharovorans, Metab. niabensis, Metab. malikii, and Metab. litoralis. Marseille-P9898 differed from Metab. halosaccharovorans, Metab. niabensis, Metab. malikii, and Metab. litoralis in Pentadecanoic acid and 11-methyl-Dodecanoic acid.
Table 6.
Cellular fatty acid composition of Marseille-P9898 compared with related species. Taxa: 1, Marseille-P9898; 2, Metabacillus niabensis 4T19T; 3, Metabacillus halosaccharovorans E33T; 4, Metabacillus litoralis SW-211T. The data of strain E33T and strain SW-211T are obtained using previously published work [48,50]. A of the fatty acid methyl esters was performed by gas-liquid chromatography according to the instructions for the Microbial Identification System (MIDI). Data of Marseille-P9898 and 4T19T strains were obtained in the present study. tr, Trace (<1 %); −, not detected; +, present; NA, data not available.
3.5. Antibiotic Susceptibility
Drugs such as amikacin, fosfomycin, benzylpenicillin, ciprofloxacin, amoxicillin, ceftriaxone, daptomycin, doxycycline, rifampicin, vancomycin, and ampicillin do not inhibit Marseille-P9898 strain growth (Table 7). Comparing Marseille-P9898 strain and Metab. niabensis, we found that fosfomycin inhibited the Marseille-P9898 strain, but not Metab. Niabensis growth.
Table 7.
Antimicrobial susceptibility and MIC values of Marseille-P9898 strain. CC: Tested range of drug concentration in µg/mL (microgram/milliliter). MIC: Minimum inhibition of concentration in µg/mL (microgram/milliliter).
4. Discussion
Planarian S mediterranea is an organism model to investigate the regeneration [72] and host-pathogen relationship [7,8,9]. To understand the implication of microbiota in the S. mediterranea antimicrobial response, we investigated its composition. While, recently, we have already published the identification of Pedobacter schmidteae sp. nov., as a novel bacterium isolated from this microbiota [73], here, we report the identification of a second novel bacteria species, which belongs to Metabacillus genus called Metabacillus schmidteae sp. nov.
Description of novel bacterial species requires several approaches, the most recurrent one being the taxonogenomic approach [1,2,3]. Using this approach, we describe the main phenotypic and genotypic features of the Marseille-P9898 strain. The use of MALDI-TOF-MS showed a protein profile, but it did not allow the discrimination between the Marseille-P9898 strain and Metabacillus niabensis. 16S rRNA gene sequence presented a value of more than 98.65% [29], a cut-off to demarcate new species. We know that the conventional low divergence between two 16S rRNA genes from two organisms results in a slight and limited bacterial description [74,75]. Nonetheless, gene 16S rRNA sequence gives us the information concerning genomic sequence analysis by tools such as Genome-to-Genome Distance Calculator (GGDC) [32] and Orthologous Average Nucleotide Identity (Ortho-ANI). Genomics allows the evaluation of the degrees of genomic similarity among species [33]. Taken together, genomic data showed that Marseille-P9898 is a novel bacterial species of genus Metabacillus called Metabacillus schmidteae sp. nov.
Until now, Metabacillus have been isolated from diverse environments including soil, hyper-saline aquatic, and marine coastal region [11]. We revealed that Metabacillus schmidteae is also hosted by aquatic animals such as planarians, precisely in their microbiota. We have observed that Metabacillus schmidteae is a Gram-negative bacteria (fresh and older cultures) and it is the first Gram-negative one from Metabacillus genus, which is usually Gram-positive or Gram-variable stained [11]. It possesses a polar motile flagellum, and it is strictly aerobic as other species of the genus Metabacillus. Metabacillus schmidteae can make spores under harsh conditions as reported for other species, except for Metabacillus weihaiensis [11]. In addition, biochemical analysis shows that Metabacillus (Metabacillus schmidteae Marseille-P9898T, Metabacillus niabensis 4T19T, Metabacillus halosaccharovorans E33T, and Metabacillus litoralis SW-211T) use as substrates for their growth, D-galactose, D-glucose, D-fructose, D-cellobiose, D-maltose, D-lactose, D-melibiose, D-saccharose, D-trehalose, and D-raffinose. Chemotaxonomic analysis indicates that Metabacillus (Metabacillus schmidteae Marseille-P9898T, Metabacillus niabensis 4T19T, Metabacillus halosaccharovorans E33T, and Metabacillus litoralis SW-211T) produce membrane proteins such 15:0 iso (13-methyl-tetradecanoic acid), 15:0 anteiso (12-methyl-tetradecanoic acid), 17:0 anteiso (15-methyl-Hexadecanoic acid). The data of chemotaxonomic and biochemical analysis could be used as a protein or biochemical marker to identify the genus metabacillus. Metabacillus schmidteae is now the second bacteria newly identified in planarian microbiota [73]. Its contribution in regeneration, as well as the antimicrobial capacity of the planarian, remains to be investigated.
