
Estrogen-Mediated Neural Mechanisms of Sex Differences in Burning Mouth Syndrome


Abstract
1. Introduction
2. Methods
3. Three Pain Neural Circuits of BMS
4. Sex Differences in the Peripheral Pain Circuit
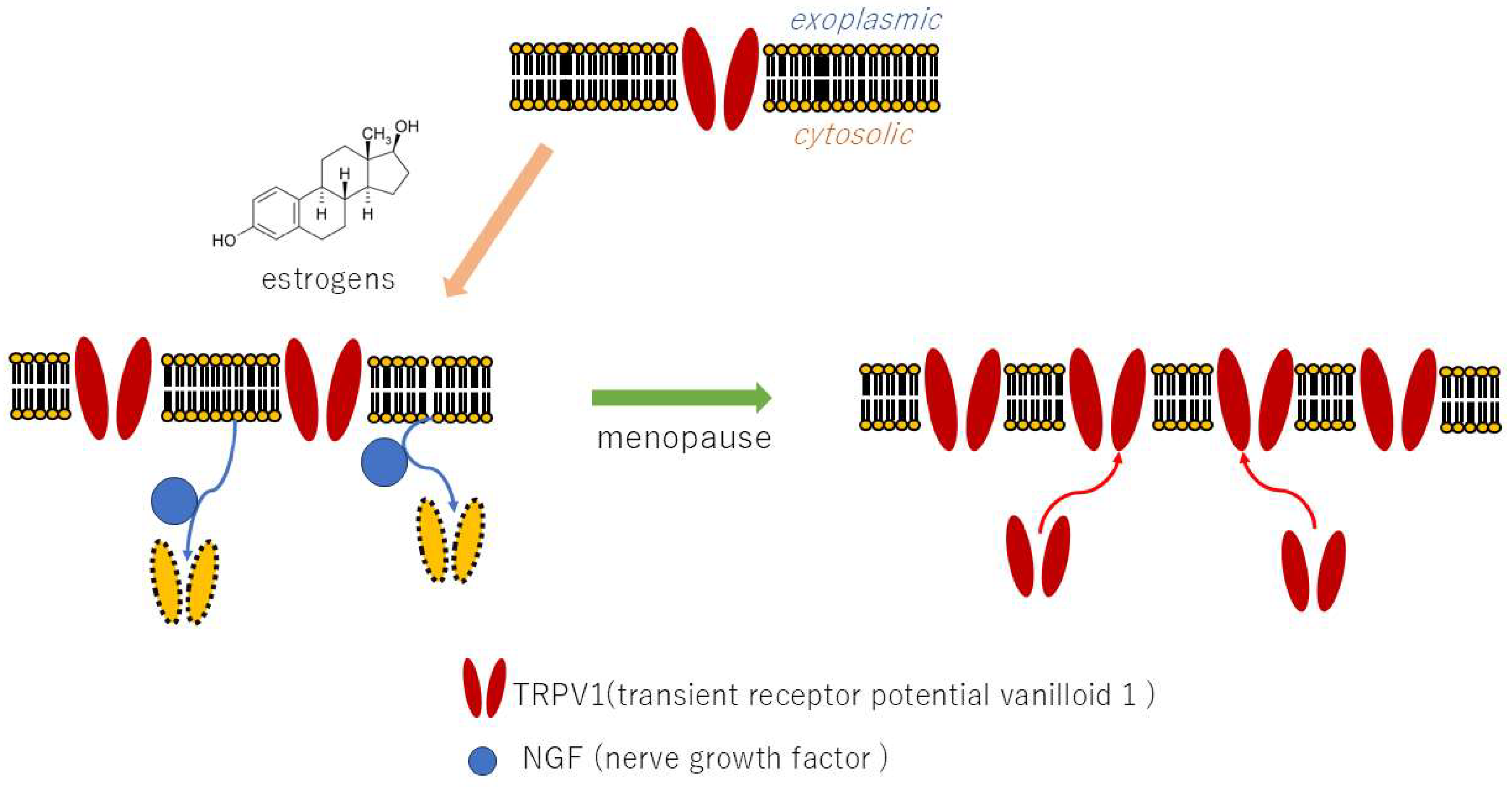
4.1. Nociceptors Expression
4.2. Wind-Up Phenomenon
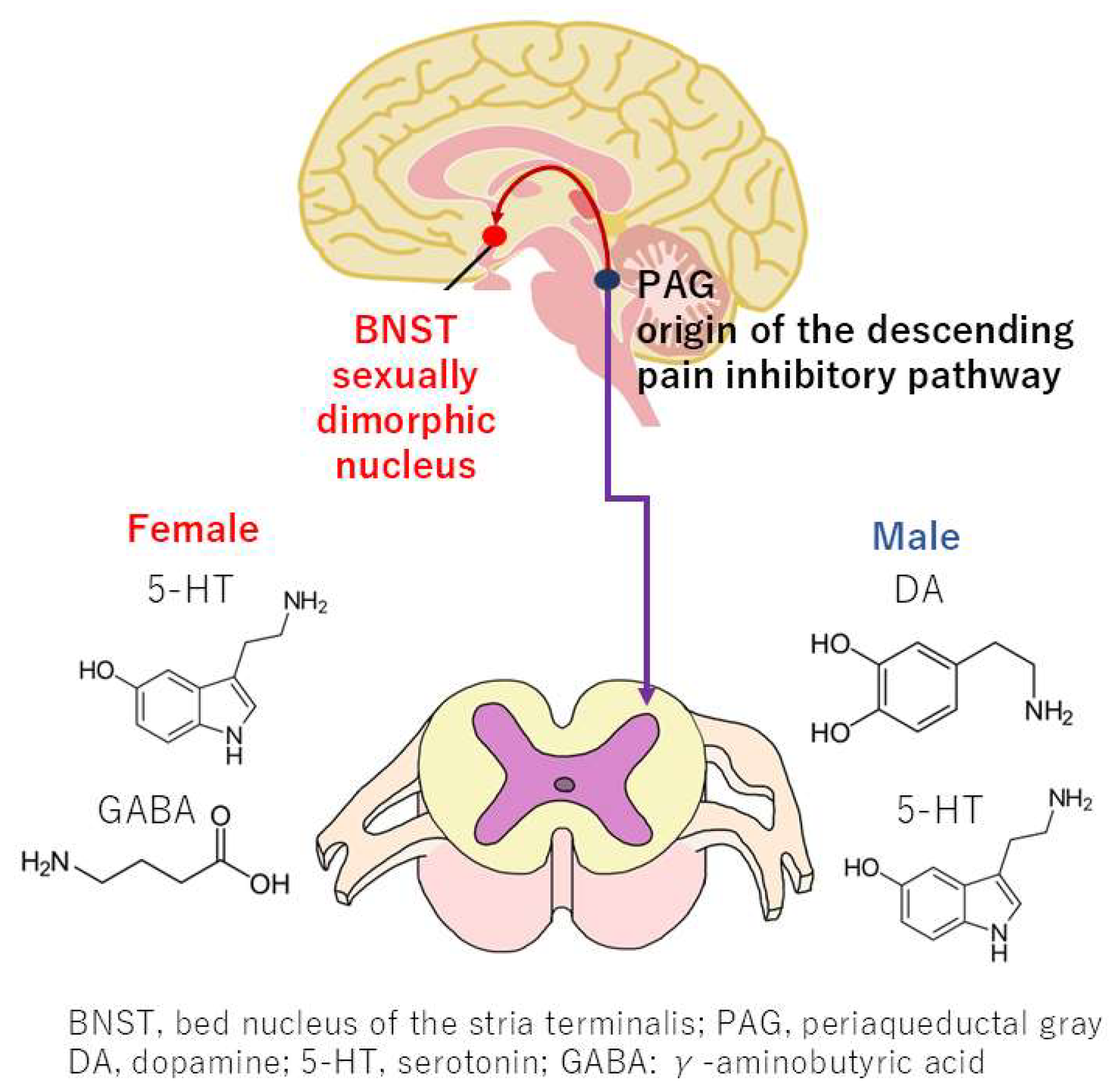
4.3. Sex Differences in the Descending Pain Inhibitory Pathway
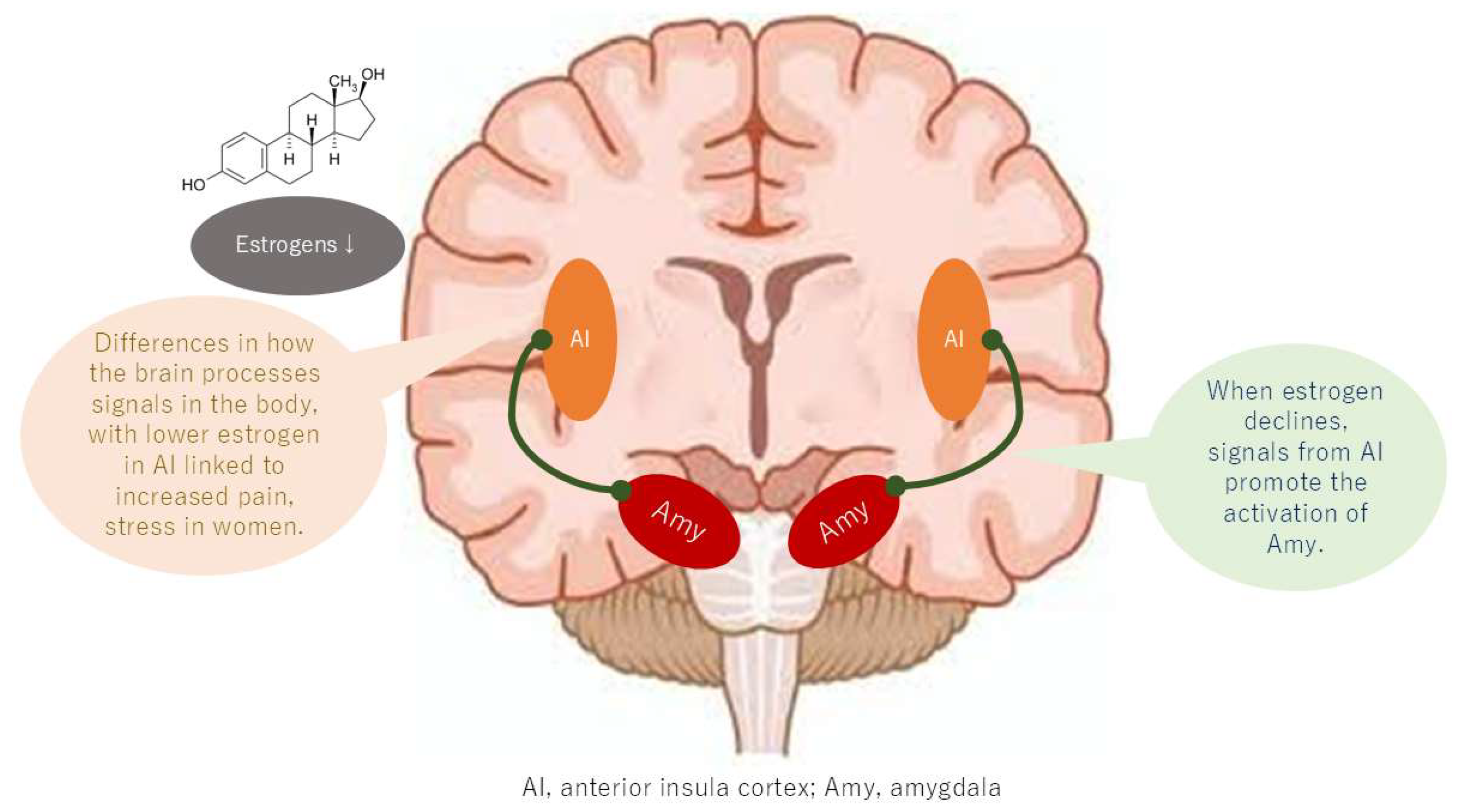
5. Sex Differences in the Brain Network Pain Circuit
5.1. Large-Scale Brain Networks
