
Cerebellar Contributions to Hypokinetic Symptoms in an Acute Lesion Parkinsonism Model

 ,
,  ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Rats and Experimental Groups
2.2. Electrodes
- Recording electrodes consisted of a stainless-steel monopolar bar (diameter: 250 μm, impedance: 3 MΩ) with a length adjusted to the target structure. The electrodes were soldered to male D-Sub crimp pins and insulated with resin, exposing 1 mm of the tip.
- Reference electrodes consisted of a No. 7 steel screw, 70%-covered with a copper wire (leaving the upper part of the screw exposed). The screw was connected to a male D-Sub crimp pin, with both joints soldered together. It was placed arbitrarily to avoid interference with the recording electrodes.
2.3. Surgical Procedures
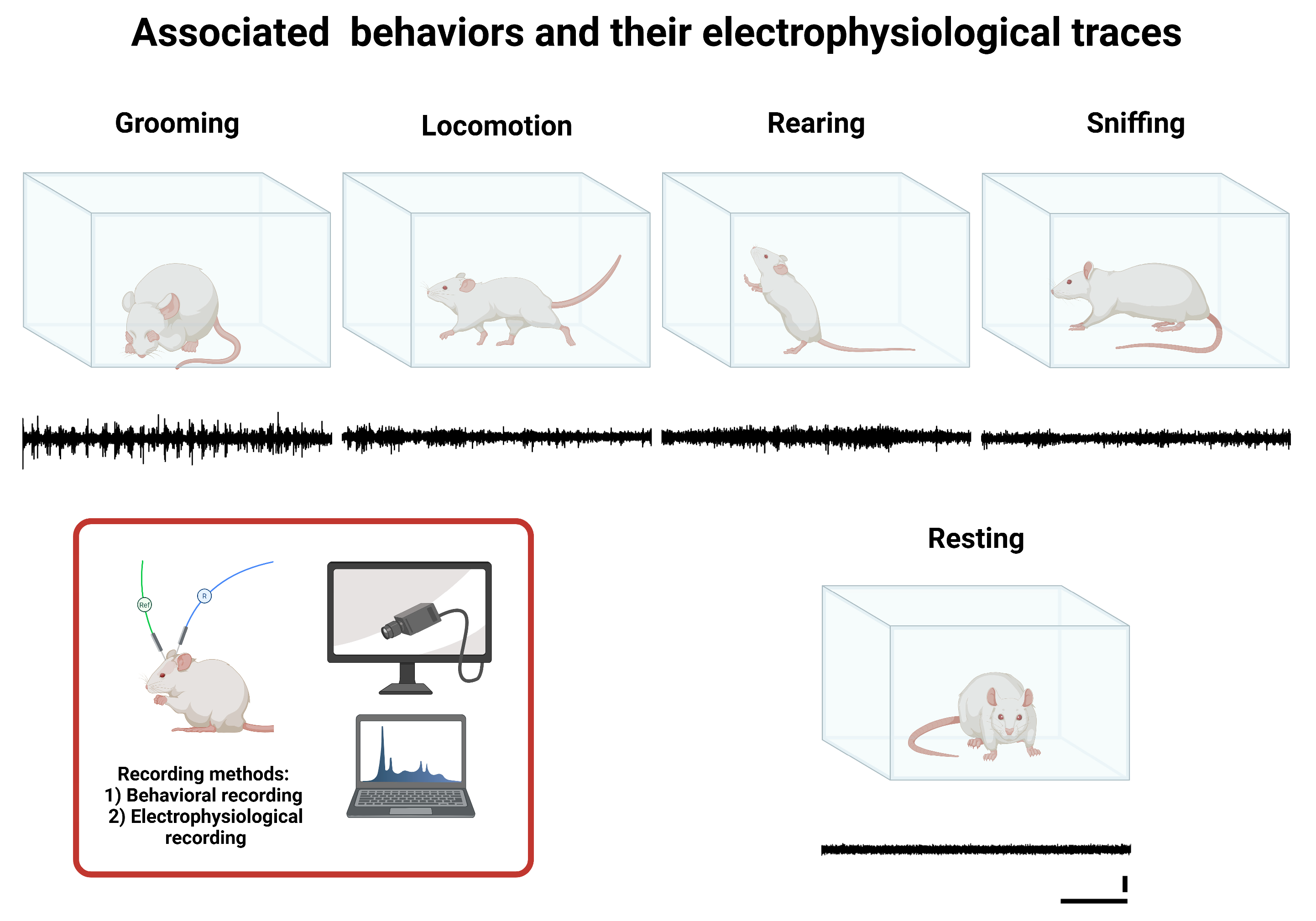
2.4. Associated Behaviors
- Resting: A basal state of inactivity in which the rat maintains a static and relaxed position without exhibiting voluntary or exploratory movements, indicating a low level of motor activation.
- Grooming: Self-care and hygiene behavior during which the rat uses its limbs, mouth, and tongue to clean and groom different body areas, such as the face, ears, limbs, and fur. This process may also have been associated with emotional state and stress levels.
- Locomotion (walking): Active horizontal movement involving the voluntary displacement of the rat through the environment. This behavior is characterized by the coordinated use of all four limbs without adopting upright postures or significant changes in body orientation.
- Rearing: Active vertical exploratory behavior in which the rat elevates itself by supporting its hind limbs, typically extending the forelimbs to examine the surroundings. This behavior is indicative of active environmental stimulus search and spatial evaluation.
- Sniffing: An exploratory action focused on the sense of smell when resting. The rat performs nasal movements to capture and analyze chemical stimuli in the environment, facilitating food identification, social signal detection, and threat assessment.
2.5. Collection of Data
2.5.1. Electrophysiological Recording
2.5.2. Behavioral Recording
2.6. Electrophysiological Analysis and Power Spectral Density
Spectral Analysis
- Area under the curve (AUC) calculation per frequency band:
- 2.
- Normalization by bandwidth:
2.7. Statistical Analysis
- Intergroup comparisons: The Mann–Whitney U test was applied for each measurement week to compare the control and experimental groups. This test was used to identify significant differences in each behavior between the two groups.
- Intragroup comparisons: The Friedman test was employed to evaluate global differences across the different measurement periods within each group. In cases where the Friedman test was significant, post hoc analyses were performed using the Wilcoxon tests with Holm adjustment to identify specific differences between weeks.
- Correlation analysis: A correlation analysis was conducted to explore the relationship between time and normalized PSD values. For this, a Spearman correlation test was used. This analysis was performed independently of brain structure and measurement week.