5. Conclusions
Using taxonogenomic analysis, we described and characterized the Marseille-P9898 strain, hereby for the first time isolated from planarian S. schmidteae microbiota. Marseille-P9898 is phylogenetically related to Metabacillus genus. Chemotaxonomic and biochemical analysis and genomics comparison allowed considering Marseille-P9898 as a novel species of Metabacillus genus. For this strain Marseille-P9898, we propose the name of Metabacillus schmidteae sp. nov. (Marseille-P9898T = CSUR P9898T = DSM 111480T).
5.1. Protologue
Metabacillus schmidteae (schmid.te’ae. N.L. gen. n. schmidteae of the planarian genus Schmidtea, from which Marseille-P9898 strain was isolated) is a bacterium belonging to the Bacillaceae family within Firmicutes phylum. Marseille-P9898T type strain is strictly aerobic, alkaliphilic, Gram-negative, spore-forming, rod-shaped, and motile. After 2 days at 28 °C on Columbia agar at pH 7.5, colonies were small, circular, smooth, white, and convex. Growth occurs at 19–50 °C with 0–12 g/L NaCl and pH 7.5–10. Optimal growth occurs at 28 °C, pH 8, and 10 g/L NaCl. Catalase positive and oxidase negative. Reactions were positive for alkaline phosphatase, esterase (C4), esterase lipase (C8), Naphthol-AS-BI-phosphohydrolase, β-glucuronidase, and α-glucosidase; assimilate to glycerol, L-arabinose, D-xylose, methyl-βD-xylopyranoside, D-galactose, D-glucose, D-fructose, D-mannose, D-mannitol, methyl-αD-glucopyranoside, N-acetylglucosamine, amygdalin, esculin ferric citrate, salicin, D-cellobiose, D-maltose, D-lactose, D-melibiose, D-saccharose, D-trehalose, inulin, D-melezitose, gentiobiose, D-lyxose, D-raffinose, starch, D-arabitol, 2-nitrophenyl-βD-galactopyranoside, L-tryptophan, natriumpyruvat, 4-Nitrophenyl-βD-Galactopyranoside, malic acid, and trisodium citrate. Negative reactions were observed for lipase (C14), leucine arylamidase, valine arylamidase, cystine arylamidase, trypsin, α-chymotrypsin, acid phosphatase, α-galactosidase, β-galactosidase, β-glucosidase, and N-acetyl-β-glucosaminidase; no assimilate to α-mannosidase, α-fucosidase, erythritol, D-arabinose, D-ribose, L-xylose, D-adonitol, L-sorbose, L-rhamnose, Dulcitol, Inositol, D-sorbitol, methyl-αD-mannopyranoside, arbutin, glycogen, xylitol, D-turanose, D-tagatose, D-fucose, L-fucose, L-arabitol, potassium gluconate, potassium 2-ketogluconate, potassium 5-ketogluconate, L-arginin, L-lysin, L-ormithin, trinatriumcitrat, natriumthiosulfat, urea, indole production, gelatin, potassium nitrate, capric acid, adipic acid, and phenylacetic acid. Marseille-P9898T strain is sensitive to amikacin, fosfomycin, benzylpenicillin, ciprofloxacin, amoxicillin, ceftriaxone, daptomycin, doxycycline, rifampicin, vancomycin, and ampicillin. Fatty acids were 12-methyl-tetradecanoic acid (62.6%), 13-methyl-tetradecanoic acid (12.5%), Hexadecanoic acid (7.1%), 15-methyl-Hexadecanoic acid (3.8%), and 12-methyl-Tridecanoic acid (3.3%). Marseille-P9898T type strain (CSUR P9898T = DSM 111480) was isolated from planarian microbiota. The 16S rRNA gene sequence and genome sequence were deposited in GenBank under accession numbers LR797940 and CAESCH000000000.1, respectively.
