
La Crosse Virus Circulation in Virginia, Assessed via Serosurveillance in Wildlife Species



Abstract
1. Introduction
2. Materials and Methods
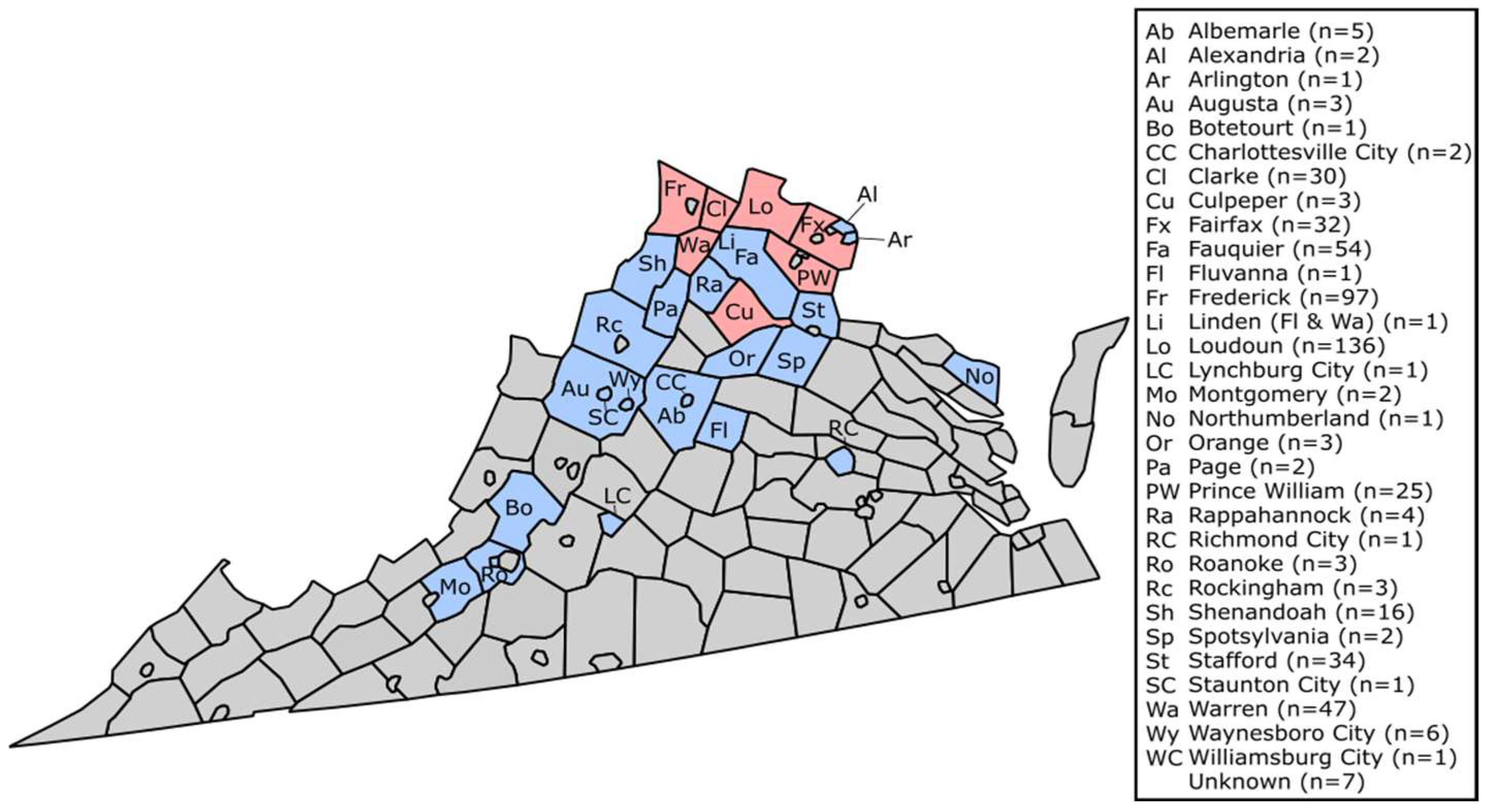
2.1. Sample Collection
2.2. Power Analysis
2.3. Serological Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weaver, S.C.; Reisen, W.K. Present and Future Arboviral Threats. Antiviral. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Mordecai, E.A.; Ryan, S.J.; Cldwell, J.M.; Shah, M.M.; LaBeaud, A.D. Climate change could shift disease burden from malaria to arboviruses in Africa. Lancet Planet. Health 2020, 4, e416–e423. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Gao, X.; Gould, E.A. Factors responsible for the emergence of arboviruses: Strategies, challenges and limitations for their control. Emer. Micro. Infect. 2015, 4, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Fischer, M.; Dominguez, S.R.; Tyler, K.L.; Abzug, M.J. Encephalitis in Children. Infect. Dis. Clin. North Am. 2018, 32, 145–162. [Google Scholar] [CrossRef]
- Hughes, H.R.; Adkins, S.; Alkhovshiy, A.; Beer, M.; Blair, C.; Calisher, C.H.; Drebot, M.; Lambert, A.J.; de Souza, W.M.; Marklewitz; et al. ICTV Virus Taxonomy Profile: Peribunyaviridae. J. Gen. Virol. 2020, 101, 1–2. [Google Scholar] [CrossRef]
- Thompson, W.H.; Kalfayan, B.; Anslow, R.O. Isolation of California encephalitis group virus from a fatal human illness. Am. J. Epidemiol. 1965, 81, 245–253. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. “La Crosse Encephalitis—Statistics & Maps”; US Department of Health and Human Services: Atlanta, GA, USA, 2022. [Google Scholar]
- McJunkin, J.E.; de los Reyes, E.C.; Irazuzta, J.E.; Caceres, M.J.; Khan, R.R.; Minnich, L.L.; Fu, F.D.; Lovett, G.D.; Tsai, T.; Thompson, A. La Crosse Encephalitis in Children. N. Eng. J. Med. 2001, 344, 801–807. [Google Scholar] [CrossRef]
- Gaensbauer, J.T.; Lindsey, N.P.; Messacar, K.; Staples, J.E.; Fischer, M. Neuroinvasive Arboviral Disease in the United States: 2003 to 2012. Pediatrics 2014, 134, e642–e650. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. “La Crosse Encephalitis—Virus”; US Department of Health and Human Services: Atlanta, GA, USA, 2022. [Google Scholar]
- Balfour Jr., H.H.; Siem, R.A.; Bauer, H.; Quie, P.G. California Arbovirus (La Crosse) Infections I. Clinical and laboratory findings in 66 children with meningoencephalitis. Pediatrics 1973, 52, 680–691. [Google Scholar]
