
System Complexity in Influenza Infection and Vaccination: Effects upon Excess Winter Mortality



Abstract
1. The Excess Winter Mortality (EWM) Calculation
2. Introduction
3. Materials and Methods
3.1. Sources of the Data
3.2. Adjusting EWM for Each Country to a US-Equivalent
3.3. Method for Excluding Outlying EWM Values
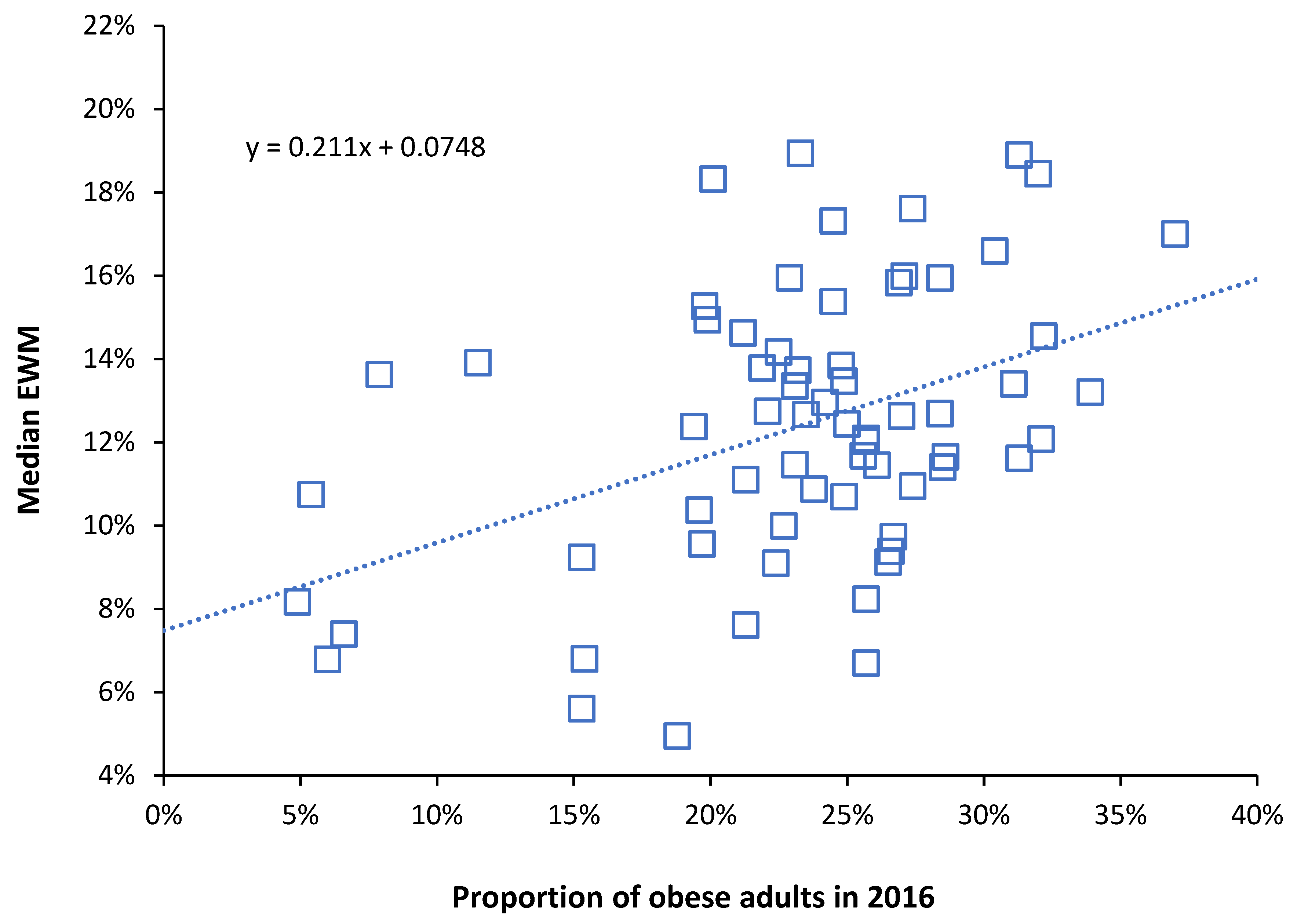
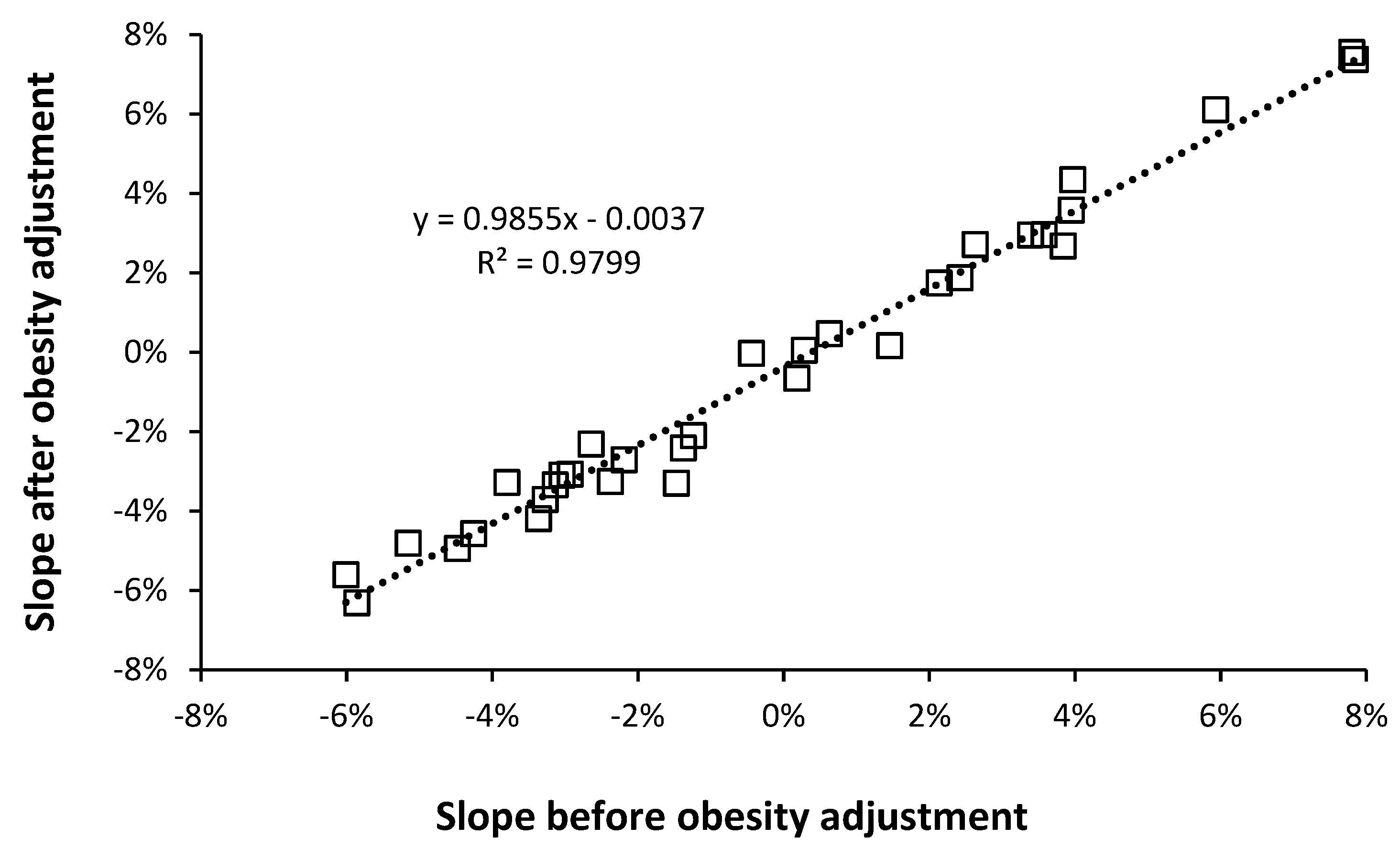
3.4. Adjustment of EWM for Obesity Relative to the USA
3.5. EWM in US States since 2008
3.6. Data Manipulation
4. Results
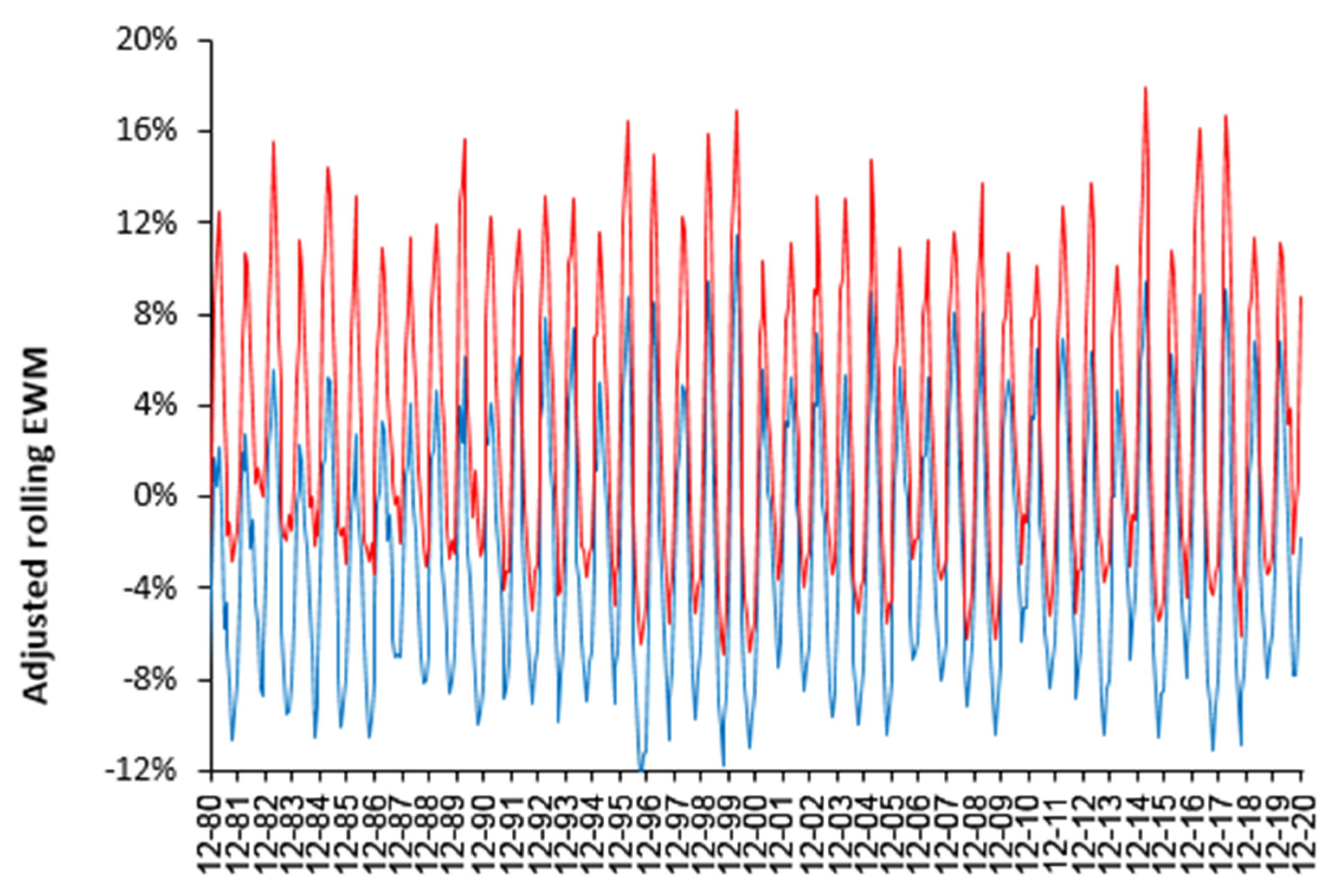
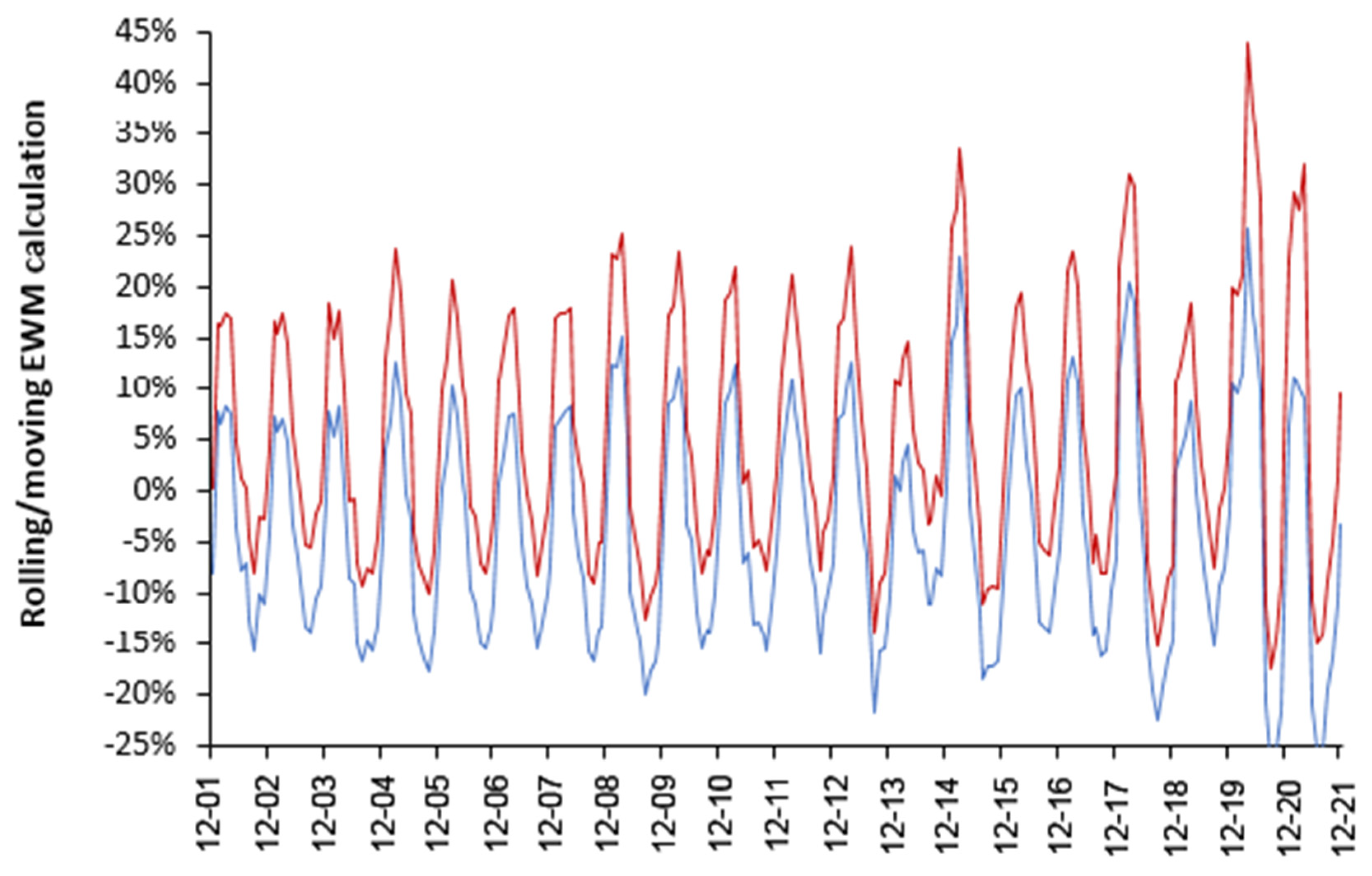
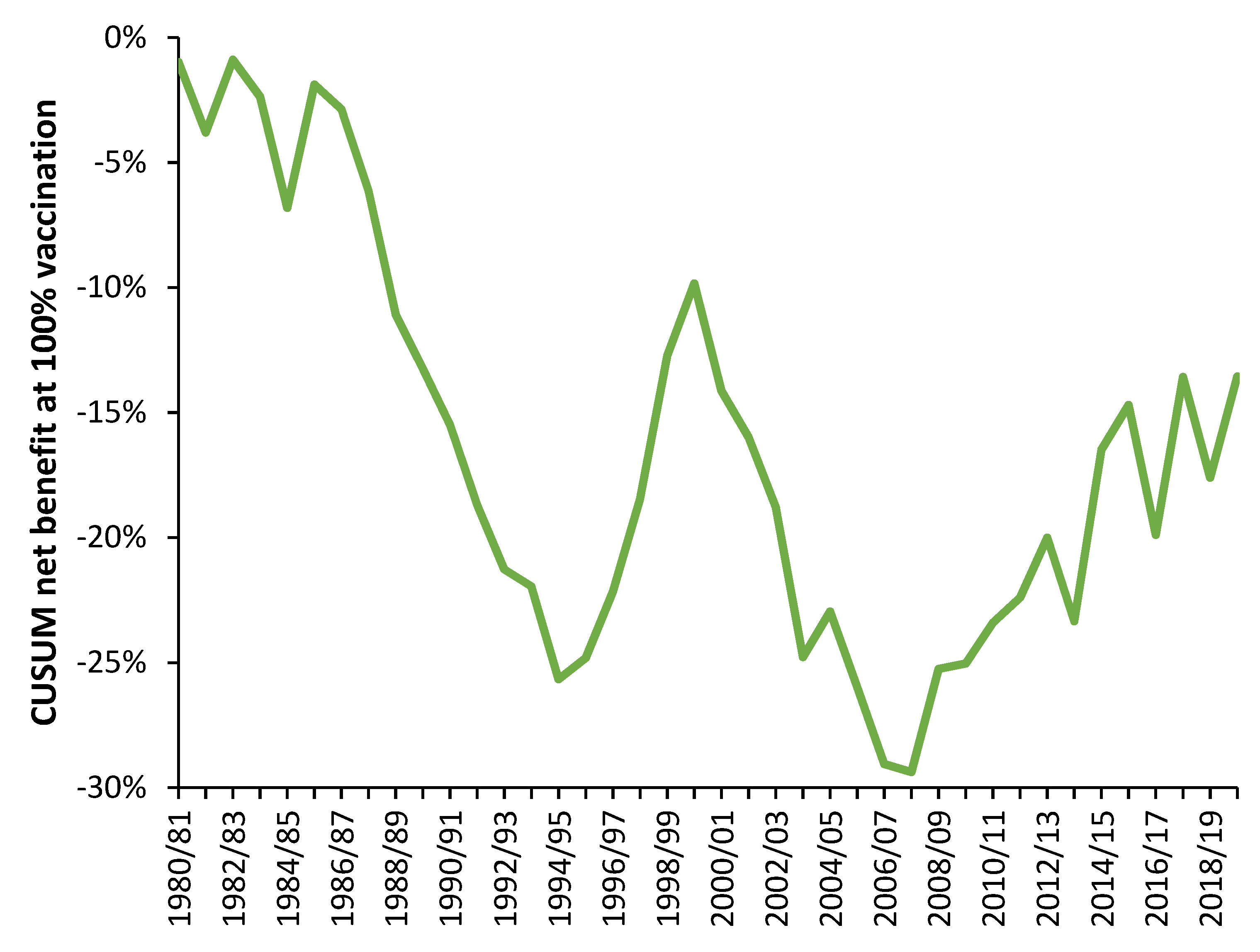