5.2. Nucleotide Sequence Accession Number
The 16S rRNA gene sequence and genome sequence were deposited in GenBank under accession numbers LR797940 and CAESCH000000000.1, respectively. Raw data of Illumina MiSeq paired-end and MinION sequencing were deposited in EMBL-EBI under run accession ERR4143811 and ERR4143810; and experiment Accession ERX4111084 and ERX4111083, respectively.
5.3. Deposit in Culture Collections
Marseille-P9898T strain was deposited in the CSUR and DSMZ strain collections under numbers CSUR P9898 and DSM 111480, respectively.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/microbiolres12020021/s1, Figure S1: Colonies color Colonies color of Metabacillus schmidteae, Figure S2: Electron Micrograph of Metabacillus schmidteae Spore, Figure S3: MALDI-TOF spectrum of Metabacillus schmidteae, Table S1: GenBank accession numbers of the type strains.
Author Contributions
L.J.K. conceived the experiments, realised the experiments, analysed the data, prepared figures and drafted the manuscript. D.A.R., E.G. and P.-E.F. designed the experiments, conceived the experiments, analysed the data, drafted and finalised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the Méditerranée-Infection foundation, the National Research Agency under the program “Investissements d’avenir”, reference ANR-10-IAHU-03 and by Région Provence Alpes Côte d’Azur and European funding FEDER IHUBIOTK.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The 16S rRNA gene sequence and genome sequence were deposited in GenBank under accession numbers LR797940 and CAESCH000000000.1, respectively. Raw data of Illumina MiSeq paired-end and MinION sequencing were deposited in EMBL-EBI under run accession ERR4143811 and ERR4143810; and experiment Accession ERX4111084 and ERX4111083, respectively.
Acknowledgments
LJK is a student at the Aix-Marseille University and funded by the Méditerranée-Infection foundation. The study was funded by the Méditerranée-Infection foundation, the National Research Agency under the program “Investissements d’avenir”, reference ANR-10-IAHU-03 and by Région Provence Alpes Côte d’Azur and European funding FEDER IHUBIOTK. We also thank Aurelia Caputo (IHU Méditerranée-Infection, France) for submitting the raw data assembly, 16S rRNA and genomic sequences to GenBank and Nicholas Armstrong (IHU Méditerranée-Infection, France) for cellular fatty acid methyl ester analysis. We thank Giovanna Mottola (C2VN-AMU-INSERM 1263-INRAE 1260 and AP-HM, Marseille, France) for her help in English editing.
Conflicts of Interest
The authors have no conflicts of interest to declare. The funding sources had no role in the study design, data collection and analysis, decision to publish, or manuscript preparation.
Abbreviations
| DSMZ | Deutsche Sammlung von Mikroorganismen und Zellkulturen |
| CSUR | Collection de Souches de l′Unité des Rickettsies |
| dDDH | digital DNA-DNA hybridization |
| GGDC | Genome-to-Genome Distance Calculator |
| MALDI-TOF-MS | Matrix-Assisted Laser Desorption Ionization Time-Of-Flight Mass Spectrometry |
| MEGA | Molecular Evolutionary Genetics Analysis |
| FAME | Fatty Acid Methyl Ester |
References
- Ramasamy, D.; Mishra, A.K.; Lagier, J.-C.; Padhmanabhan, R.; Rossi, M.; Sentausa, E.; Raoult, D.; Fournier, P.-E. A Polyphasic strategy incorporating genomic data for the taxonomic description of novel bacterial species. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 2, 384–391. [Google Scholar] [CrossRef]