- Calisher, C.H. Medically important arboviruses of the United States and Canada. Clin. Microbiol. Rev. 1994, 7, 89–116. [Google Scholar] [CrossRef]
- Jones, T.F.; Craig, A.S.; Nasci, R.S.; Patterson, L.E.R.; Erwin, P.C.; Gerhardt, R.R.; Ussery, X.T.; Schaffner, W. Newly Recognized Focus of La Crosse Encephalitis in Tennessee. Clin. Infect. Dis. 1999, 28, 93–97. [Google Scholar] [CrossRef]
- Bewick, S.; Agusto, F.; Calabrese, J.M.; Muturi, E.J.; Fagan, W.F. Epidemiology of La Crosse Virus Emergence, Appalachia Region, United States. Emerg. Infect. Dis. 2016, 22, 1921–1929. [Google Scholar] [CrossRef]
- Leisnham, P.; Juliano, S. Impacts of climate, land use, and biological invasion on the ecology of immature Aedes mosquitoes: Implications for La Crosse emergence. EcoHealth 2012, 9, 217–228. [Google Scholar] [CrossRef]
- Nasci, R.S.; Moore, C.G.; Biggerstaff, B.J.; Panella, N.A.; Liu, H.Q.; Karabatsos, N.; Davis, B.S.; Brannon, E.S. La Crosse Encephalitis Virus Habitat Associations in Nicholas County, West Virginia. J. Med. Entomol. 2000, 37, 559–570. [Google Scholar] [CrossRef]
- Haddow, A.D.; Odoi, A. The Incidence Risk, Clustering, and Clinical Presentation of La Crosse Virus Infections in the Eastern United States, 2003-2007. PLoS ONE 2009, 4, e6145. [Google Scholar] [CrossRef]
- Kitron, U.; Swanson, J.; Crandell, M.; Sullivan, P.J.; Anderson, J.; Garro, R.; Haramis, L.D.; Grimstad, P.R. Introduction of Aedes albopictus into a La Crosse virus—Enzootic site in Illinois. Emerg. Infect. Dis. 1998, 4, 627–630. [Google Scholar] [CrossRef]
- Grimstad, P.R.; Kobayashi, J.F.; Zhang, M.B.; Craig, G.B. Recently introduced Aedes albopictus in the United States: Potential vector of La Crosse virus (Bunyaviridae: California serogroup). J. Am. Mosq. Control Assoc. 1989, 5, 422–427. [Google Scholar]
- Gerhardt, R.R.; Gottfried, K.L.; Apperson, C.S.; Davis, B.S.; Erwin, P.C.; Smith, A.B.; Panella, N.A.; Powell, E.E.; Nasci, R.S. First isolation of La Crosse virus from naturally infected Aedes albopictus. Emerg. Infect. Dis. 2001, 7, 807–811. [Google Scholar] [CrossRef]
- Bevins, S.N. Establishment and Abundance of a Recently Introduced Mosquito Species Ochlerotatus japonicus (Diptera: Culicidae) in the Southern Appalachians, USA. J. Med. Entomol. 2007, 44, 945–952. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J.; Andre, R.G. Laboratory Transmission of La Crosse Virus by Ochlerotatus j. japonicus (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Vahey, G.M.; Lindsey, N.P.; Stapes, J.E.; Hills, S.L. La Crosse Virus Disease in the United States, 2003–2019. Am. J. Trop. Med. Hyg. 2021, 105, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. ArboNET Disease Maps; US Department of Health and Human Services: Atlanta, GA, USA, 2023. [Google Scholar]