3. Results
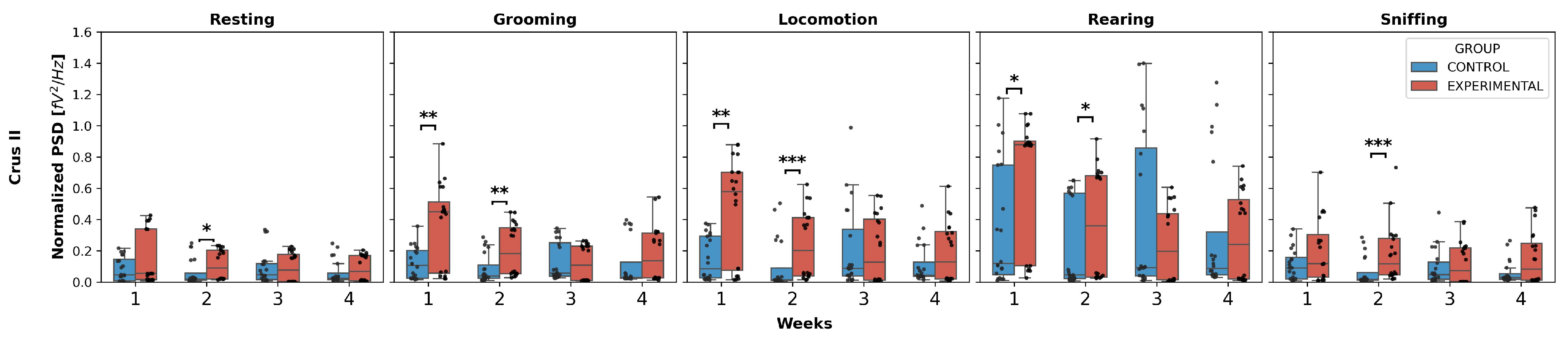
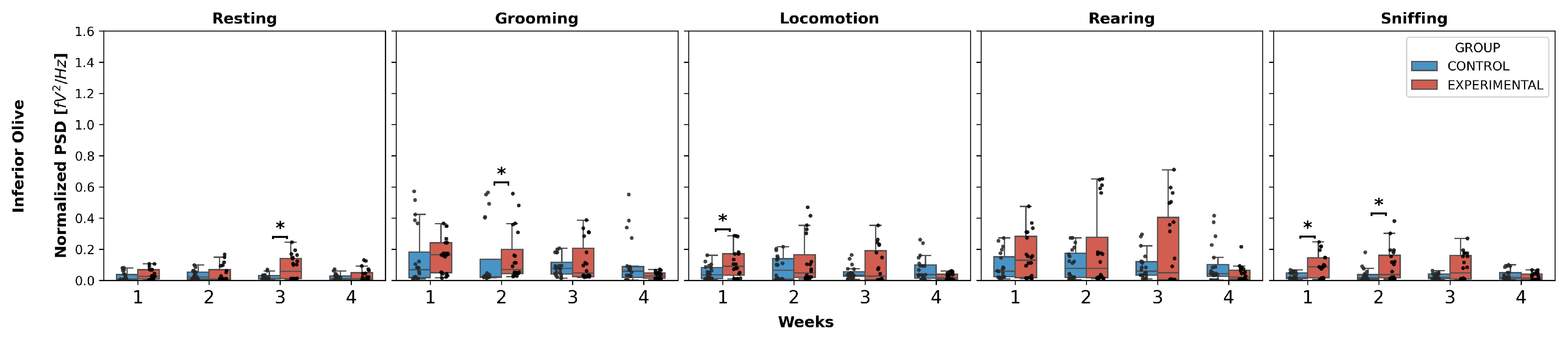
3.1. Intergroup Comparison
- Grooming: W1 (U = 96.0, p = 0.0051), W2 (U = 86.0, p = 0.0021);
- Locomotion: W1 (U = 100.0, p = 0.0071), W2 (U = 73.0, p = 0.0006);
- Rearing: W1 (U = 117.0, p = 0.0256), W2 (U = 119.0, p = 0.0294).
- Grooming: (U = 49.0, p < 0.0001);
- Locomotion: (U = 75.5, p = 0.0008);
- Sniffing: (U = 99.0, p = 0.0066).
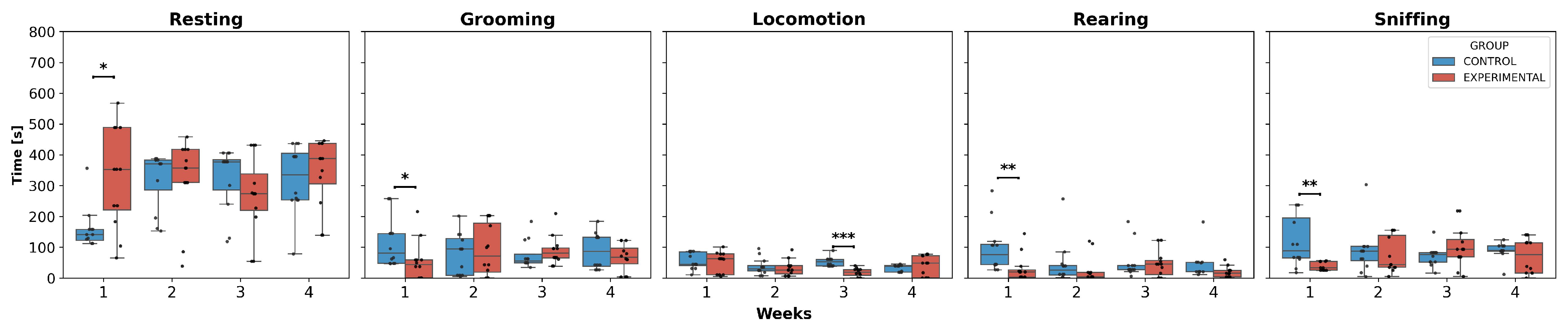
3.2. Intragroup Comparison
- Resting: W3 and W4 showed a decrease compared to W2.
- Grooming: W3 and W4 showed a decrease compared to W1, while in W3, this decrease was also observed compared to W2, and in W4, it showed an increase compared to W3.
- Locomotion: a decrease was observed in W2, W3, and W4 compared to W1.
- Rearing: W2, W3, and W4 showed a significant decrease compared to W1, a decrease in W3 and W4 compared to W2 was also observed, and W4 showed an increase compared to W3.
- Resting: W4 represented a decrease compared to W3.
- Grooming: W4 exhibited a significant decrease compared to W1, W2, and W3.
- Locomotion and rearing: a decreasing pattern was observed in W4, but only compared to W1 and W2, with no differences in the remaining weeks.
- Sniffing: W4 decreased compared to W1, W2, and W3, respectively.
- Resting: there was a decrease only in W4 compared to W3.
- Grooming, locomotion, and sniffing: W4 exhibited a significant decrease compared to W2.