4.1. EWM Shows Extreme Spatiotemporal Volatility
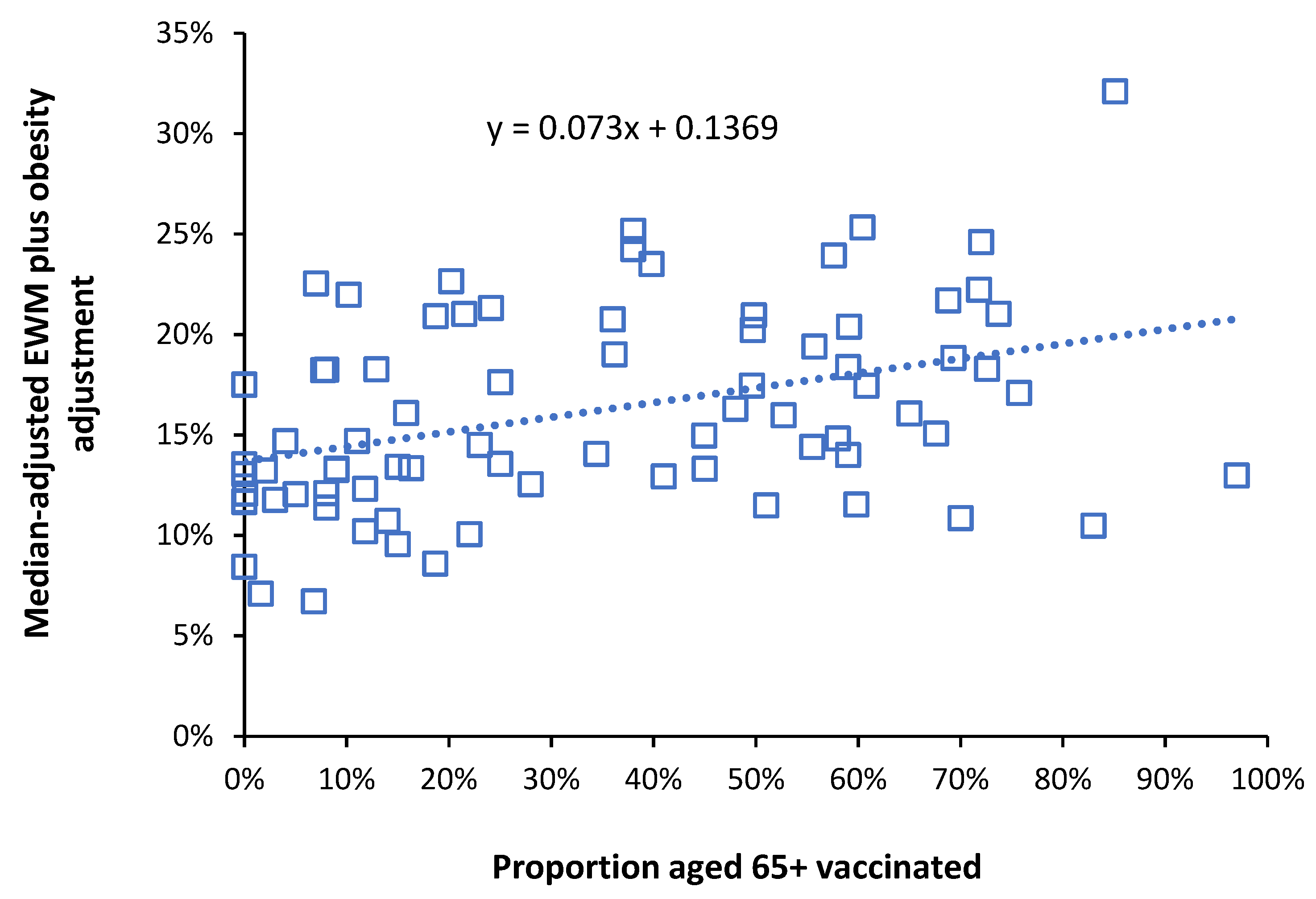
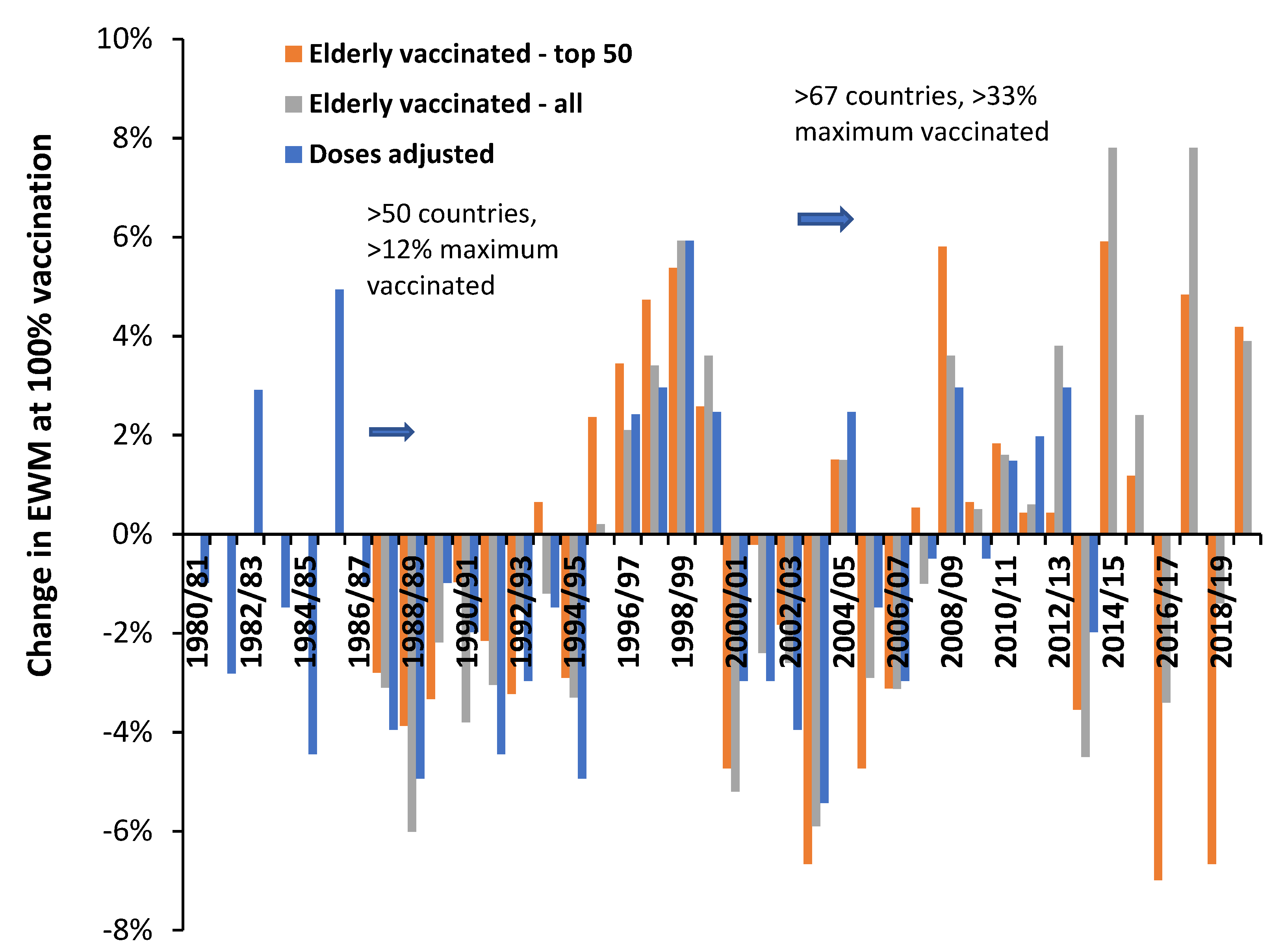
4.2. Influenza Vaccination in the Elderly
- Data from all available countries
- The 50 countries with the highest number of years of available data
- #1 plus data from US states (available for 2007/08 onward) [26]