- Graham, S.P.; Hassan, H.K.; Chapman, T.; White, G.; Guyer, C.; Unnasch, T.R. Serosurveillance of Eastern Equine Encephalitis Virus in Amphibians and Reptiles from Alabama, USA. Am. J. Trop. Med. Hyg. 2012, 86, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.P.; Tennant, R.A.; Kozak III, J.A.; Cope, E.H.; Walsh, J.D. Trapping and serosurveillance of live wild birds for arboviruses. TBFMCA 2008, 8, 10–14. [Google Scholar]
- Eastwood, G.; Sang, R.C.; Guerbois, M.; Taracha, E.L.N.; Weaver, S.C. Enzootic Circulation of Chikungunya Virus in East Africa: Serological Evidence for Non-human Kenyan Primates. Am. J. Trop. Med. Hyg. 2017, 97, 1399–1404. [Google Scholar] [CrossRef]
- Eastwood, G.; Goodman, S.J.; Hilgery, N.; Cruz, M.; Kramer, L.D.; Cunningham, A.A. Using Avian Surveillance in Ecuador to Assess the Imminence of West Nile Virus Incursion to Galápagos. EcoHealth 2014, 11, 53–62. [Google Scholar] [CrossRef]
- Boromisa, R.D.; Grimstad, P.R. Seroconversion rates to Jamestown Canyon virus among six populations of white-tailed deer (Odocoileus virginianus) in Indiana. J. Wildl. Dis. 1987, 23, 23–33. [Google Scholar] [CrossRef]
- Neitzel, D.F.; Grimstad, P.R. Serological evidence of California group and Cache Valley virus infection in Minnesota white-tailed deer. J. Wildl. Dis. 1991, 27, 230–237. [Google Scholar] [CrossRef]
- Dohoo, I.R.; Martin, S.W.; Stryhn, H. Veterinary Epidemiologic Research, 2nd ed.; AVC Incorporated: Charlottetown, PE, Canada, 2003. [Google Scholar]
- Hopkins, M.C.; Zink, S.D.; Paulson, S.L.; Hawley, D.M. Influence of Forest Disturbance on La Crosse Virus Risk in Southwestern Virginia. Insects 2019, 11, 28. [Google Scholar] [CrossRef]
- Moulton, D.W.; Thompson, W.H. California Group Virus Infections in Small, Forest-Dwelling Mammals of Wisconsin: Some Ecological Considerations. Am. J. Trop. Med. Hyg. 1971, 20, 474–482. [Google Scholar] [CrossRef]
- Amundson, T.E.; Yuill, T.M.; DeFoliart, G.R. Experimental La Crosse Virus Infection of Red Fox (Vulpes fulva), Raccoon (Procyon lotor), Opossum (Didelphis virginiana), and Woodchuck (Marmota monax). Am. J. Trop. Med. Hyg. 1985, 34, 586–595. [Google Scholar] [CrossRef]