3.3. Behavioral Analysis
3.4. Correlation Analysis
4. Discussion
5. Perspectives and Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP | Anteroposterior |
| AUC | Area Under the Curve |
| BG | Basal Ganglia |
| DBS | Deep Brain Stimulation |
| DN | Dentate Nucleus |
| DV | Dorsoventral |
| IO | Inferior Olive |
| ML | Mediolateral |
| MUA | Multiunit Activity |
| PD | Parkinson’s Disease |
| PSD | Power Spectral Density |
| VLS | Ventrolateral Striatum |
Appendix A
Appendix A.1. Median Values of Normalized PSD and Behavior Execution Time

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | Behavior | Week | Control [fV2/Hz] | Experimental [fV2/Hz] | ||||
|---|---|---|---|---|---|---|---|---|
| Median | Q1 | Q3 | Median | Q1 | Q3 | |||
| Crus II | Resting | 1 | 0.0460 | 0.0079 | 0.1452 | 0.0543 | 0.0158 | 0.3390 |
| 2 | 0.0187 | 0.0111 | 0.0579 | 0.0895 | 0.0195 | 0.2038 | ||
| 3 | 0.0463 | 0.0159 | 0.1181 | 0.0775 | 0.0025 | 0.1758 | ||
| 4 | 0.0241 | 0.0165 | 0.0569 | 0.0696 | 0.0081 | 0.1704 | ||
| Grooming | 1 | 0.1062 | 0.0245 | 0.2015 | 0.4488 | 0.0568 | 0.5124 | |
| 2 | 0.0399 | 0.0231 | 0.1105 | 0.1817 | 0.0533 | 0.3471 | ||
| 3 | 0.0572 | 0.0371 | 0.2513 | 0.1072 | 0.0100 | 0.2297 | ||
| 4 | 0.0304 | 0.0236 | 0.1269 | 0.1352 | 0.0247 | 0.3144 | ||
| Locomotion | 1 | 0.0844 | 0.0258 | 0.2938 | 0.5793 | 0.0738 | 0.7022 | |
| 2 | 0.0136 | 0.0093 | 0.0901 | 0.2016 | 0.0404 | 0.4122 | ||
| 3 | 0.0870 | 0.0394 | 0.3372 | 0.1269 | 0.0112 | 0.4024 | ||
| 4 | 0.0437 | 0.0356 | 0.1270 | 0.1301 | 0.0204 | 0.3227 | ||
| Rearing | 1 | 0.1192 | 0.0482 | 0.7493 | 0.8787 | 0.1040 | 0.9025 | |
| 2 | 0.0455 | 0.0223 | 0.5684 | 0.3583 | 0.0314 | 0.6795 | ||
| 3 | 0.0926 | 0.0389 | 0.8566 | 0.1967 | 0.0132 | 0.4365 | ||
| 4 | 0.0864 | 0.0456 | 0.3204 | 0.2417 | 0.0197 | 0.5270 | ||
| Sniffing | 1 | 0.0907 | 0.0206 | 0.1563 | 0.1171 | 0.0321 | 0.3034 | |
| 2 | 0.0163 | 0.0117 | 0.0611 | 0.1168 | 0.0448 | 0.2793 | ||
| 3 | 0.0478 | 0.0183 | 0.1281 | 0.0730 | 0.0020 | 0.2184 | ||
| 4 | 0.0289 | 0.0182 | 0.0533 | 0.0831 | 0.0115 | 0.2468 | ||
| Structure | Behavior | Week | Control [fV2/Hz] | Experimental [fV2/Hz] | ||||
|---|---|---|---|---|---|---|---|---|
| Median | Q1 | Q3 | Median | Q1 | Q3 | |||
| Inferior olive | Resting | 1 | 0.00969 | 0.00404 | 0.0382 | 0.026 | 0.01 | 0.0687 |
| 2 | 0.0218 | 0.0088 | 0.0522 | 0.00969 | 0.0084 | 0.0694 | ||
| 3 | 0.0115 | 0.00908 | 0.0307 | 0.0564 | 0.0117 | 0.14166 | ||
| 4 | 0.012 | 0.00478 | 0.029 | 0.0117 | 0.00264 | 0.05 | ||
| Grooming | 1 | 0.0685 | 0.0177 | 0.18222 | 0.164119 | 0.0493 | 0.241812 | |
| 2 | 0.0246 | 0.0203 | 0.135983 | 0.0717 | 0.0438 | 0.197312 | ||
| 3 | 0.0758 | 0.0412 | 0.116298 | 0.0473 | 0.0242 | 0.205878 | ||
| 4 | 0.0588 | 0.0207 | 0.0888 | 0.0293 | 0.0137 | 0.0469 | ||
| Locomotion | 1 | 0.0355 | 0.0124 | 0.0821 | 0.0912 | 0.0343 | 0.170791 | |
| 2 | 0.0649 | 0.0082 | 0.139841 | 0.0464 | 0.0194 | 0.16514 | ||
| 3 | 0.0334 | 0.0259 | 0.0544 | 0.0271 | 0.00386 | 0.190242 | ||
| 4 | 0.0365 | 0.013 | 0.0973 | 0.0159 | 0.00768 | 0.0398 | ||
| Rearing | 1 | 0.0591 | 0.0229 | 0.151807 | 0.129852 | 0.018 | 0.282026 | |
| 2 | 0.0767 | 0.0199 | 0.174694 | 0.0775 | 0.0176 | 0.275724 | ||
| 3 | 0.0593 | 0.0327 | 0.11991 | 0.0491 | 0.00321 | 0.404475 | ||
| 4 | 0.0435 | 0.0248 | 0.102217 | 0.0211 | 0.00745 | 0.0644 | ||
| Sniffing | 1 | 0.0193 | 0.0123 | 0.0482 | 0.0865 | 0.0156 | 0.146023 | |
| 2 | 0.0162 | 0.00815 | 0.0379 | 0.0382 | 0.0151 | 0.160667 | ||
| 3 | 0.0206 | 0.011 | 0.0409 | 0.0468 | 0.00948 | 0.159506 | ||
| 4 | 0.0203 | 0.00933 | 0.0486 | 0.0145 | 0.00375 | 0.0417 | ||
| Structure | Behavior | Week | Control [fV2/Hz] | Experimental [fV2/Hz] | ||||
|---|---|---|---|---|---|---|---|---|
| Median | Q1 | Q3 | Median | Q1 | Q3 | |||
| Dentate nucleus | Resting | 1 | 0.00636 | 0.00394 | 0.0189 | 0.00691 | 0.00584 | 0.021 |
| 2 | 0.007 | 0.00506 | 0.0119 | 0.0117 | 0.00544 | 0.0698 | ||
| 3 | 0.00642 | 0.00334 | 0.0192 | 0.00934 | 0.0022 | 0.0507 | ||
| 4 | 0.00479 | 0.00319 | 0.00528 | 0.00615 | 0.00167 | 0.0226 | ||
| Grooming | 1 | 0.0219 | 0.0169 | 0.0319 | 0.0238 | 0.0115 | 0.139797 | |
| 2 | 0.0209 | 0.014 | 0.029 | 0.0825 | 0.0341 | 0.14262 | ||
| 3 | 0.0354 | 0.0285 | 0.0447 | 0.0296 | 0.00697 | 0.0554 | ||
| 4 | 0.00964 | 0.00741 | 0.0179 | 0.0217 | 0.00805 | 0.0407 | ||
| Locomotion | 1 | 0.01 | 0.0086 | 0.0224 | 0.0123 | 0.00832 | 0.0485 | |
| 2 | 0.00866 | 0.00701 | 0.0106 | 0.0469 | 0.0126 | 0.123047 | ||
| 3 | 0.0219 | 0.0102 | 0.0317 | 0.0155 | 0.00246 | 0.0544 | ||
| 4 | 0.00602 | 0.00484 | 0.0158 | 0.0246 | 0.00516 | 0.0322 | ||
| Rearing | 1 | 0.0177 | 0.0129 | 0.0231 | 0.0164 | 0.0136 | 0.0823 | |
| 2 | 0.0175 | 0.0134 | 0.0198 | 0.0681 | 0.0104 | 0.13338 | ||
| 3 | 0.0294 | 0.0204 | 0.0326 | 0.02 | 0.00276 | 0.0952 | ||
| 4 | 0.00811 | 0.00538 | 0.0288 | 0.0163 | 0.00377 | 0.0638 | ||