- Fournier, P.-E.; Lagier, J.-C.; Dubourg, G.; Raoult, D. From Culturomics to Taxonomogenomics: A need to change the taxonomy of prokaryotes in clinical microbiology. Anaerobe 2015, 36, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Morel, A.-S.; Dubourg, G.; Prudent, E.; Edouard, S.; Gouriet, F.; Casalta, J.-P.; Fenollar, F.; Fournier, P.E.; Drancourt, M.; Raoult, D. Complementarity between targeted real-time specific PCR and conventional Broad-Range 16S rDNA PCR in the syndrome-driven diagnosis of infectious diseases. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Rosselló-Móra, R.; Busse, H.J.; Ludwig, W.; Kämpfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. Int. J. Syst. Evol. Microbiol. 2010, 60 Pt 1, 249–266. [Google Scholar] [CrossRef]
- Drancourt, M.; Berger, P.; Raoult, D. Systematic 16S rRNA gene sequencing of atypical clinical isolates identified 27 new bacterial species associated with Humans. J. Clin. Microbiol. 2004, 42, 2197–2202. [Google Scholar] [CrossRef] [PubMed]
- Wayne, L.G. International committee on systematic bacteriology: Announcement of the report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Zent. Bakteriol. Mikrobiol. Hyg. Ser. A Med. Microbiol. Infect. Dis. Virol. Parasitol. 1988, 268, 433–434. [Google Scholar] [CrossRef]
- Abnave, P.; Mottola, G.; Gimenez, G.; Boucherit, N.; Trouplin, V.; Torre, C.; Conti, F.; Ben Amara, A.; Lepolard, C.; Djian, B.; et al. Screening in planarians identifies Morn2 as a key component in lc3-associated phagocytosis and resistance to bacterial infection. Cell Host. Microbe 2014, 16, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Maciel, E.I.; Jiang, C.; Barghouth, P.G.; Nobile, C.J.; Oviedo, N.J. The planarian schmidtea mediterranea is a new model to study host-pathogen interactions during fungal infections. Dev. Comp. Immunol. 2019, 93, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Torre, C.; Ghigo, É. Planaria: An immortal worm to clarify human immune response. Med. Sci. 2015, 31, 20–22. [Google Scholar] [CrossRef][Green Version]
- Seng, P.; Abat, C.; Rolain, J.M.; Colson, P.; Lagier, J.-C.; Gouriet, F.; Fournier, P.E.; Drancourt, M.; La Scola, B.; Raoult, D. Identification of rare pathogenic bacteria in a clinical microbiology laboratory: Impact of Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2013, 51, 2182–2194. [Google Scholar] [CrossRef]
- Patel, S.; Gupta, R.S. A phylogenomic and comparative genomic framework for resolving the polyphyly of the genus Bacillus: Proposal for six new genera of Bacillus species, Peribacillus gen. nov., Cytobacillus gen. nov., Mesobacillus gen. nov., Neobacillus gen. nov., Metabacillus gen. nov. and Alkalihalobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 406–438. [Google Scholar] [CrossRef] [PubMed]
- Fritze, D. Taxonomy of the Genus Bacillus and Related Genera: The aerobic endospore-forming bacteria. Phytopathology 2004, 94, 1245–1248. [Google Scholar] [CrossRef] [PubMed]
- Cohn, F. Untersuchungen über Bacterien. Beiträge Biologie Pflanzen 1872, 1, 127–224. [Google Scholar]
- Fritze, D.; Pukall, R. Reclassification of bioindicator strains Bacillus subtilis DSM 675 and Bacillus subtilis DSM 2277 as Bacillus Atrophaeus. Int. J. Syst. Evol. Microbiol. 2001, 51, 35–37. [Google Scholar] [CrossRef]
- Fischer, A. Untersuchungen über bakterien. Jahrbücher Wissenschaftliche Botanik 1895, 27, 1–163. [Google Scholar]
- GIBBONS, N.E.; MURRAY, R.G.E. Proposals concerning the higher taxa of bacteria. Int. J. Syst. Evol. Microbiol. 1978, 28, 1–6. [Google Scholar] [CrossRef]
- Arahal, D.R.; Ventosa, A. Moderately halophilic and halotolerant species of bacillus and related genera. In Applications and Systematics of Bacillus and Relatives; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; pp. 83–99. [Google Scholar] [CrossRef]
- Márquez, M.C.; Sánchez-Porro, C.; Ventosa, A. halophilic and haloalkaliphilic, aerobic endospore-forming bacteria in soil. In Endospore-Forming Soil Bacteria; Logan, N.A., Vos, P., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 309–339. [Google Scholar] [CrossRef]