- Cully, J.; Heard, P.; Wesson, D.; Craig, G. Antibodies to La Crosse virus in eastern chipmunks in Indiana near an Aedes albopictus population. J. Am. Mosq. Control. Assoc. 1992, 7, 651–653. [Google Scholar]
- Gauld, L.W.; Yuill, T.M.; Hanson, R.P.; Sinha, S.K. Isolation of La Crosse Virus (California Encephalitis Group) from Chipmunk (Tamias striatus), an Amplifier Host. Am. J. Trop. Med. Hyg. 1975, 24, 999–1005. [Google Scholar] [CrossRef]
- Dressler, R.L.; Ganaway, J.R.; Storm, G.L.; Tzilkowski, W.M. Serum antibody prevalence for Herpesvirus sylvilagus, Bacillus piliformis and California serogroup arboviruses in cottontail rabbits from Pennsylvania. J. Wildl. Dis. 1988, 24, 352–355. [Google Scholar] [CrossRef]
- Calisher, C.H.; Pretzman, C.I.; Muth, D.J.; Parsons, M.A.; Peterson, E.D. Serodiagnosis of La Crosse virus infections in humans by detection of immunoglobulin M class antibodies. J. Clin. Microbiol. 1986, 23, 667–671. [Google Scholar] [CrossRef]
- Tounzen, M.R.; Epperson, D.; Taulman, J.F. Home range and habitat selection of Eastern gray squirrels (Sciurus carolinensis) in a small urban hardwood forest. Trans. Kans. Acad. Sci. 2012, 115, 89–101. [Google Scholar] [CrossRef]
- Tent, T.T.; Rongstad, O.J. Home Range and Survival of Cottontail Rabbits in Southwestern Wisconsin. J. Wildl. Manag. 1974, 38, 459–472. [Google Scholar]
- Kwiecinski, G.C. Marmota monax. Mamm. Species 1998, 591, 1–8. [Google Scholar] [CrossRef]
- Virginia Department of Conservation and Recreation, Division of Natural Heritage. Overview of the Physiography and Vegetation of Virginia; Virginia Department of Conservation and Recreation: Richmond, VA, USA, 2021. [Google Scholar]
- Cavallini, P. Variation in the social system of the red fox. Ethol. Ecol. Evol. 1996, 8, 323–342. [Google Scholar] [CrossRef]
- Virginia Department of Health, Office of Epidemiology. Virginia HIV Epidemiology Profile 2016; Virginia Department of Health: Richmond, VA, USA, 2016. [Google Scholar]
- Kosoy, O.; Rabe, I.; Geissler, A.; Adjemian, J.; Panella, A.; Laven, J.; Basile, A.J.; Velez, J.; Griffith, K.; Wong, D.; et al. Serological Survey for Antibodies to Mosquito-Borne Bunyaviruses Among US National Park Service and US Forest Service Employees. Vector Borne Zoonotic Dis. 2016, 16, 191–198. [Google Scholar] [CrossRef]