| Sniffing | 1 | 0.018 | 0.00971 | 0.0262 | 0.012 | 0.00914 | 0.0667 | |
| 2 | 0.00878 | 0.00669 | 0.0113 | 0.0214 | 0.00944 | 0.0668 | ||
| 3 | 0.0153 | 0.00733 | 0.0182 | 0.0126 | 0.00177 | 0.0472 | ||
| 4 | 0.00657 | 0.00387 | 0.0116 | 0.00616 | 0.00232 | 0.0257 | ||
| Behavior | Week | Control [s] | Experimental [s] | ||||
|---|---|---|---|---|---|---|---|
| Median | Q1 | Q3 | Median | Q1 | Q3 | ||
| Resting | 1 | 141.15 | 122.23 | 157.90 | 352.80 | 221.68 | 488.50 |
| 2 | 370.70 | 285.76 | 383.15 | 356.95 | 309.80 | 417.40 | |
| 3 | 377.05 | 285.35 | 384.26 | 273.55 | 219.68 | 338.51 | |
| 4 | 334.92 | 253.10 | 404.83 | 387.70 | 305.84 | 437.00 | |
| Grooming | 1 | 80.53 | 47.45 | 144.60 | 43.23 | 0.00 | 59.00 |
| 2 | 94.25 | 9.00 | 127.95 | 71.00 | 18.90 | 177.85 | |
| 3 | 55.72 | 49.15 | 77.73 | 81.10 | 65.25 | 97.88 | |
| 4 | 86.98 | 38.54 | 132.46 | 66.80 | 45.69 | 96.45 | |
| Locomotion | 1 | 44.70 | 40.41 | 85.04 | 62.60 | 10.60 | 78.05 |
| 2 | 30.00 | 22.96 | 40.46 | 25.70 | 14.23 | 40.05 | |
| 3 | 52.55 | 39.39 | 60.55 | 18.58 | 8.59 | 27.45 | |
| 4 | 38.40 | 18.70 | 40.60 | 48.60 | 0.00 | 72.00 | |
| Rearing | 1 | 75.30 | 43.70 | 109.53 | 19.65 | 3.20 | 26.11 |
| 2 | 25.43 | 9.53 | 40.73 | 3.70 | 2.55 | 18.40 | |
| 3 | 27.30 | 27.30 | 40.60 | 45.25 | 11.63 | 56.64 | |
| 4 | 20.90 | 20.45 | 51.40 | 15.40 | 3.30 | 24.55 | |
| Sniffing | 1 | 88.05 | 65.43 | 194.66 | 33.20 | 25.54 | 53.95 |
| 2 | 87.25 | 56.30 | 102.50 | 43.50 | 34.60 | 138.31 | |
| 3 | 76.63 | 52.05 | 82.60 | 93.25 | 68.60 | 124.55 | |
| 4 | 88.95 | 87.00 | 104.30 | 75.90 | 16.00 | 114.56 | |
Appendix A.2. PSD Statistical Test Values for Intragroup Comparison
| Structure | Behavior | Friedman Statistical Value | Wilcoxon Statistical Value by Weeks Compared | |||||
|---|---|---|---|---|---|---|---|---|
| (1, 2) | (1, 3) | (1, 4) | (2, 3) | (2, 4) | (3, 4) | |||
| Crus II | Resting | 18.78 | 87 | 56 | 56 | 21 | 16 | 102 |
| Grooming | 48.48 | 28 | 0 | 10 | 0 | 37 | 0 | |
| Locomotion | 29.70 | 16 | 0 | 4.5 | 54 | 50 | 92 | |
| Rearing | 54.18 | 1 | 0 | 0 | 0 | 11 | 18 | |
| Sniffing | 15.18 | 102 | 50 | 64 | 56 | 65 | 57 | |
| Structure | Behavior | Friedman p-Value | Wilcoxon p-Value by Weeks Compared | |||||
|---|---|---|---|---|---|---|---|---|
| (1, 2) | (1, 3) | (1, 4) | (2, 3) | (2, 4) | (3, 4) | |||
| Crus II | Resting | 0.0003036 | 1.0 | 1.0 | 1.0 | 0.0280724 | 0.0116043 | 1.0 |
| Grooming | 0.0000000 | 0.0840797 | 0.0000916 | 0.0031986 | 0.0000916 | 0.2641983 | 0.0000916 | |
| Locomotion | 0.0000016 | 0.0116043 | 0.0000916 | 0.0007629 | 1.0 | 0.8797684 | 1.0 | |
| Rearing | 0.0000000 | 0.0001564 | 0.0000916 | 0.0000916 | 0.0000916 | 0.0039864 | 0.0164070 | |
| Sniffing | 0.0016691 | 1.0 | 0.8797684 | 1.0 | 1.0 | 1.0 | 1.0 | |
| Structure | Behavior | Friedman Statistical Value | Wilcoxon Statistical Value by Weeks Compared | |||||
|---|---|---|---|---|---|---|---|---|
| (1, 2) | (1, 3) | (1, 4) | (2, 3) | (2, 4) | (3, 4) | |||
| Inferior olive | Resting | 21.30 | 93 | 43 | 88 | 43 | 48 | 23 |
| Grooming | 42.54 | 92 | 52 | 0 | 19 | 0 | 0 | |
| Locomotion | 27.42 | 93 | 75 | 5 | 54 | 0 | 57 | |
| Rearing | 23.40 | 69 | 87 | 10 | 73 | 13 | 53 | |
| Sniffing | 34.90 | 86 | 96 | 0 | 52 | 1 | 0 | |
| Structure | Behavior | Friedman p-Value | Wilcoxon p-Value by Weeks Compared | |||||
|---|---|---|---|---|---|---|---|---|
| (1, 2) | (1, 3) | (1, 4) | (2, 3) | (2, 4) | (3, 4) | |||
| Inferior olive | Resting | 0.0000912 | 1.0 | 0.4423752 | 1.0 | 0.4423752 | 0.6553650 | 0.0302315 |
| Grooming | 0.0000000 | 1.0 | 0.9203777 | 0.0000687 | 0.0152245 | 0.0000687 | 0.0000687 | |
| Locomotion | 0.0000048 | 1.0 | 1.0 | 0.0005531 | 0.9903870 | 0.0000687 | 1.0 | |
| Rearing | 0.0000333 | 1.0 | 1.0 | 0.0022964 | 1.0 | 0.0045319 | 0.9570465 | |
| Sniffing | 0.0000001 | 1.0 | 1.0 | 0.0000687 | 1.0 | 0.0001144 | 0.0000687 | |
| Structure | Behavior | Friedman Statistical Value | Wilcoxon Statistical Value by Weeks Compared | |||||
|---|---|---|---|---|---|---|---|---|
| (1, 2) | (1, 3) | (1, 4) | (2, 3) | (2, 4) | (3, 4) | |||
| Dentate nucleus | Resting | 12.12 | 59 | 65 | 72 | 87 | 42 | 9 |
| Grooming | 22.02 | 60 | 104 | 54.5 | 25 | 0 | 35 | |
| Locomotion | 13.14 | 56 | 93 | 97 | 49 | 17 | 81 | |
| Rearing | 12.78 | 51 | 60 | 96 | 70 | 31 | 87 | |
| Sniffing | 20.22 | 62 | 83 | 36 | 52 | 0 | 51 | |
| Structure | Behavior | Friedman p-Value | Wilcoxon p-Value by Weeks Compared | |||||
|---|---|---|---|---|---|---|---|---|
| (1, 2) | (1, 3) | (1, 4) | (2, 3) | (2, 4) | (3, 4) | |||
| Dentate nucleus | Resting | 0.0069832 | 1.0 | 1.0 | 1.0 | 1.0 | 0.5841675 | 0.0032101 |
| Grooming | 0.0000646 | 1.0 | 1.0 | 1.0 | 0.0692863 | 0.0001030 | 0.2626419 | |
| Locomotion | 0.0043434 | 1.0 | 1.0 | 1.0 | 1.0 | 0.0185566 | 1.0 | |