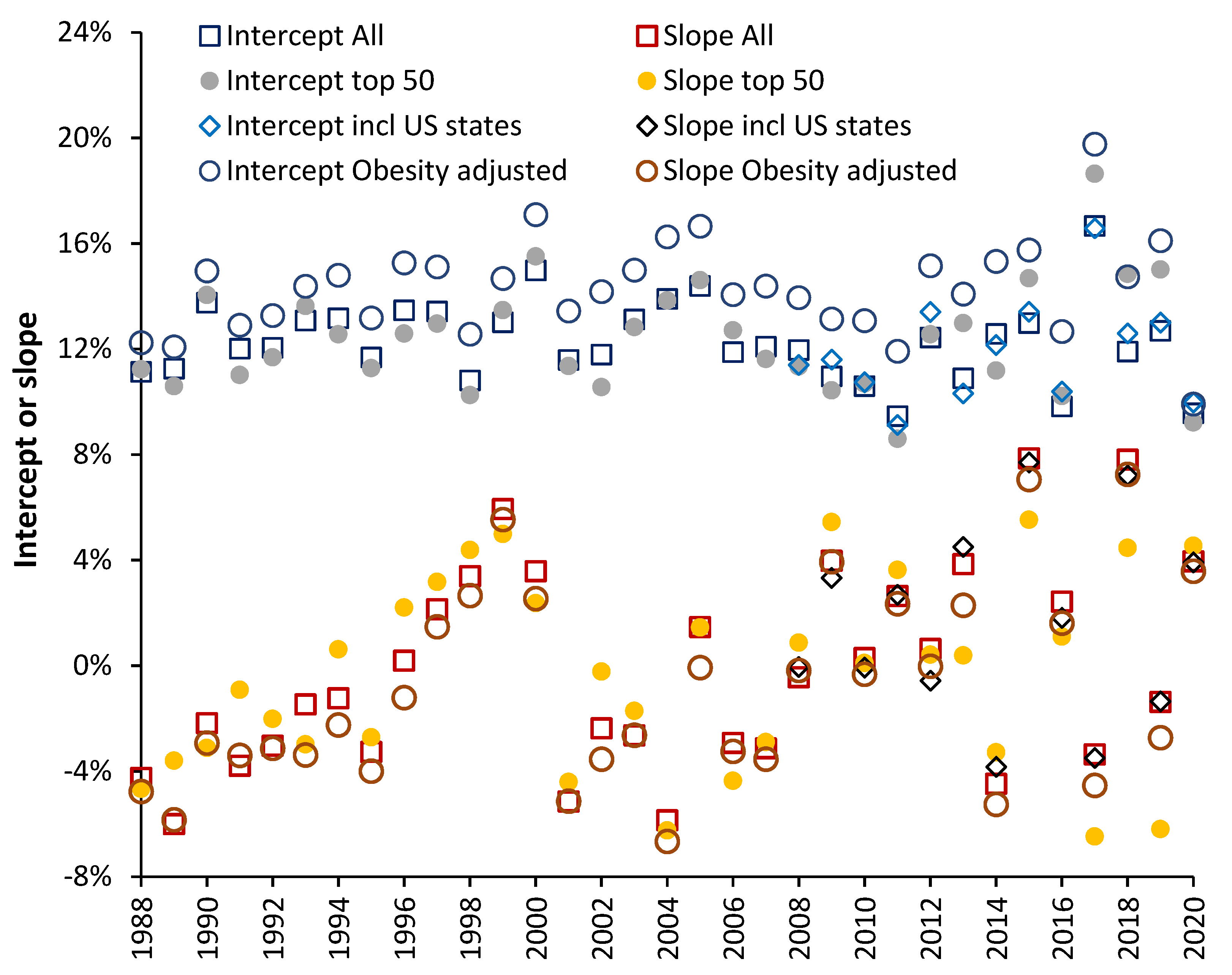
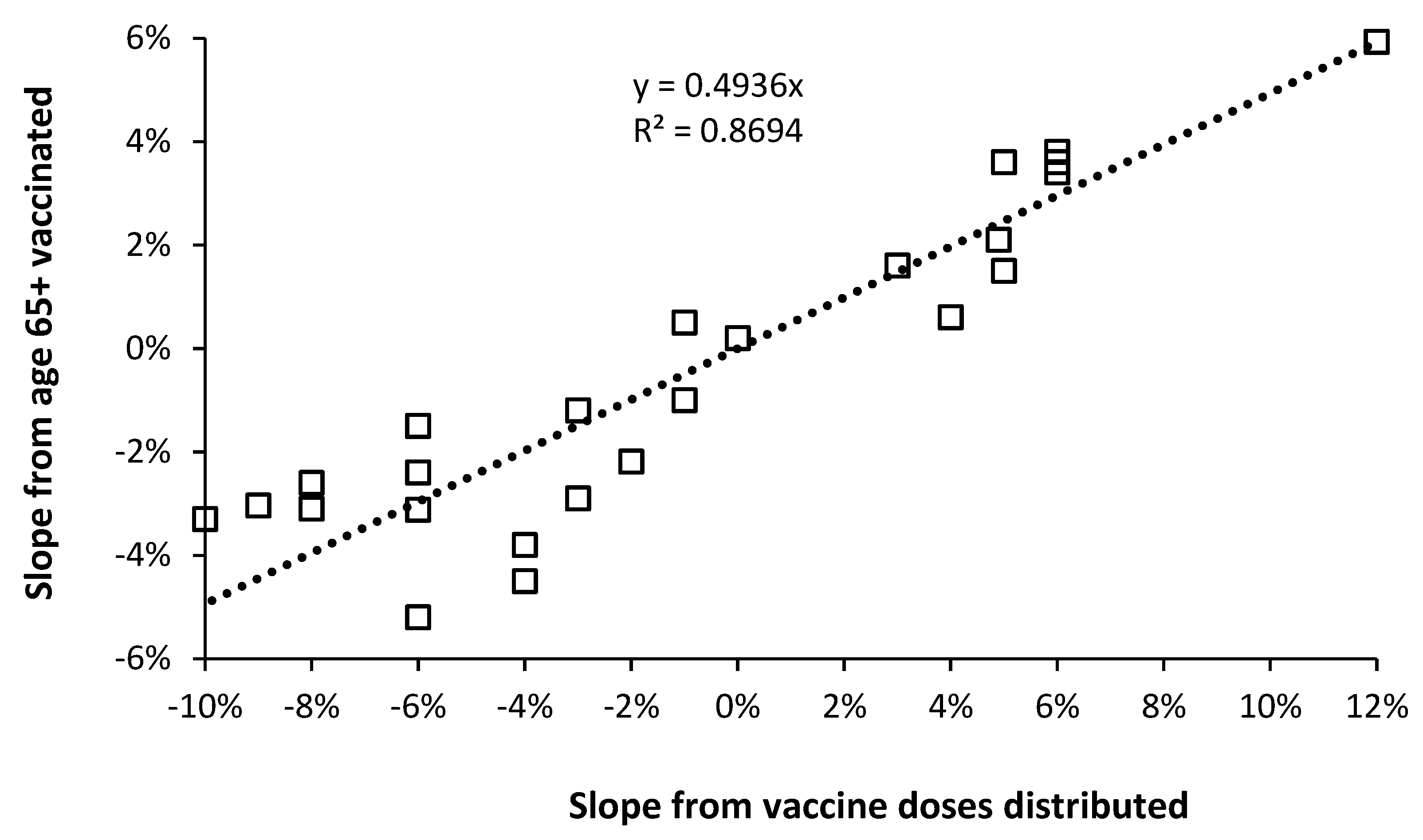
4.3. Comparison with a Previous Study
4.4. Further Validation of the Results
4.5. The Values for the Slope Follow an Extreme Value Distribution
5. Discussion
5.1. What Is the “Real” Long-Term Effect?
5.2. Limitations of Our Earlier Hypothesis
5.3. Adjustment for Obesity
5.4. Implications of High International Variation
5.5. 2014/15 as an Example of Poor Vaccine Matching
5.6. Roles for Pathogen Interference
5.7. Could Vaccine Effectiveness Be an Illusion Created by Pathogen Interference?
5.8. Heliobiology and Additional Hidden Complexity
5.9. Implications of an Extreme Value Distribution for the Slope
5.10. Minimum Value of the Slope
5.11. Biochemical and Immune Health
5.12. A Potential Basis for Extreme Variation in the Net Effects of Influenza Vaccination
6. Pragmatic Implications to Health Care Services
7. Implications to Influenza Policy
8. Limitations and Future Research
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A



References
- D’Angiolella, L.S.; Lafranconi, A.; Cortesi, P.A.; Rota, S.; Cesana, G.; Mantovani, L.G. Costs and effectiveness of influenza vaccination: A systematic review. Ann. Ist. Super. Sanita 2018, 54, 49–57. [Google Scholar]
- Boahen, C.; Joosten, L.; Netea, M.; Kumar, V. Conceptualization of population-specific human functional immune-genomics projects to identify factors that contribute to variability in immune and infectious diseases. Heliyon 2021, 7, e06755. [Google Scholar] [CrossRef] [PubMed]
- Burel, J.G.; Qian, Y.; Arlehamn, C.L.; Weiskopf, D.; Zapardiel-Gonzalo, J.; Taplitz, R.; Gilman, R.H.; Saito, M.; De Silva, A.D.; Vijayanand, P.; et al. An integrated workflow to assess technical and biological variability of cell population frequencies in human peripheral blood by flow cytometry. J. Immunol. 2017, 198, 1748–1758. [Google Scholar] [CrossRef] [PubMed]
- Brodin, P.; Davis, M. Human immune system variation. Nat. Rev. Immunol. 2017, 17, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Lakshmikanth, T.; Muhammad, S.A.; Olin, A.; Chen, Y.; Mikes, J.; Fagerberg, L.; Gummesson, A.; Bergström, G.; Uhlen, M.; Brodin, P. Human immune system variation during 1 year. Cell Rep. 2020, 32, 107923. [Google Scholar] [CrossRef]
- Alpert, A.; Pickman, Y.; Leipold, M.; Rosenberg-Hasson, Y.; Ji, X.; Gaujoux, R.; Rabani, H.; Starosvetsky, E.; Kveler, K.; Schaffert, S.; et al. A clinically meaningful metric of immune age derived from high-dimensional longitudinal monitoring. Nat. Med. 2019, 25, 487–495. [Google Scholar] [CrossRef]
- Jones, E.; Sheng, J.; Carlson, J.; Wang, S. Aging-induced fragility of the immune system. J. Theor. Biol. 2021, 510, 110473. [Google Scholar] [CrossRef]
- Glaser, R.; Kiecolt-Glaser, J. Stress-induced immune dysfunction, implications for health. Nat. Rev. Immunol. 2005, 5, 243–251. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, X.; Jiang, D.; Yan, D.; Guan, Z.; Zhou, Y.; Liu, X.; Huang, C.; Ding, C.; Lan, L.; et al. Associations between temperature and influenza activity, A national time series study in China. Int. J. Environ. Res. Public Health 2021, 18, 10846. [Google Scholar] [CrossRef]
- Lowen, A.C.; Mubareka, S.; Steel, J.; Palese, P. Influenza virus transmission is dependent on relative humidity and temperature. PLoS Pathog. 2007, 3, e151. [Google Scholar] [CrossRef]
- Roussel, M.; Pontier, D.; Cohen, J.-M.; Lina, B.; Fouchet, D. Quantifying the role of weather on seasonal influenza. BMC Public Health 2016, 16, 441. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Liu, T.; Geng, X.; Yang, G. Seasonal pattern of influenza and the association with meteorological factors based on wavelet analysis in Jinan City, Eastern China, 2013–2016. Peerj 2020, 8, e8626. [Google Scholar] [CrossRef] [PubMed]
- Peci, A.; Winter, A.-L.; Li, Y.; Gnaneshan, S.; Liu, J.; Mubareka, S.; Gubbay, J.B. Effects of absolute humidity, relative humidity, temperature, and wind speed on influenza activity in Toronto, Ontario, Canada. Appl. Environ. Microbiol. 2019, 85, e02426-18. [Google Scholar] [CrossRef] [PubMed]
- Kendon, M.; McCarthy, M. The UK’s wet and stormy winter of 2013/2014. Weather 2015, 70, 40–47. [Google Scholar] [CrossRef]
- Public Health England. Surveillance of Influenza and Other Respiratory Viruses in the United Kingdom, Winter 2013/14. June 2014. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/325203/Flu_annual_report_June_2014.pdf (accessed on 6 December 2021).
- Jiang, T. Modeling influenza sequence evolution for vaccination. BMC Proc. 2012, 6, P44. [Google Scholar] [CrossRef]
- Lam, E.K.S.; Morris, D.H.; Hurt, A.C.; Barr, I.G.; Russell, C.A. The impact of climate and antigenic evolution on seasonal influenza virus epidemics in Australia. Nat. Commun. 2020, 11, 2741. [Google Scholar] [CrossRef]
- Zhuang, Q.; Wang, S.; Liu, S.; Hou, G.; Li, J.; Jiang, W.; Wang, K.; Peng, C.; Liu, D.; Guo, A.; et al. Diversity and distribution of type A influenza viruses, an updated panorama analysis based on protein sequences. Virol. J. 2019, 16, 85. [Google Scholar] [CrossRef]
- Xue, K.S.; Stevens-Ayers, T.; Campbell, A.P.; Englund, J.A.; Pergam, S.A.; Boeckh, M.; Bloom, J.D. Parallel evolution of influenza across multiple spatiotemporal scales. eLife 2017, 6, e26875. [Google Scholar] [CrossRef]
- Jansen, A.J.G.; Spaan, T.; Low, H.Z.; Di Iorio, D.; Brand, J.V.D.; Tieke, M.; Barendrecht, A.; Rohn, K.; Van Amerongen, G.; Stittelaar, K.; et al. Influenza-induced thrombocytopenia is dependent on the subtype and sialoglycan receptor and increases with virus pathogenicity. Blood Adv. 2020, 4, 2967–2978. [Google Scholar] [CrossRef]
- Hu, J.; Ma, C.; Liu, X. PA-X, a key regulator of influenza A virus pathogenicity and host immune responses. Med. Microbiol. Immunol. 2018, 207, 255–269. [Google Scholar] [CrossRef] [PubMed]