- Adjemian, J.; Weber, I.B.; McQuiston, J.; Griffith, K.S.; Mead, P.S.; Nicholson, W.; Roche, A.; Schriefer, M.; Fischer, M.; Kosoy, O.; et al. Zoonotic Infections Among Employees from Great Smoky Mountains and Rocky Mountain National Parks, 2008–2009. Vector Borne Zoonotic Dis. 2012, 12, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.B.; Peterson, K.E. Cross reactivity of neutralizing antibodies to the encephalitic California Serogroup orthobunyaviruses varies by virus and genetic relatedness. Sci. Rep. 2021, 11, 16424. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Number of Samples | LACV-Seroprevalence (when N > 15) |
|---|---|---|
| Eastern Gray Squirrel | 264 | 1.89% |
| Eastern Cottontail | 131 | 2.29% |
| Groundhog | 56 | 1.79% |
| Red Fox | 39 | 2.56% |
| Eastern Chipmunk | 23 | 0% |
| Eastern Fox Squirrel | 5 | - |
| Flying Squirrel | 5 | - |
| American Beaver | 3 | - |
| Gray Fox | 1 | - |
| Total | 527 | 1.90% |
| Species | Date Sampled (MM/DD/YYYY) | Age | Sex | VA County of Origin | Sero End-Point Titer |
|---|---|---|---|---|---|
| Eastern Gray Squirrel-1 | 06/06/2020 | Juvenile | M | Frederick | 1:160 |
| Eastern Gray Squirrel-2 | 05/21/2021 | Juvenile | U | Prince William | 1:40 |
| Groundhog-1 | 06/02/2021 | Sub-Adult | U | Loudoun | 1:320 |
| Eastern Gray Squirrel-3 | 09/09/2021 | Juvenile | F | Loudoun | 1:320 |
| Eastern Cottontail-1 | 03/31/2022 | Infant | U | Warren | 1:40 |
| Red Fox-1 | 04/03/2022 | Adult | U | Loudoun | 1:40 |
| Eastern Cottontail-2 | 04/29/2022 | Adult | F | Loudoun | 1:40 |
| Eastern Gray Squirrel-4 | 07/17/2022 | Adult | M | Fairfax | 1:40 |
| Eastern Cottontail-3 | 09/20/2022 | Juvenile | U | Loudoun | 1:40 |
| Eastern Gray Squirrel-5 | 10/03/2022 | Juvenile | F | Culpeper | 1:80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faw, L.R.; Riley, J.; Eastwood, G. La Crosse Virus Circulation in Virginia, Assessed via Serosurveillance in Wildlife Species. Infect. Dis. Rep. 2023, 15, 360-369. https://doi.org/10.3390/idr15040036
Faw LR, Riley J, Eastwood G. La Crosse Virus Circulation in Virginia, Assessed via Serosurveillance in Wildlife Species. Infectious Disease Reports. 2023; 15(4):360-369. https://doi.org/10.3390/idr15040036
Chicago/Turabian StyleFaw, Lindsey R., Jennifer Riley, and Gillian Eastwood. 2023. "La Crosse Virus Circulation in Virginia, Assessed via Serosurveillance in Wildlife Species" Infectious Disease Reports 15, no. 4: 360-369. https://doi.org/10.3390/idr15040036
APA StyleFaw, L. R., Riley, J., & Eastwood, G. (2023). La Crosse Virus Circulation in Virginia, Assessed via Serosurveillance in Wildlife Species. Infectious Disease Reports, 15(4), 360-369. https://doi.org/10.3390/idr15040036