- Ventosa, A. Unusual micro-organisms from unusual habitats: Hypersaline environments. In Prokaryotic Diversity: Mechanisms and Significance; Lappin-Scott, H.M., Logan, N.A., Oyston, P.C.F., Eds.; Society for General Microbiology Symposia; Cambridge University Press: Cambridge, UK, 2006; pp. 223–254. [Google Scholar] [CrossRef]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of Moderately Halophilic Aerobic Bacteria. Microbiol Mol. Biol. Rev 1998, 62, 504–544. [Google Scholar] [CrossRef]
- De la Haba, R.R.; Sánchez-Porro, C.; Marquez, M.C.; Ventosa, A. Taxonomy of Halophiles. In Extremophiles Handbook; Horikoshi, K., Ed.; Springer: Tokyo, Japan, 2011; pp. 255–308. [Google Scholar] [CrossRef]
- Seng, P.; Drancourt, M.; Gouriet, F.; La Scola, B.; Fournier, P.-E.; Rolain, J.M.; Raoult, D. Ongoing Revolution in Bacteriology: Routine Identification of Bacteria by Matrix-Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry. Clin. Infect. Dis. 2009, 49, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Ravi, R.K.; Walton, K.; Khosroheidari, M. MiSeq: A Next Generation Sequencing platform for genomic analysis. Methods Mol. Biol. 2018, 1706, 223–232. [Google Scholar] [CrossRef]
- Karamitros, T.; Magiorkinis, G. Multiplexed targeted sequencing for Oxford Nanopore MinION: A detailed library preparation procedure. Methods Mol. Biol. 2018, 1712, 43–51. [Google Scholar] [CrossRef]
- Jain, M.; Olsen, H.E.; Paten, B.; Akeson, M. The Oxford Nanopore MinION: Delivery of Nanopore sequencing to the genomics community. Genome Biol. 2016, 17. [Google Scholar] [CrossRef]
- Wingett, S.W.; Andrews, S. FastQ Screen: A tool for multi-genome mapping and quality control. F1000Res 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M.; Spröer, C.; Klenk, H.-P. When should a DDH experiment be mandatory in microbial taxonomy? Arch. Microbiol. 2013, 195, 413–418. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial dna in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Auch, A.F.; von Jan, M.; Klenk, H.-P.; Göker, M. Digital DNA-DNA Hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Ouk Kim, Y.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Unaogu, I.C.; Gugnani, H.C.; Boiron, P. The enzymatic profile of some pathogenic aerobic actinomycetes as determined by api-zym method. J. Med. Mycol. 2008, 9, 235. [Google Scholar]
- Gruner, E.; von Graevenitz, A.; Altwegg, M. The API ZYM System: A Tabulated Review from 1977 to Date. J. Microbiol. Methods 1992, 16, 101–118. [Google Scholar] [CrossRef]
- Humble, M.W.; King, A.; Phillips, I. API ZYM: A Simple rapid system for the detection of bacterial enzymes. J. Clin. Pathol. 1977, 30, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Søgaard, P.; Gahrn-Hansen, B.; Zhou, H.P.; Frederiksen, W. An Investigation of three commercial methods for rapid identification of non-enteric gram-negative rods. application on pseudomonas paucimobilis and some other pseudomonas species. Acta Pathol. Microbiol. Immunol. Scand. B 1986, 94, 357–363. [Google Scholar] [CrossRef] [PubMed]
- MK, B.; DA, B.; GL, C.; JG, G. Comparison of five commercial methods for the identification of non- fermentative and oxidase positive fermentative gram-negative Bacilli. N. Z. J. Med. Lab. Technol. 1988, 42, 8–12. [Google Scholar]
- Swanson, E.C.; Collins, M.T. Use of the API 20E System to identify veterinary enterobacteriaceae. J. Clin. Microbiol. 1980, 12, 10–14. [Google Scholar] [CrossRef]
- Smith, P.B.; Tomfohrde, K.M.; Rhoden, D.L.; Balows, A. API System: A multitube micromethod for identification of enterobacteriaceae. Appl. Microbiol. 1972, 24, 449–452. [Google Scholar] [CrossRef]
- Véron, M.; Le Minor, L. Nutrition and taxonomy of “enterobacteriaceae” and related bacteria. III. Nutritional characters and differentiation of the taxonomic groups. Ann. Microbiol. 1975, 126, 125–147. [Google Scholar]