| Rearing | 0.0051373 | 1.0 | 1.0 | 1.0 | 1.0 | 0.1603966 | 1.0 | |
| Sniffing | 0.0001528 | 1.0 | 1.0 | 0.2907944 | 1.0 | 0.0001030 | 1.0 | |
References
- Keener, A.; Bordelon, Y. Parkinsonism. Semin. Neurol. 2016, 36, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, E.; Wenning, G.; Poewe, W. The Diagnosis of Parkinson’s Disease. Lancet Neurol. 2006, 5, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Höllerhage, M. Secondary Parkinsonism Due to Drugs, Vascular Lesions, Tumors, Trauma, and Other Insults. Int. Rev. Neurobiol. 2019, 149, 377–418. [Google Scholar]
- Viveros-Martínez, I.; Zarate-Calderon, C.; Chi-Castañeda, D.; Carrillo, P.; Aranda-Abreu, G.E.; Martínez, A.J.; Manzo, J.; Coria, G.A.; García, L.I. Characterizing Secondary and Atypical Parkinsonisms: Defining Features and Clinical Variability. Neuroglia 2024, 5, 467–487. [Google Scholar] [CrossRef]
- Hayes, M.T. Parkinson’s Disease and Parkinsonism. Am. J. Med. 2019, 132, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, T. Changing Views of the Pathophysiology of Parkinsonism. Mov. Disord. 2019, 34, 1130–1143. [Google Scholar] [CrossRef]
- Li, R.; Zou, T.; Wang, X.; Wang, H.; Hu, X.; Xie, F.; Meng, L.; Chen, H. Basal Ganglia Atrophy–Associated Causal Structural Network Degeneration in Parkinson’s Disease. Hum. Brain Mapp. 2022, 43, 1145–1156. [Google Scholar] [CrossRef]
- Franco, G.; Lazzeri, G.; Di Fonzo, A. Parkinsonism and Ataxia. J. Neurol. Sci. 2022, 433, 120020. [Google Scholar] [CrossRef]
- Kemaladina, I.; Gazali, S.; Fadhila, L.; Amra, K. Characteristics of Parkinson’s Disease Cardinal Symptoms in a Neurology Outpatient Clinic. J. Neurol. Sci. 2023, 455, 121767. [Google Scholar] [CrossRef]
- Shin, H.-W.; Hong, S.-W.; Youn, Y.C. Clinical Aspects of the Differential Diagnosis of Parkinson’s Disease and Parkinsonism. J. Clin. Neurol. 2022, 18, 259. [Google Scholar] [CrossRef]
- Moustafa, A.A.; Chakravarthy, S.; Phillips, J.R.; Gupta, A.; Keri, S.; Polner, B.; Frank, M.J.; Jahanshahi, M. Motor Symptoms in Parkinson’s Disease: A Unified Framework. Neurosci. Biobehav. Rev. 2016, 68, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Fahn, S. Classification of Movement Disorders. Mov. Disord. 2011, 26, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Sarasso, E.; Gardoni, A.; Zenere, L.; Emedoli, D.; Balestrino, R.; Grassi, A.; Basaia, S.; Tripodi, C.; Canu, E.; Malcangi, M.; et al. Neural Correlates of Bradykinesia in Parkinson’s Disease: A Kinematic and Functional MRI Study. NPJ Park. Dis. 2024, 10, 167. [Google Scholar] [CrossRef]
- Bologna, M.; Espay, A.J.; Fasano, A.; Paparella, G.; Hallett, M.; Berardelli, A. Redefining Bradykinesia. Mov. Disord. 2023, 38, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Herz, D.M.; Brown, P. Moving, Fast and Slow: Behavioural Insights into Bradykinesia in Parkinson’s Disease. Brain 2023, 146, 3576–3586. [Google Scholar] [CrossRef]
- Baradaran, N.; Tan, S.N.; Liu, A.; Ashoori, A.; Palmer, S.J.; Wang, Z.J.; Oishi, M.M.K.; McKeown, M.J. Parkinson’s Disease Rigidity: Relation to Brain Connectivity and Motor Performance. Front. Neurol. 2013, 4, 67. [Google Scholar] [CrossRef]
- Abusrair, A.H.; Elsekaily, W.; Bohlega, S. Tremor in Parkinson’s Disease: From Pathophysiology to Advanced Therapies. Tremor Other Hyperkinetic Mov. 2022, 12, 29. [Google Scholar] [CrossRef]
- Pan, M.-K.; Ni, C.-L.; Wu, Y.-C.; Li, Y.-S.; Kuo, S.-H. Animal Models of Tremor: Relevance to Human Tremor Disorders. Tremor Other Hyperkinetic Mov. 2018, 8, 587. [Google Scholar] [CrossRef]
- Salamone, J.D.; Mayorga, A.J.; Trevitt, J.T.; Cousins, M.S.; Conlan, A.; Nawab, A. Tremulous Jaw Movements in Rats:A Model of Parkinsonian Tremor. Prog. Neurobiol. 1998, 56, 591–611. [Google Scholar] [CrossRef]
- Waku, I.; Magalhães, M.S.; Alves, C.O.; de Oliveira, A.R. Haloperidol-induced Catalepsy as an Animal Model for Parkinsonism: A Systematic Review of Experimental Studies. Eur. J. Neurosci. 2021, 53, 3743–3767. [Google Scholar] [CrossRef]
- Herrera-Meza, G.; Manzo, J.; Hernández, M.E.; Miquel, M.; García, L.I. Induction of Mandibular Tremor Using Electrolytic Lesion of the Ventrolateral Striatum or Using Subchronic Haloperidol Therapy in Male Rats: An Electromyographic Comparison. Neurol. (Engl. Ed.) 2014, 29, 416–422. [Google Scholar] [CrossRef]
- Vásquez-Celaya, L.; Marín, G.; Hernández, M.E.; Carrillo, P.; Pérez, C.A.; Coria-Avila, G.A.; Manzo, J.; Miquel, M.; García, L.I. Functional Correlation between Cerebellum and Basal Ganglia: A Parkinsonism Model. Neurología 2024, 39, 555–563. [Google Scholar] [CrossRef]
- Vásquez-Celaya, L.; Marín-Márquez, G.; Manzo, J.; Carrillo-Castilla, P.; Martínez, A.J.; Ortiz Pulido, R.; Zempoalteca Ramírez, R.; Coria-Avila, G.A.; García, L.I. Electrophysiological Characterization of Cerebellar Responses during Exploration and Grooming Behaviors in a Rat Model of Parkinsonism. Brain Sci. 2023, 13, 537. [Google Scholar] [CrossRef] [PubMed]