- van Asten, L.; van den Wijngaard, C.; van Pelt, W.; van De Kassteele, J.; Meijer, A.; van Der Hoek, W.; Kretzschmar, M.; Koopmans, M. Mortality attributable to 9 common infections, significant effect of influenza A, respiratory syncytial virus, influenza B, norovirus, and parainfluenza in elderly persons. J. Infect. Dis. 2012, 206, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Seo, E.; Yoo, R.N.; Sung, H.; Lee, J. Clinical significance of viral-bacterial codetection among young children with respiratory tract infections, Findings of RSV, influenza, adenoviral infections. Medicine 2020, 99, e18504. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Blanchard, N. Microbial (co)infections, Powerful immune influencers. PLoS Pathog. 2022, 18, e1010212. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, S.; Ishida, T.; Tachibana, H.; Ito, Y.; Ito, A.; Hashimoto, T. Impact of bacterial coinfection on clinical outcomes in pneumococcal pneumonia. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1839–1847. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.P.; Ponomarenko, A. Trends in excess winter mortality (EWM) from 1900/01 to 2019/20—Evidence for a complex system of multiple long-term trends. Int. J. Environ. Res. Public Health 2022, 19, 3407. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. CDC Seasonal Flu Vaccine Effectiveness Studies. Available online: https://www.cdc.gov/flu/vaccines-work/effectiveness-studies.htm (accessed on 6 February 2021).
- Simonsen, L.; Reichert, T.A.; Viboud, C.; Blackwelder, W.C.; Taylor, R.J.; Miller, M.A. Impact of influenza vaccination on seasonal mortality in the US elderly population. Arch. Intern. Med. 2005, 165, 265–272. [Google Scholar]
- Rizzo, C.; Viboud, C.; Montomoli, E.; Simonsen, L.; Miller, M.A. Influenza–related mortality in the Italian elderly, no decline associated with increasing vaccination coverage. Vaccine 2006, 24, 6468–6475. [Google Scholar] [CrossRef]
- Siegenfeld, A.; Bar-Yam, Y. An introduction to complex systems science and its applications. Complexity 2020, 2020, 6105872. [Google Scholar] [CrossRef]
- Rutter, H.; Savona, N.; Glonti, K.; Bibby, J.; Cummins, S.; Finegood, D.T.; Greaves, F.; Harper, L.; Hawe, P.; Moore, L.; et al. The need for a complex systems model of evidence for public health. Lancet 2017, 390, 2602–2604. [Google Scholar] [CrossRef]
- Serpa, C.; Forouharfar, A. Fractalization of chaos and complexity, Proposition of a new method in the study of complex systems. In Chaos, Complexity and Leadership 2020; Erçetin, Ş.Ş., Açıkalın, Ş.N., Vajzović, E., Eds.; Springer Proceedings in Complexity; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Adak, D.; Bairagi, N. Bifurcation analysis of a multidelayed HIV model in presence of immune response and understanding of in-host viral dynamics. Math. Methods Appl. Sci. 2019, 42, 4256–4272. [Google Scholar] [CrossRef]
- McKenzie, D.R.; Hart, R.; Bah, N.; Ushakov, D.S.; Muñoz-Ruiz, M.; Feederle, R.; Hayday, A.C. Normality sensing licenses local T cells for innate-like tissue surveillance. Nat. Immunol. 2022, 23, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Rickenbach, C.; Gericke, C. Specificity of Adaptive Immune Responses in Central Nervous System Health, Aging and Diseases. Front. Neurosci. 2022, 15, 806260. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.A.; Sahu, A.R.; Saxena, S.; Hussain, S.; Pandey, A.; Kanchan, S.; Sahoo, A.P.; Mishra, B.; Tiwari, A.K.; Mishra, B.P.; et al. Systems biology approach, Panacea for unravelling host-virus interactions and dynamics of vaccine induced immune response. Gene Rep. 2016, 5, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Banks, H.T.; Davidian, M.; Hu, S.; Kepler, G.M.; Rosenberg, E.S. Modelling HIV immune response and validation with clinical data. J. Biol. Dyn. 2008, 2, 357–385. [Google Scholar] [CrossRef]
- Craddock, T.J.; Fritsch, P.; Rice, M.A., Jr.; del Rosario, R.M.; Miller, D.B.; Fletcher, M.A.; Klimas, N.G.; Broderick, G. A Role for Homeostatic Drive in the Perpetuation of Complex Chronic Illness, Gulf War Illness and Chronic Fatigue Syndrome. PLoS ONE 2014, 9, e84839. [Google Scholar] [CrossRef]
- Waters, R.S.; Perry, J.S.A.; Han, S.; Bielekova, B.; Gedeon, T. The effects of interleukin-2 on immune response regulation. Math. Med. Biol. 2018, 35, 79–119. [Google Scholar] [CrossRef]
- El Karkri, J.; Boudchich, F.; Volpert, V.; Aboulaich, R. Stability Analysis of a Delayed Immune Response Model to Viral Infection. Differ. Equ. Dyn. Syst. 2022. [Google Scholar] [CrossRef]
- Elaiw, A.; AlShamrani, N. Stability of an adaptive immunity pathogen dynamics model with latency and multiple delays. Math. Methods Appl. Sci. 2018, 41, 6645–6672. [Google Scholar] [CrossRef]
- Bocharov, G.; Meyerhans, A.; Bessonov, N.; Trofimchuk, S.; Volpert, V. Modelling the dynamics of virus infection and immune response in space and time. Int. J. Parallel Emergent Distrib. Syst. 2017, 34, 341–355. [Google Scholar] [CrossRef]
- Wang, A.; Xiao, Y.; Smith, R. Multiple Equilibria in a Non-smooth Epidemic Model with Medical-Resource Constraints. Bull. Math. Biol. 2019, 81, 963–994. [Google Scholar] [CrossRef]
- Reluga, T.; Medlock, J.; Perelson, A. Backward bifurcations and multiple equilibria in epidemic models with structured immunity. J. Theor. Biol. 2008, 252, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Kumar, S. Dynamic analysis of the role of innate immunity in SEIS epidemic model. Eur. Phys. J. Plus 2021, 136, 439. [Google Scholar] [CrossRef] [PubMed]
- Majnarić-Trtica, L.; Vitale, B. Systems biology as a conceptual framework for research in family medicine, use in predicting response to influenza vaccination. Prim. Health Care Res. Dev. 2011, 12, 310–321. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jones, R. Excess winter mortality (EWM) as a dynamic forensic tool, Where, when, which conditions, gender, ethnicity, and age. Int. J. Environ. Res. Public Health 2021, 18, 2161. [Google Scholar] [CrossRef]
- WHO. Prevalence of Obesity among Adults. Available online: https://www.who.int/data/gho/indicator-metadata-registry/imr-details/2389 (accessed on 3 March 2022).