- Bergey, D.H.; Krieg, N.R.; Holt, J.G. Bergey’s Manual of Systematic Bacteriology; Williams & Wilkins: Baltimore, MD, USA, 1984. [Google Scholar]
- Rogosa, M.; Sharpe, M.E. An Approach to the Classification of the Lactobacilli. J. Appl. Bacteriol. 1960, 22, 329–340. [Google Scholar]
- Sharpe, M.E.; Hill, L.R.; Lapage, S.P. Pathogenic Lactobacilli. J. Med. Microbiol. 1973, 6, 281–286. [Google Scholar] [CrossRef]
- Kwon, S.-W.; Lee, S.-Y.; Kim, B.-Y.; Weon, H.-Y.; Kim, J.-B.; Go, S.-J.; Lee, G.-B. Bacillus niabensis sp. nov., isolated from cotton-waste composts for mushroom cultivation. Int. J. Syst. Evol. Microbiol. 2007, 57, 1909–1913. [Google Scholar] [CrossRef] [PubMed]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids. MIDI 1990, 20, 1–6. [Google Scholar]
- Dione, N.; Sankar, S.A.; Lagier, J.-C.; Khelaifia, S.; Michele, C.; Armstrong, N.; Richez, M.; Abrahão, J.; Raoult, D.; Fournier, P.-E. Genome Sequence and Description of Anaerosalibacter massiliensis Sp. Nov. New Microbes New Infect. 2016, 10, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Mehrshad, M.; Amoozegar, M.A.; Didari, M.; Bagheri, M.; Fazeli, S.A.S.; Schumann, P.; Spröer, C.; Sánchez-Porro, C.; Ventosa, A. Bacillus halosaccharovorans sp. nov., a moderately halophilic bacterium from a hypersaline lake. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 8, 2776–2781. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Ahmed, I.; Kudo, T.; Iqbal, M.; Lee, Y.-J.; Fujiwara, T.; Ohkuma, M. A heavy metal tolerant novel bacterium, Bacillus malikii sp. nov., isolated from tannery effluent wastewater. Antonie Van Leeuwenhoek 2015, 108, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Oh, T.-K. Bacillus litoralis sp. nov., isolated from a tidal flat of the yellow sea in korea. Int. J. Syst. Evol. Microbiol. 2005, 55 Pt 5, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Delaporte, B.; Sasson, A. Study of bacteria from arid soils of Morocco: Brevibacterium haloterans n. sp. and Brevibacterium frigoritolerans n. sp. Comptes Rendus Hebd. Seances Acad. Sci. Ser. D Sci. Nat. 1967, 264, 2257–2260. [Google Scholar]
- Liu, G.H.; Liu, B.; Wang, J.P.; Che, J.M.; Li, P.F. Reclassification of Brevibacterium frigoritolerans DSM 8801T as Bacillus frigoritolerans comb. nov. Based on Genome Analysis. Curr. Microbiol. 2020, 77, 1916–1923. [Google Scholar] [CrossRef]
- Tiago, I.; Pires, C.; Mendes, V.; Morais, P.V.; da Costa, M.S.; Veríssimo, A. Bacillus foraminis sp. nov., isolated from a non-saline alkaline groundwater. Int. J. Syst. Evol. Microbiol. 2006, 56 Pt 11, 2571. [Google Scholar] [CrossRef]
- Kanso, S.; Greene, A.C.; Patel, B.K.C. Bacillus subterraneus sp. nov., an iron- and manganese-reducing bacterium from a deep subsurface australian thermal aquifer. Int. J. Syst. Evol. Microbiol. 2002, 52 Pt 3, 869. [Google Scholar] [CrossRef]
- Wieser, M.; Worliczek, H.; Kämpfer, P.; Busse, H.-J. Bacillus herbersteinensis sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55 Pt 5, 2119–2123. [Google Scholar] [CrossRef]
- NAGEL, M.; ANDREESEN, J.R. Bacillus niacini sp. nov., a nicotinate-metabolizing mesophile isolated from soil. Int. J. Syst. Evol. Microbiol. 1991, 41, 134–139. [Google Scholar] [CrossRef][Green Version]
- Albert, R.A.; Archambault, J.; Rosselló-Mora, R.; Tindall, B.J.; Matheny, M. Bacillus acidicola sp. nov., a novel mesophilic, acidophilic species isolated from acidic sphagnum peat bogs in wisconsin. Int. J. Syst. Evol. Microbiol. 2005, 55 Pt 5, 2125–2130. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Gao, J.-S.; Zhang, L.; Zhang, C.-W.; Ma, X.-T.; Zhang, J. Bacillus oryzisoli sp. nov., isolated from rice rhizosphere. Int. J. Syst. Evol. Microbiol. 2016, 66, 3432–3436. [Google Scholar] [CrossRef]
- Priest, F.G.; Goodfellow, M.; Todd, C. A numerical classification of the genus bacillus. J. Gen. Microbiol. 1988, 134, 1847–1882. [Google Scholar] [CrossRef]