- Bologna, M.; Paparella, G.; Fasano, A.; Hallett, M.; Berardelli, A. Evolving Concepts on Bradykinesia. Brain 2020, 143, 727–750. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.M.; Galley, S.; Johnson, S.; Stevenson, J.; Huang, X.; McKeown, M.J. The Role of the Cerebellum in the Pathophysiology of Parkinson’s Disease. Can. J. Neurol. Sci. J. Can. Des Sci. Neurologiques 2013, 40, 299–306. [Google Scholar] [CrossRef]
- Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Norma Oficial Mexicana NOM-062-ZOO-1999, Especificaciones Técnicas para la Producción, Cuidado y uso de Animales de Laboratorio. Available online: https://www.gob.mx/cms/uploads/attachment/file/203498/NOM-062-ZOO-1999_220801.pdf (accessed on 20 March 2025).
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Cetinkaya, E.; Lang, E.J.; Sahin, M. Sensorimotor Content of Multi-Unit Activity Recorded in the Paramedian Lobule of the Cerebellum Using Carbon Fiber Microelectrode Arrays. Front. Neurosci. 2024, 18, 1232653. [Google Scholar] [CrossRef]
- Yang, A.I.; Vanegas, N.; Lungu, C.; Zaghloul, K.A. Beta-Coupled High-Frequency Activity and Beta-Locked Neuronal Spiking in the Subthalamic Nucleus of Parkinson’s Disease. J. Neurosci. 2014, 34, 12816–12827. [Google Scholar] [CrossRef]
- Mattia, M.; Ferraina, S.; Del Giudice, P. Dissociated Multi-Unit Activity and Local Field Potentials: A Theory Inspired Analysis of a Motor Decision Task. Neuroimage 2010, 52, 812–823. [Google Scholar] [CrossRef]
- Rey, H.G.; Pedreira, C.; Quian Quiroga, R. Past, Present and Future of Spike Sorting Techniques. Brain Res. Bull. 2015, 119, 106–117. [Google Scholar] [CrossRef]
- Andrianarivelo, A.; Stein, H.; Gabillet, J.; Batifol, C.; Jalil, A.; Cayco Gajic, N.A.; Graupner, M. Cerebellar Interneuron Activity Is Triggered by Reach Endpoint during Learning of a Complex Locomotor Task. bioRxiv 2023. [Google Scholar] [CrossRef]
- Yu, H.; Sternad, D.; Corcos, D.M.; Vaillancourt, D.E. Role of Hyperactive Cerebellum and Motor Cortex in Parkinson’s Disease. Neuroimage 2007, 35, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Bostan, A.C.; Strick, P.L. The Basal Ganglia and the Cerebellum: Nodes in an Integrated Network. Nat. Rev. Neurosci. 2018, 19, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Tok, S.; Ahnaou, A.; Drinkenburg, W. Functional Neurophysiological Biomarkers of Early-Stage Alzheimer’s Disease: A Perspective of Network Hyperexcitability in Disease Progression. J. Alzheimer’s Dis. 2022, 88, 809–836. [Google Scholar] [CrossRef] [PubMed]
- Latorre, A.; Rocchi, L.; Paparella, G.; Manzo, N.; Bhatia, K.P.; Rothwell, J.C. Changes in Cerebellar Output Abnormally Modulate Cortical Myoclonus Sensorimotor Hyperexcitability. Brain 2024, 147, 1412–1422. [Google Scholar] [CrossRef]
- Kopf, M.; Martini, J.; Stier, C.; Ethofer, S.; Braun, C.; Li Hegner, Y.; Focke, N.K.; Marquetand, J.; Helfrich, R.F. Aperiodic Activity Indexes Neural Hyperexcitability in Generalized Epilepsy. eNeuro 2024, 11, ENEURO.0242-24.2024. [Google Scholar] [CrossRef]
- Li, T.; Le, W.; Jankovic, J. Linking the Cerebellum to Parkinson Disease: An Update. Nat. Rev. Neurol. 2023, 19, 645–654. [Google Scholar] [CrossRef]
- Llinás, R.R. Inferior Olive Oscillation as the Temporal Basis for Motricity and Oscillatory Reset as the Basis for Motor Error Correction. Neuroscience 2009, 162, 797–804. [Google Scholar] [CrossRef]
- Mitoma, H.; Kakei, S.; Tanaka, H.; Manto, M. Morphological and Functional Principles Governing the Plasticity Reserve in the Cerebellum: The Cortico-Deep Cerebellar Nuclei Loop Model. Biology 2023, 12, 1435. [Google Scholar] [CrossRef]
- Luque, N.R.; Garrido, J.A.; Carrillo, R.R.; D’Angelo, E.; Ros, E. Fast Convergence of Learning Requires Plasticity between Inferior Olive and Deep Cerebellar Nuclei in a Manipulation Task: A Closed-Loop Robotic Simulation. Front. Comput. Neurosci. 2014, 8, 97. [Google Scholar] [CrossRef]
- Winslow, B.D.; Tresco, P.A. Quantitative Analysis of the Tissue Response to Chronically Implanted Microwire Electrodes in Rat Cortex. Biomaterials 2010, 31, 1558–1567. [Google Scholar] [CrossRef]
- Polikov, V.S.; Tresco, P.A.; Reichert, W.M. Response of Brain Tissue to Chronically Implanted Neural Electrodes. J. Neurosci. Methods 2005, 148, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Duret, L.C.; Nagoshi, E. The Intertwined Relationship between Circadian Dysfunction and Parkinson’s Disease. Trends Neurosci. 2025, 48, 62–76. [Google Scholar] [CrossRef]
- Kozai, T.D.Y.; Jaquins-Gerstl, A.S.; Vazquez, A.L.; Michael, A.C.; Cui, X.T. Brain Tissue Responses to Neural Implants Impact Signal Sensitivity and Intervention Strategies. ACS Chem. Neurosci. 2015, 6, 48–67. [Google Scholar] [CrossRef]