- Global Obesity Observatory. World Obesity Federation Global Obesity Observatory. Available online: https://data.worldobesity.org/ (accessed on 3 March 2022).
- CDC. Vaccine Coverage among Nursing Home Residents. Available online: https://www.cdc.gov/flu/fluvaxview/interactive-nursing-home.htm (accessed on 3 March 2022).
- OECD Data. Influenza Vaccination Rates. Available online: https://data.oecd.org/healthcare/influenza-vaccination-rates.htm (accessed on 12 March 2022).
- Wachs, S. What Is a CUSUM Chart and When Should I Use One? Available online: https://accendoreliability.com/cusum-chart-use-one/ (accessed on 12 March 2022).
- Public Health England. Surveillance of Influenza and Other Respiratory Viruses in the United Kingdom, Winter 2014 to 2015. May 2015. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/429617/Annualreport_March2015_ver4.pdf (accessed on 25 February 2022).
- Jones, R. Year-to-year variation in deaths in English Output Areas (OA), and the interaction between a presumed infectious agent and influenza in 2015. SMU Med. J. 2017, 4, 37–69. Available online: http//smu.edu.in/content/dam/manipal/smu/smims/Volume4No2July2017/SMU%20Med%20J%20(July%202017)%20-%204.pdf (accessed on 25 March 2022).
- Gonçalves, A.R.; Iten, A.; Suter-Boquete, P.; Schibler, M.; Kaiser, L.; Cordey, S. Hospital surveillance of influenza strains: A concordant image of viruses identified by the Swiss Sentinel system? Influenza Other Resp Viruses 2017, 11, 41–47. [Google Scholar] [CrossRef]
- Australian Government, Department of Health. Australian Influenza Surveillance Report. Available online: https://www1.health.gov.au/internet/main/publishing.nsf/Content/cda-surveil-ozflu-flucurr.htm (accessed on 12 March 2022).
- Simonsen, L.; Viboud, C. The Art of Modeling the Mortality Impact of Winter-Seasonal Pathogens. J. Infect. Dis. 2012, 206, 625–627. [Google Scholar] [CrossRef]
- National Foundation for Infectious Diseases. Respiratory Syncytial Virus in Older Adults, A Hidden Annual Epidemic. Available online: https://www.nfid.org/wp-content/uploads/2019/08/rsv-report.pdf (accessed on 12 March 2022).
- Taubenberger, J.K.; Morens, D.M. The pathology of influenza virus infections. Annu. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef]
- Opatowski, L.; Baguelin, M.; Eggo, R.M. Influenza interaction with cocirculating pathogens and its impact on surveillance, pathogenesis, and epidemic profile, A key role for mathematical modelling. PLoS Pathog. 2018, 14, e1006770. [Google Scholar] [CrossRef]
- Zheng, X.; Song, Z.; Li, Y.; Zhang, J.; Wang, X.-L. Possible interference between seasonal epidemics of influenza and other respiratory viruses in Hong Kong, 2014–2017. BMC Infect. Dis. 2017, 17, 772. [Google Scholar] [CrossRef]
- Hedberg, P.; Johansson, N.; Ternhag, A.; Abdel-Halim, L.; Hedlund, J.; Nauclér, P. Bacterial co-infections in community-acquired pneumonia caused by SARS-CoV-2, influenza virus and respiratory syncytial virus. BMC Infect. Dis. 2022, 22, 108. [Google Scholar] [CrossRef] [PubMed]
- Novella, S. COVID-19 Lockdown and the Flu. Neurologica Blog, 15 June 2020. Available online: https://theness.com/neurologicablog/index.php/covid-19-lockdown-and-the-flu/ (accessed on 11 March 2022).
- Doroshenko, A.; Lee, N.; MacDonald, C.; Zelyas, N.; Asadi, L.; Kanji, J.N. Decline of Influenza and Respiratory Viruses with COVID-19 Public Health Measures, Alberta, Canada. Mayo Clin. Proc. 2021, 96, 3042–3052. [Google Scholar] [CrossRef] [PubMed]
- PHE. National Influenza Report. 1 October 2020—Week 40 Report. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/923246/National_Influenza_report_1_October_2020_week_40.pdf (accessed on 12 March 2022).
- Institute for Government. Timeline for UK Coronavirus Lockdowns. Available online: https://www.instituteforgovernment.org.uk/charts/uk-government-coronavirus-lockdowns (accessed on 1 January 2022).
- Koutsakos, M.; Wheatley, A.K.; Laurie, K.; Kent, S.J.; Rockman, S. Influenza lineage extinction during the COVID-19 pandemic? Nat. Rev. Microbiol. 2021, 19, 741–742. [Google Scholar] [CrossRef] [PubMed]
- Danino, D.; Ben-Shimol, S.; van der Beek, B.A.; Givon-Lavi, N.; Avni, Y.S.; Greenberg, D.; Weinberger, D.M.; Dagan, R. Decline in Pneumococcal Disease in Young Children during the COVID-19 Pandemic in Israel Associated with Suppression of seasonal Respiratory Viruses, despite Persistent Pneumococcal Carriage, A Prospective Cohort Study. Clin. Infect. Dis. 2021, ciab1014. [Google Scholar]
- Mondal, P.; Sinharoy, A.; Gope, S. The Influence of COVID-19 on Influenza and Respiratory Syncytial Virus Activities. Infect. Dis. Rep. 2022, 14, 17. [Google Scholar] [CrossRef]
- Dadashi, M.; Khaleghnejad, S.; Elkhichi, P.A.; Goudarzi, M.; Goudarzi, H.; Taghavi, A.; Vaezjalali, M.; Hajikhani, B. COVID-19 and Influenza Co-infection, A Systematic Review and Meta-Analysis. Front. Med. 2021, 8, 681469. [Google Scholar] [CrossRef]
- Jackson, M.L.; Nelson, J.C. The test-negative design for estimating influenza vaccine effectiveness. Vaccine 2013, 31, 2165–2168. [Google Scholar] [CrossRef]
- Fireman, B.; Lee, J.; Lewis, N.; Bembom, O.; van der Laan, M.; Baxter, R. Influenza vaccination and mortality, differentiating vaccine effects from bias. Am. J. Epidemiol. 2009, 170, 650–656. [Google Scholar] [CrossRef]
- Cowling, B.J.; Fang, V.J.; Nishiura, H.; Chan, K.-H.; Ng, S.; Ip, D.K.M.; Chiu, S.S.; Leung, G.M.; Peiris, J.S.M. Increased risk of noninfluenza respiratory virus infections associated with receipt of inactivated influenza vaccine. Clin. Infect. Dis. 2012, 54, 1778–1783. [Google Scholar] [CrossRef]
- Rikin, S.; Jia, H.; Vargas, C.Y.; de Belliard, Y.C.; Reed, C.; LaRussa, P.; Larson, E.L.; Saiman, L.; Stockwell, M.S. Assessment of temporally–related acute respiratory illness following influenza vaccination. Vaccine 2018, 36, 1958–1964. [Google Scholar] [CrossRef]
- Hansen, K.P.; Benn, C.S.; Aamand, T.; Buus, M.; da Silva, I.; Aaby, P.; Fisker, A.B.; Thysen, S.M. Does Influenza Vaccination during Pregnancy Have Effects on Non-Influenza Infectious Morbidity? A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Vaccines 2021, 9, 1452. [Google Scholar] [CrossRef] [PubMed]
- Van Beek, J.; Veenhoven, R.H.; Bruin, J.P.; Boxtel, R.A.J.V.; De Lange, M.M.A.; Meijer, A.; Sanders, E.A.M.; Rots, N.Y.; Luytjes, W. Influenza-like Illness Incidence Is Not Reduced by Influenza Vaccination in a Cohort of Older Adults, Despite Effectively Reducing Laboratory-Confirmed Influenza Virus Infections. J. Infect. Dis. 2017, 216, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Lauzon-Joset, J.; Scott, N.; Mincham, K.; Stumbles, P.; Holt, P.; Strickland, D. Pregnancy Induces a Steady-State Shift in Alveolar Macrophage M1/M2 Phenotype That Is Associated with a Heightened Severity of Influenza Virus Infection, Mechanistic Insight Using Mouse Models. J Infect. Dis. 2019, 219, 1823–1831. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, B.N.; Hunter, J.F.; Cross, M.P.; Acevedo, A.M.; Pressman, S.D. When is affect variability bad for health? The association between affect variability and immunee response to the influenza vaccination. J. Psychosom. Res. 2018, 104, 41–47. [Google Scholar] [CrossRef]
- Berencsi, G.; Kapusinszky, B.; Rigó, Z.; Szomor, K. Interference among viruses circulating and administered in Hungary from 1931 to 2008. Acta Microbiol. Immunol. Hung. 2010, 57, 73–86. [Google Scholar] [CrossRef]
- NOAA. Coronal Mass Ejections. Available online: https://www.swpc.noaa.gov/phenomena/coronal-mass-ejections (accessed on 7 March 2022).