- Suresh, K.; Prabagaran, S.R.; Sengupta, S.; Shivaji, S. Bacillus indicus sp. nov., an arsenic-resistant bacterium isolated from an aquifer in west bengal, india. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 4, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Seiler, H.; Wenning, M.; Schmidt, V.; Scherer, S. Bacillus gottheilii sp. nov., isolated from a pharmaceutical manufacturing site. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 3, 867–872. [Google Scholar] [CrossRef]
- Nielsen, P.; Fritze, D.; Priest, F.G. Phenetic diversity of alkaliphilic bacillus strains: Proposal for nine new species. Microbiology 1995, 141, 1745–1761. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, D.-F.; Khieu, T.-N.; Son, C.K.; Zhang, X.-M.; Cheng, J.; Tian, X.-P.; Zhang, S.; Li, W.-J. Bacillus tianshenii sp. nov., isolated from a marine sediment sample. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 6, 1998–2002. [Google Scholar] [CrossRef]
- Shivaji, S.; Chaturvedi, P.; Begum, Z.; Pindi, P.K.; Manorama, R.; Padmanaban, D.A.; Shouche, Y.S.; Pawar, S.; Vaishampayan, P.; Dutt, C.B.S.; et al. Janibacter hoylei sp. nov., Bacillus isronensis sp. nov. and Bacillus aryabhattai sp. nov., isolated from cryotubes used for collecting air from the upper atmosphere. Int. J. Syst. Evol. Microbiol. 2009, 59 Pt 12, 2977–2986. [Google Scholar] [CrossRef]
- Parag, B.; Sasikala, C.; Ramana, C.V. Bacillus endolithicus sp. nov., isolated from pebbles. Int. J. Syst. Evol. Microbiol. 2015, 65, 4568–4573. [Google Scholar] [CrossRef]
- Heyrman, J.; Vanparys, B.; Logan, N.A.; Balcaen, A.; Rodríguez-Díaz, M.; Felske, A.; De Vos, P. Bacillus novalis sp. nov., Bacillus vireti sp. nov., Bacillus soli sp. nov., Bacillus bataviensis sp. nov. and Bacillus drentensis sp. nov., from the drentse a grassland. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 1, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Balcázar, J.L.; Pintado, J.; Planas, M. Bacillus galliciensis sp. nov., isolated from faeces of wild seahorses (Hippocampus Guttulatus). Int. J. Syst. Evol. Microbiol. 2010, 60 Pt 4, 892–895. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-H.; Tian, X.-R.; Ruan, Y.; Yang, L.-L.; He, Z.-Q.; Tang, S.-K.; Li, W.-J.; Shi, H.; Chen, Y.-G. Bacillus crassostreae sp. nov., isolated from an oyster (Crassostrea Hongkongensis). Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 5, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a Natural System of Organisms: Proposal for the domains archaea, bacteria, and eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef]
- Ludwig, W.; Schleifer, K.-H.; Whitman, W.B. Bacilli class. Nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; American Cancer Society: Atlanta, GA, USA, 2015. [Google Scholar] [CrossRef]
- Vos, P.D. Bacillales. In Bergey’s Manual of Systematics of Archaea and Bacteria; American Cancer Society: Atlanta, GA, USA, 2015. [Google Scholar] [CrossRef]
- Reddien, P.W. The Cellular and Molecular Basis for Planarian Regeneration. Cell 2018, 175, 327–345. [Google Scholar] [CrossRef]
- Kangale, L.J.; Raoult, D.; Ghigo, E.; Fournier, P.E. Pedobacter schmidteae sp. nov., a new bacterium isolated from the microbiota of the planarian Schmidtea mediterranea. Sci. Rep. 2020, 10, 6113. [Google Scholar] [CrossRef]
- Pei, A.Y.; Oberdorf, W.E.; Nossa, C.W.; Agarwal, A.; Chokshi, P.; Gerz, E.A.; Jin, Z.; Lee, P.; Yang, L.; Poles, M.; et al. Diversity of 16S rRNA genes within individual prokaryotic genomes. Appl. Environ. Microbiol. 2010, 76, 3886–3897. [Google Scholar] [CrossRef]
- Ochman, H.; Elwyn, S.; Moran, N.A. Calibrating bacterial evolution. Proc. Natl. Acad. Sci. USA 1999, 96, 12638–12643. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).