- Keren-Happuch, E.; Chen, S.A.; Ho, M.R.; Desmond, J.E. A Meta-analysis of Cerebellar Contributions to Higher Cognition from PET and FMRI Studies. Hum. Brain Mapp. 2014, 35, 593–615. [Google Scholar]
- Habas, C. Functional Connectivity of the Cognitive Cerebellum. Front. Syst. Neurosci. 2021, 15, 642225. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Sakurai, Y. Inactivation of Cerebellar Cortical Crus II Disrupts Temporal Processing of Absolute Timing but Not Relative Timing in Voluntary Movements. Front. Syst. Neurosci. 2016, 10, 00016. [Google Scholar] [CrossRef]
- Zhu, J.; Hasanbegović, H.; Liu, L.D.; Gao, Z.; Li, N. Activity Map of a Cortico-Cerebellar Loop Underlying Motor Planning. Nat. Neurosci. 2023, 26, 1916–1928. [Google Scholar] [CrossRef] [PubMed]
- DeAngelo, V.; Gehan, A.; Paliwal, S.; Ho, K.; Hilliard, J.D.; Chiang, C.-H.; Viventi, J.; McConnell, G.C. Cerebellar Activity in Hemi-Parkinsonian Rats during Volitional Gait and Freezing. Brain Commun. 2024, 6, fcae246. [Google Scholar] [CrossRef]
- Ruigrok, T.J.H.; Wang, X.; Sabel-Goedknegt, E.; Coulon, P.; Gao, Z. A Disynaptic Basal Ganglia Connection to the Inferior Olive: Potential for Basal Ganglia Influence on Cerebellar Learning. Front. Syst. Neurosci. 2023, 17, 1176126. [Google Scholar] [CrossRef]
- Bhuvanasundaram, R.; Washburn, S.; Krzyspiak, J.; Khodakhah, K. Zona Incerta Modulation of the Inferior Olive and the Pontine Nuclei. Netw. Neurosci. 2024, 8, 260–274. [Google Scholar] [CrossRef]
- Chen, X.Y.; Wang, Y.; Chen, Y.; Chen, L.; Wolpaw, J.R. The Inferior Olive Is Essential for Long-Term Maintenance of a Simple Motor Skill. J. Neurophysiol. 2016, 116, 1946–1955. [Google Scholar] [CrossRef] [PubMed]
- Ezra-Nevo, G.; Volk, N.; Ramot, A.; Kuehne, C.; Tsoory, M.; Deussing, J.; Chen, A. Inferior Olive CRF Plays a Role in Motor Performance under Challenging Conditions. Transl. Psychiatry 2018, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, I.T.; Hoang, H.; Schweighofer, N.; Kawato, M. Adaptive Coupling of Inferior Olive Neurons in Cerebellar Learning. Neural Netw. 2013, 47, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Hartstone, W.G.; Brown, M.H.; Kelly, G.C.; Tate, W.J.; Kuo, S.; Dwork, A.J.; Louis, E.D.; Faust, P.L. Dentate Nucleus Neuronal Density: A Postmortem Study of Essential Tremor Versus Control Brains. Mov. Disord. 2021, 36, 995–999. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Gan, C.; Cao, X.; Ji, M.; Sun, H.; Yuan, Y.; Zhang, K. Altered Functional Connectivity of Cerebellar Dentate Nucleus in Peak-Dose Dyskinesia in Parkinson’s Disease. Front. Aging Neurosci. 2022, 14, 943179. [Google Scholar] [CrossRef]
- He, N.; Huang, P.; Ling, H.; Langley, J.; Liu, C.; Ding, B.; Huang, J.; Xu, H.; Zhang, Y.; Zhang, Z.; et al. Dentate Nucleus Iron Deposition Is a Potential Biomarker for Tremor-dominant Parkinson’s Disease. NMR Biomed. 2017, 30, e3554. [Google Scholar] [CrossRef]
- Ma, H.; Chen, H.; Fang, J.; Gao, L.; Ma, L.; Wu, T.; Hou, Y.; Zhang, J.; Feng, T. Resting-State Functional Connectivity of Dentate Nucleus Is Associated with Tremor in Parkinson’s Disease. J. Neurol. 2015, 262, 2247–2256. [Google Scholar] [CrossRef]
- Jolicoeur, F.B.; Rivest, R.; Drumheller, A. Hypokinesia, Rigidity, and Tremor Induced by Hypothalamic 6-OHDA Lesions in the Rat. Brain Res. Bull. 1991, 26, 317–320. [Google Scholar] [CrossRef]
- Jiang, X.; Yang, J.; Wang, Z.; Jia, J.; Wang, G. Functional Interaction of Abnormal Beta and Gamma Oscillations on Bradykinesia in Parkinsonian Rats. Brain Res. Bull. 2024, 209, 110911. [Google Scholar] [CrossRef]
- Yoshida, J.; Oñate, M.; Khatami, L.; Vera, J.; Nadim, F.; Khodakhah, K. Cerebellar Contributions to the Basal Ganglia Influence Motor Coordination, Reward Processing, and Movement Vigor. J. Neurosci. 2022, 42, 8406–8415. [Google Scholar] [CrossRef]
- Lyons, J.L.; Cohen, A.B. Selective Cerebellar and Basal Ganglia Injury in Neuroleptic Malignant Syndrome. J. Neuroimaging 2013, 23, 240–241. [Google Scholar] [CrossRef] [PubMed]
- Contreras-López, R.; Alatriste-León, H.; Díaz-Hernández, E.; Ramírez-Jarquín, J.O.; Tecuapetla, F. The Deep Cerebellar Nuclei to Striatum Disynaptic Connection Contributes to Skilled Forelimb Movement. Cell Rep. 2023, 42, 112000. [Google Scholar] [CrossRef]
- Middleton, S.J.; Racca, C.; Cunningham, M.O.; Traub, R.D.; Monyer, H.; Knöpfel, T.; Schofield, I.S.; Jenkins, A.; Whittington, M.A. High-Frequency Network Oscillations in Cerebellar Cortex. Neuron 2008, 58, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Milardi, D.; Quartarone, A.; Bramanti, A.; Anastasi, G.; Bertino, S.; Basile, G.A.; Buonasera, P.; Pilone, G.; Celeste, G.; Rizzo, G.; et al. The Cortico-Basal Ganglia-Cerebellar Network: Past, Present and Future Perspectives. Front. Syst. Neurosci. 2019, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Jha, M.K.; Lee, W.-H.; Suk, K. Functional Polarization of Neuroglia: Implications in Neuroinflammation and Neurological Disorders. Biochem. Pharmacol. 2016, 103, 1–16. [Google Scholar] [CrossRef]