- Madanchi, A.; Absalan, M.; Lohmann, G.M.; Anvari, M.; Tabar, M.R.R. Strong short-term non-linearity of solar irradiance fluctuations. Sol. Energy 2017, 144, 1–9. [Google Scholar] [CrossRef]
- Výbošt’oková, T.; Švanda, M. Statistical analysis of the correlation between anomalies in the Czech electric power grid and geomagnetic activity. Space Weather 2019, 17, 1208–1218. [Google Scholar] [CrossRef]
- Zenchenko, T.A.; Breus, T.K. The Possible Effect of Space Weather Factors on Various Physiological Systems of the Human Organism. Atmosphere 2021, 12, 346. [Google Scholar] [CrossRef]
- Palmer, S.J.; Rycroft, M.J.; Cermack, M. Solar and geomagnetic activity, extremely low frequency magnetic and electric fields and human health at the Earth’s surface. Surv. Geophys. 2006, 27, 557–595. [Google Scholar] [CrossRef]
- Papathanasopoulos, P.; Preka-Papadema, P.; Gkotsinas, A.; Dimisianos, N.; Hillaris, A.; Katsavrias, C.; Antonakopoulos, G.; Moussas, X.; Andreadou, E.; Georgiou, V.; et al. The possible effects of the solar and geomagnetic activity on multiple sclerosis. Clin. Neurol. Neurosurg. 2016, 146, 82–89. [Google Scholar] [CrossRef]
- Goldwater, P.N.; Oberg, E.O. Infection, Celestial Influences, and Sudden Infant Death Syndrome: A New Paradigm. Cureus 2021, 13, 17449. [Google Scholar] [CrossRef] [PubMed]
- Zaporozhan, V.; Ponomarenko, A. Mechanisms of geomagnetic field influence on gene expression using influenza as a model system, basics of physical epidemiology. Int. J. Environ. Res. Public Health 2010, 7, 938–965. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.W. A hypothesis: Sunspot cycles may detect pandemic influenza A in 1700-2000 A.D. Med. Hypotheses 2006, 67, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D.P. Influenza pandemics, solar activity cycles, and vitamin D. Med. Hypotheses 2010, 74, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Nasirpour, M.H.; Sharifi, A.; Ahmadi, M.; Jafarzadeh Ghoushchi, S. Revealing the relationship between solar activity and COVID-19 and forecasting of possible future viruses using multi-step autoregression (MSAR). Environ. Sci. Pollut. Res. Int. 2021, 28, 38074–38084. [Google Scholar] [CrossRef]
- Principia Scientific International. Weakening Trend in Solar Cycles Since SC21. Available online: https://nextgrandminimum.wordpress.com/2019/06/20/weakening-trend-in-solar-cycle-since-sc21/ (accessed on 11 April 2022).
- Ramos, P.; Louzada, F.; Ramos, E.; Dey, S. The Fréchet distribution: Estimation and application—An overview. J. Stat. Manag. Syst. 2020, 23, 549–578. [Google Scholar] [CrossRef]
- Jones, R.; Sleet, G.; Pearce, O.; Wetherill, M. Complex changes in blood biochemistry revealed by a composite score derived from Principal Component Analysis: Effects of age, patient acuity, end of life, day-of week, and potential insights into the issues surrounding the ‘Weekend’ effect in hospital mortality. J. Adv. Med. Med. Res. 2016, 18, 1–28. [Google Scholar] [CrossRef]
- von Wyl, V. Proximity to death and health care expenditure increase revisited: A 15-year panel analysis of elderly persons. Health Econ. Rev. 2019, 9, 9–16. [Google Scholar] [CrossRef]
- Vogel, N.; Schilling, O.K.; Wahl, H.-W.; Beekman, A.T.F.; Penninx, B.W.J.H. Time-to-death-related change in positive and negative affect among older adults approaching the end of life. Psychol. Aging 2013, 28, 128–141. [Google Scholar] [CrossRef]
- White, N.; Cunningham, W. Is Terminal Drop Pervasive or Specific? J. Gerontol. 1988, 43, P141–P144. [Google Scholar] [CrossRef]
- Bäckman, L.; Macdonald, S.W. Death and Cognition. Eur. Psychol. 2006, 11, 224–235. [Google Scholar] [CrossRef]
- Stanaway, F.F.; Gnjidic, D.; Blyth, F.M.; Le Couteur, D.; Naganathan, V.; Waite, L.; Seibel, M.; Handelsman, D.J.; Sambrook, P.N.; Cumming, R. How fast does the Grim Reaper walk? Receiver operating characteristics curve analysis in healthy men aged 70 and over. BMJ 2011, 343, d7679. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.; Smith, C.; Garten, R.J.; Levine, M.Z.; Chung, J.R.; Jackson, M.L.; Jackson, L.A.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; et al. Influence of Birth Cohort on Effectiveness of 2015–2016 Influenza Vaccine Against Medically Attended Illness Due to 2009 Pandemic Influenza A(H1N1) Virus in the United States. J. Infect. Dis. 2018, 218, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Kissling, E.; Pozo, F.; Buda, S.; Vilcu, A.M.; Gherasim, A.; Brytting, M.; Domegan, L.; Gómezet, V.; Meijer, A.; Lazar, M.; et al. Low 2018/19 vaccine effectiveness against influenza A(H3N2) among 15-64-year-olds in Europe: Exploration by birth cohort. Eurosurveillance 2019, 24, 1900604. [Google Scholar] [CrossRef]
- Sanniklova, T.; Romanyukha, A.; Barbi, E.; Caselli, G.; Franceschi, C.; Yashin, A. Modeling of Immunosenescence and Risk of Death from Respiratory Infections: Evaluation of the Role of Antigenic Load and Population Heterogeneity. Math. Model. Nat. Phenom. 2017, 12, 48–62. [Google Scholar] [CrossRef]
- Kaczorowski, K.J.; Shekhar, K.; Nkulikiyimfura, D.; Dekker, C.L.; Maecker, H.; Davis, M.M.; Chakraborty, A.K.; Brodin, P. Continuous immunotypes describe human immune variation and predict diverse responses. Proc. Natl. Acad. Sci. USA 2017, 114, E6097–E6106. [Google Scholar] [CrossRef]
- Jones, R. A Study of an unexplained and large increase in respiratory deaths in England and Wales: Is the pattern of diagnoses consistent with the potential involvement of Cytomegalovirus? J. Adv. Med. Med. Res. 2014, 4, 5179–5192. Available online: https://journaljammr.com/index.php/JAMMR/article/view/15226 (accessed on 1 January 2022). [CrossRef]
- Jones, R. Multidisciplinary insights into health care financial risk and hospital surge capacity, Part 3: Outbreaks of a new type or kind of disease create unique risk patterns and confounds traditional trend analysis. J. Health Care Financ. 2021, 47, 1–57. Available online: http://healthfinancejournal.com/index.php/johcf/article/view/242 (accessed on 1 January 2022).
- Franklin, B.; Hochlaf, D. An Economic Analysis of Flu Vaccination. The International Longevity Centre—UK. Available online: An-economic-analysis-of-flu-vaccination-18(1).pdf (accessed on 20 March 2022).
- Anderson, M.L.; Dobkin, C.; Gorry, D. The Effect of Influenza Vaccination for the Elderly on Hospitalization and Mortality: An Observational Study with a Regression Discontinuity Design. Ann. Intern. Med. 2020, 172, 445–452. [Google Scholar] [CrossRef]
- Van Ourti, T.; Bouckaert, N. The Dutch influenza vaccination policy and medication use, outpatient visits, hospitalization and mortality at age 65. Eur. J. Public Health 2020, 30, 275–280. [Google Scholar] [CrossRef]
- Sirois, C.; Boiteau, V.; Chiu, Y.; Gilca, R.; Simard, M. Exploring the associations between polypharmacy and COVID-19-related hospitalisations and deaths: A population-based cohort study among older adults in Quebec, Canada. BMJ Open 2022, 12, e060295. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
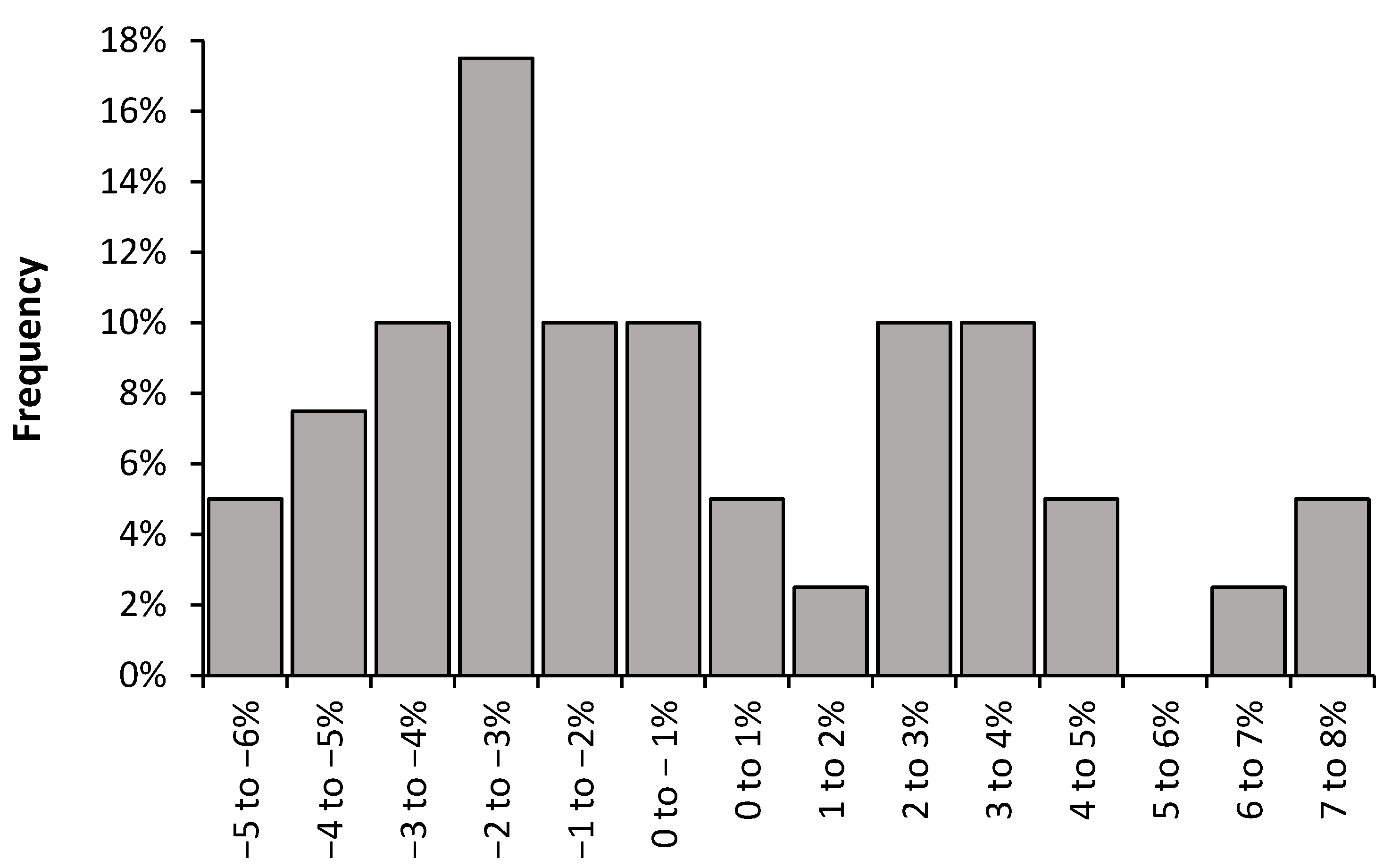{kind=link}
{kind=link}
{kind=link}
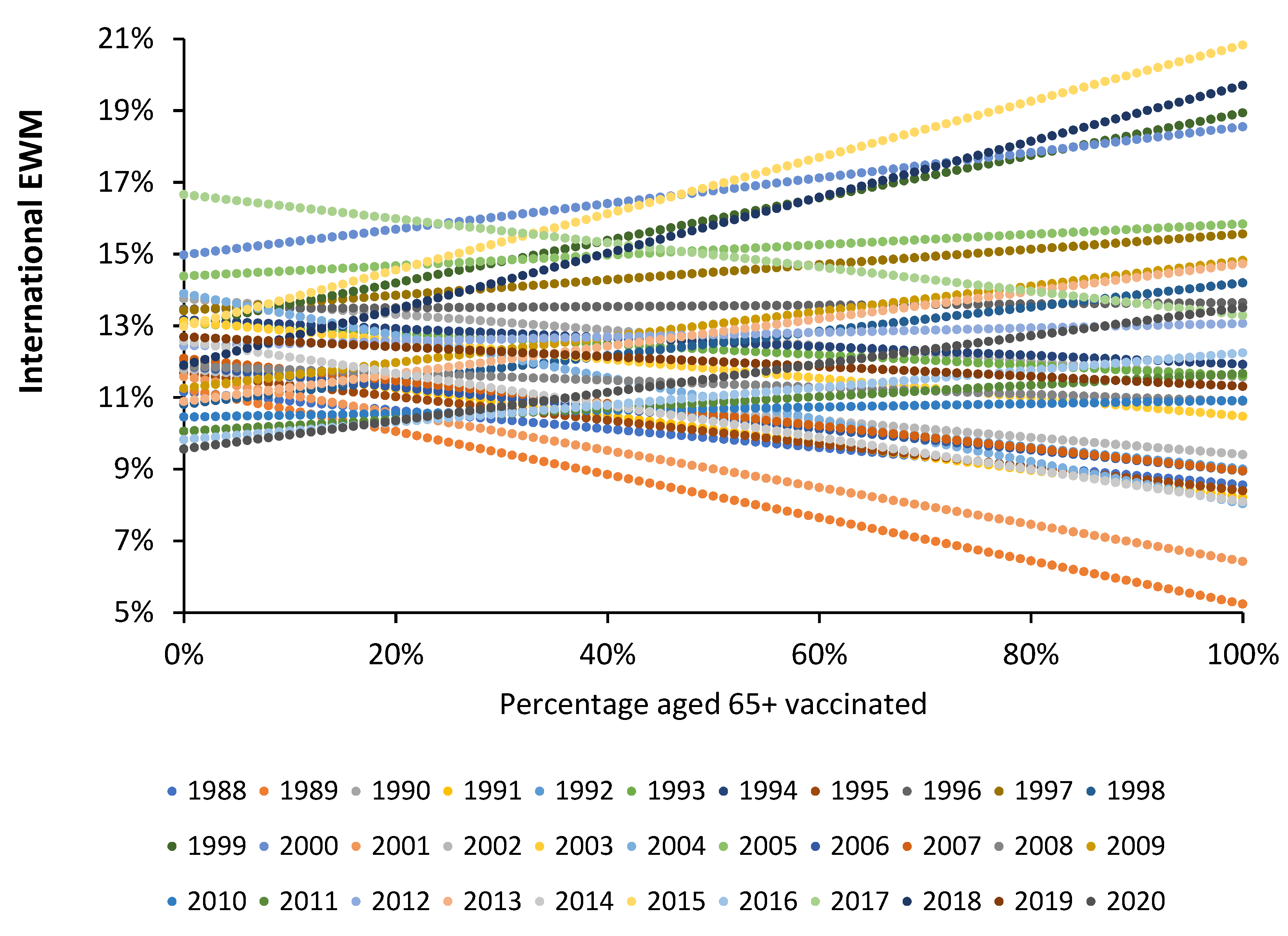{kind=link}
| Search String | Documents Identified |
|---|---|
| Influenza epidemics “complex systems” | 94,800 |
| Influenza and “systems biology” | 22,000 |
| Complex system dynamics pandemic influenza | 18,800 |
| Interactions influenza and “other pathogens” | 16,200 |
| Influenza and “pathogen interactions” | 14,600 |
| Influenza and “complex system” | 10,900 |
| Influenza vaccination and “complex system” | 4520 |
| Influenza and “pertussis complex relationship” | 570 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, R.P.; Ponomarenko, A. System Complexity in Influenza Infection and Vaccination: Effects upon Excess Winter Mortality. Infect. Dis. Rep. 2022, 14, 287-309. https://doi.org/10.3390/idr14030035
Jones RP, Ponomarenko A. System Complexity in Influenza Infection and Vaccination: Effects upon Excess Winter Mortality. Infectious Disease Reports. 2022; 14(3):287-309. https://doi.org/10.3390/idr14030035
Chicago/Turabian StyleJones, Rodney P., and Andriy Ponomarenko. 2022. "System Complexity in Influenza Infection and Vaccination: Effects upon Excess Winter Mortality" Infectious Disease Reports 14, no. 3: 287-309. https://doi.org/10.3390/idr14030035
APA StyleJones, R. P., & Ponomarenko, A. (2022). System Complexity in Influenza Infection and Vaccination: Effects upon Excess Winter Mortality. Infectious Disease Reports, 14(3), 287-309. https://doi.org/10.3390/idr14030035