- Castillo-Rangel, C.; Marin, G.; Hernández-Contreras, K.A.; Vichi-Ramírez, M.M.; Zarate-Calderon, C.; Torres-Pineda, O.; Diaz-Chiguer, D.L.; De la Mora González, D.; Gómez Apo, E.; Teco-Cortes, J.A.; et al. Neuroinflammation in Parkinson’s Disease: From Gene to Clinic: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 5792. [Google Scholar] [CrossRef]
- Harry, G.J. Microglia during Development and Aging. Pharmacol. Ther. 2013, 139, 313–326. [Google Scholar] [CrossRef]
- Herrera-Meza, G.; Manzo, J.; Hernández, M.E.; Miquel, M.; García, L.I. Inducción Del Temblor Mandibular Por Lesión Electrolítica Del Estriado Ventrolateral y Por El Tratamiento Subcrónico Con Haloperidol En Rata Macho: Un Contraste Electromiográfico. Neurología 2014, 29, 416–422. [Google Scholar] [CrossRef]
- Hariz, M.; Blomstedt, P. Deep Brain Stimulation for Parkinson’s Disease. J. Intern. Med. 2022, 292, 764–778. [Google Scholar] [CrossRef]
- Miterko, L.N.; Lin, T.; Zhou, J.; van der Heijden, M.E.; Beckinghausen, J.; White, J.J.; Sillitoe, R.V. Neuromodulation of the Cerebellum Rescues Movement in a Mouse Model of Ataxia. Nat. Commun. 2021, 12, 1295. [Google Scholar] [CrossRef]
- Tai, C.-H.; Tseng, S.-H. Cerebellar Deep Brain Stimulation for Movement Disorders. Neurobiol. Dis. 2022, 175, 105899. [Google Scholar] [CrossRef] [PubMed]
- Grobe-Einsler, M.; Baljasnikowa, V.; Faikus, A.; Schaprian, T.; Kaut, O. Cerebellar Transcranial Magnetic Stimulation Improves Motor Function in Parkinson’s Disease. Ann. Clin. Transl. Neurol. 2024, 11, 2673–2684. [Google Scholar] [CrossRef] [PubMed]
- Workman, C.D.; Fietsam, A.C.; Uc, E.Y.; Rudroff, T. Cerebellar Transcranial Direct Current Stimulation in People with Parkinson’s Disease: A Pilot Study. Brain Sci. 2020, 10, 96. [Google Scholar] [CrossRef]
- Milardi, D.; Arrigo, A.; Anastasi, G.; Cacciola, A.; Marino, S.; Mormina, E.; Calamuneri, A.; Bruschetta, D.; Cutroneo, G.; Trimarchi, F.; et al. Extensive Direct Subcortical Cerebellum-Basal Ganglia Connections in Human Brain as Revealed by Constrained Spherical Deconvolution Tractography. Front. Neuroanat. 2016, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Solstrand Dahlberg, L.; Lungu, O.; Doyon, J. Cerebellar Contribution to Motor and Non-Motor Functions in Parkinson’s Disease: A Meta-Analysis of FMRI Findings. Front. Neurol. 2020, 11, 127. [Google Scholar] [CrossRef]
- Radhakrishnan, V.; Gallea, C.; Valabregue, R.; Krishnan, S.; Kesavadas, C.; Thomas, B.; James, P.; Menon, R.; Kishore, A. Cerebellar and Basal Ganglia Structural Connections in Humans: Effect of Aging and Relation with Memory and Learning. Front. Aging Neurosci. 2023, 15, 1019239. [Google Scholar] [CrossRef]


| Structure | AP [mm] | ML [mm] | DV [mm] |
|---|---|---|---|
| VLS | −0.48 | ±4.40 | −6.80 |
| Crus II | −14.00 | 3.40 | −5.00 |
| IO | −11.80 | 0.80 | −11.00 |
| DN | −11.30 | 3.40 | −6.40 |
| Behavior | Structure | ||
|---|---|---|---|
| Crus II | IO | DN | |
| Resting | −0.038235294 | −0.05 | −0.241176471 |
| Grooming | −0.6136867 | −0.566593237 | −0.644591786 |
| Locomotion | 0.380532629 | 0.286136938 | 0.333334784 |
| Rearing | 0.085546095 | 0.209440439 | 0.292036669 |
| Sniffing | 0.579411765 | 0.658823529 | 0.579411765 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarate-Calderon, C.; Marín, G.; Viveros-Martínez, I.; Vásquez-Celaya, L.; Carrillo-Castilla, P.; Aranda-Abreu, G.E.; Chi-Castañeda, D.; García, L.I. Cerebellar Contributions to Hypokinetic Symptoms in an Acute Lesion Parkinsonism Model. Neurol. Int. 2025, 17, 72. https://doi.org/10.3390/neurolint17050072
Zarate-Calderon C, Marín G, Viveros-Martínez I, Vásquez-Celaya L, Carrillo-Castilla P, Aranda-Abreu GE, Chi-Castañeda D, García LI. Cerebellar Contributions to Hypokinetic Symptoms in an Acute Lesion Parkinsonism Model. Neurology International. 2025; 17(5):72. https://doi.org/10.3390/neurolint17050072
Chicago/Turabian StyleZarate-Calderon, Cristofer, Gerardo Marín, Iraís Viveros-Martínez, Lizbeth Vásquez-Celaya, Porfirio Carrillo-Castilla, Gonzalo E. Aranda-Abreu, Donaji Chi-Castañeda, and Luis I. García. 2025. "Cerebellar Contributions to Hypokinetic Symptoms in an Acute Lesion Parkinsonism Model" Neurology International 17, no. 5: 72. https://doi.org/10.3390/neurolint17050072
APA StyleZarate-Calderon, C., Marín, G., Viveros-Martínez, I., Vásquez-Celaya, L., Carrillo-Castilla, P., Aranda-Abreu, G. E., Chi-Castañeda, D., & García, L. I. (2025). Cerebellar Contributions to Hypokinetic Symptoms in an Acute Lesion Parkinsonism Model. Neurology International, 17(5), 72. https://doi.org/10.3390/neurolint17050072