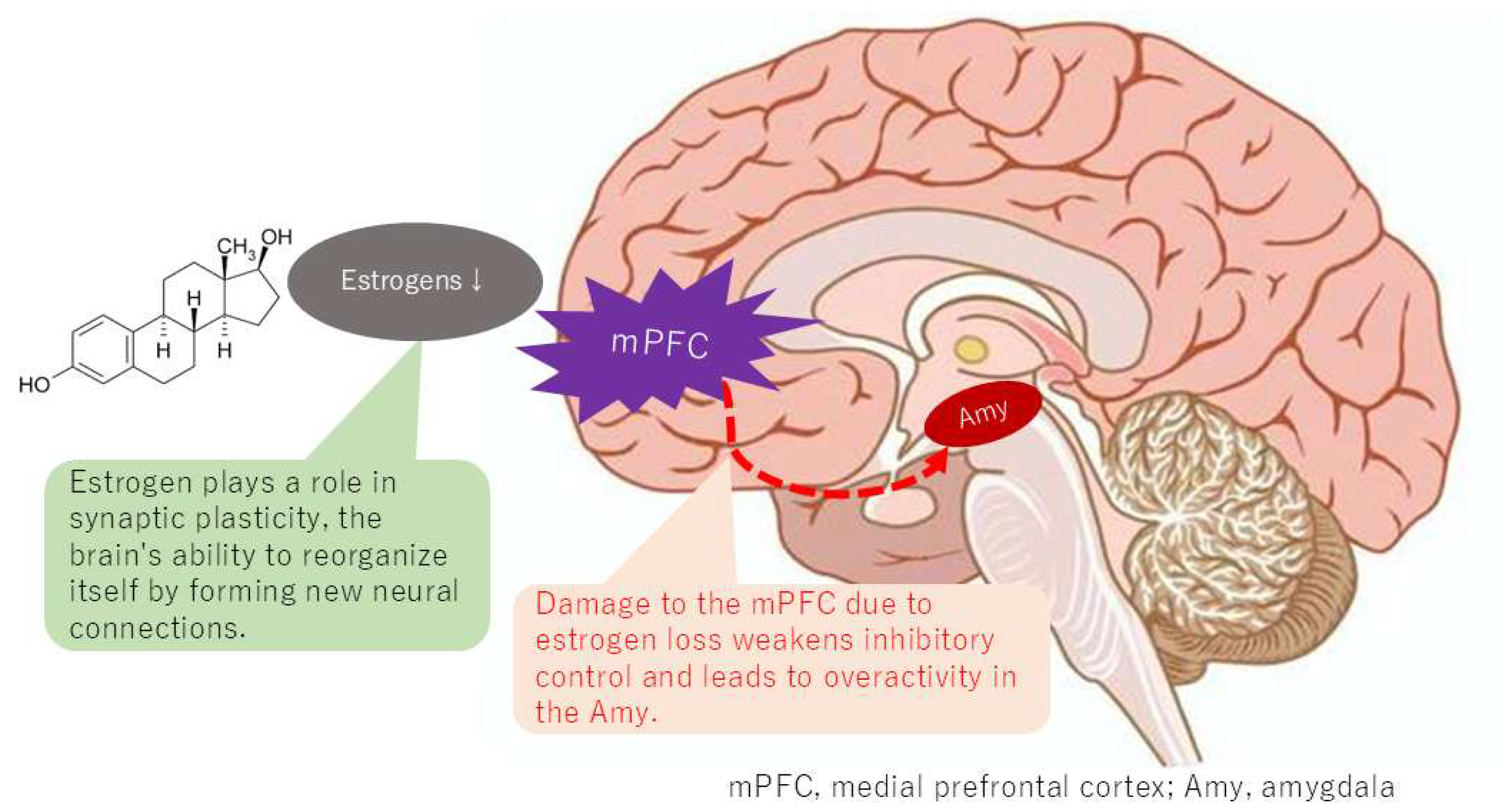
5.2. Changes in Functional Connectivity of the Default Mode Network
6. Sex Differences in the Memorized Pain Circuit
7. Levels of Evidence for Estrogen-Mediated Mechanisms
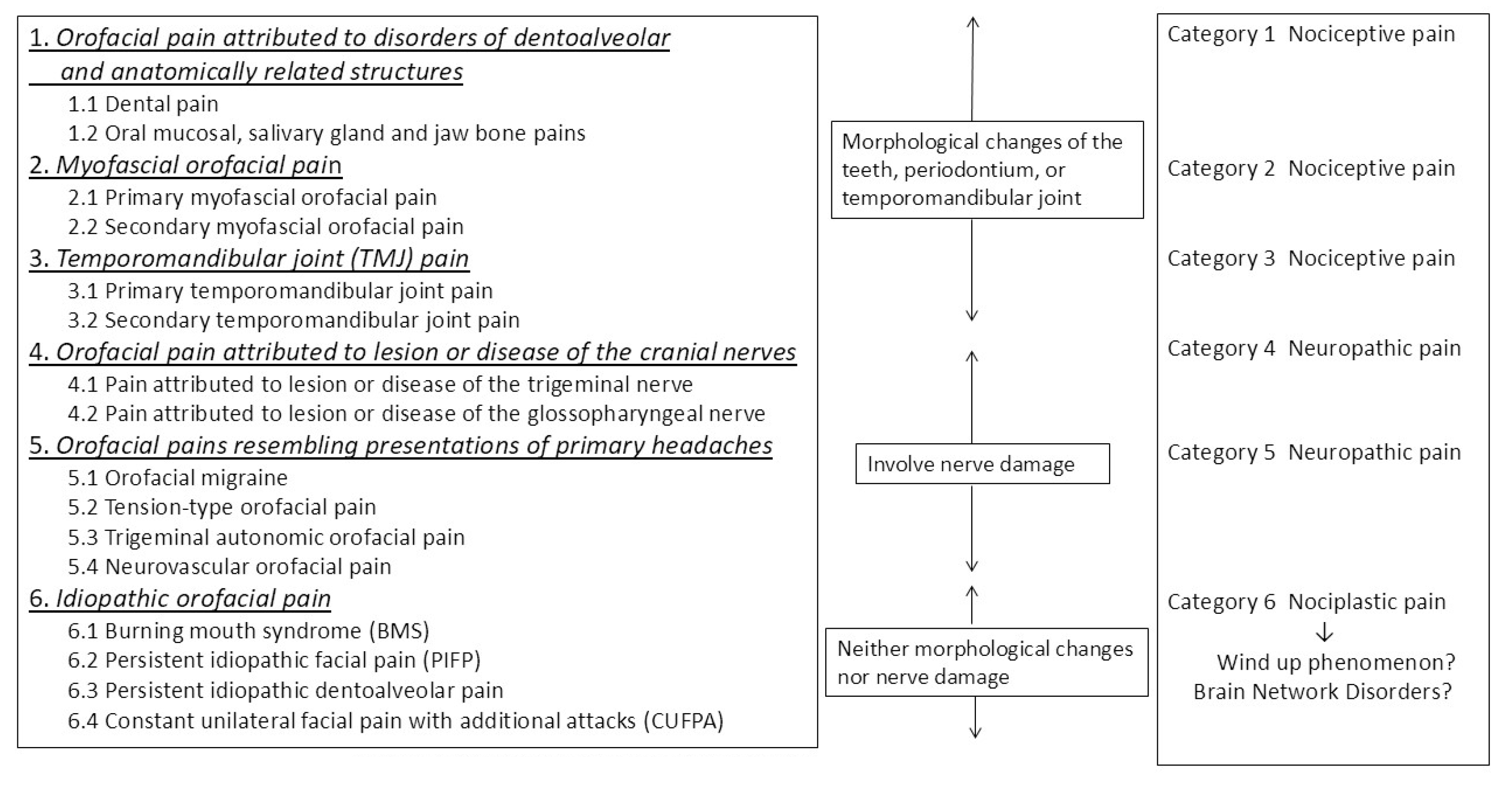
8. Sex Differences in ICOP Classification
9. Treatment
9.1. Treatment for the Peripheral Pain Circuit
9.2. Treatment for the Brain Network Pain Circuit and Memorized Pain Circuit
9.3. Hormone Replacement Therapy
10. Limitations and Future Challenges
11. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BMS | burning mouth syndrome |
| BNST | bed nucleus of stria terminalis |
| CBT | cognitive behavioral therapy |
| DMN | default mode network |
| fMRI | functional magnetic resonance imaging |
| fNIRS | functional near-infrared spectroscopy |
| HRT | hormone replacement therapy |
| IASP | International Association for the Study of Pain |
| ICOP | International Classification of Orofacial Pain |
| mPFC | medial prefrontal cortex |
| NGF | nerve growth factor |
| NMDA | N-methyl-D-aspartate |
| PAG | periaqueductal gray |
| PNS | peripheral nervous system |
| SN | salience network |
| TMS | transcranial magnetic stimulation |
| TRPV1 | transient receptor potential cation channel subfamily vanilloid member 1 |
References
- Benoliel, R.; May, A.; Svensson, P.; Pigg, M.; Law, A.; Nixdorf, D.; Renton, T.; Sharav, Y.; Ernberg, M.; Peck, C.; et al. International Classification of Orofacial Pain, 1st edition (ICOP). Cephalalgia 2020, 40, 129–221. [Google Scholar] [CrossRef]
- Kohorst, J.J.; Bruce, A.J.; Torgerson, R.R.; Schenck, L.A.; Davis, M.D.F. The prevalence of burning mouth syndrome: A population-based study. Br. J. Dermatol. 2015, 172, 1654–1656. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, W.; Yan, J.; Noma, N.; Young, A.; Yan, Z. Worldwide prevalence estimates of burning mouth syndrome: A systematic review and meta-analysis. Oral Dis. 2022, 28, 1431–1440. [Google Scholar] [CrossRef] [PubMed]
- de Lima-Souza, R.A.; Pérez-de-Oliveira, M.E.; Normando, A.G.C.; Louredo, B.V.R.; Mariano, F.V.; Farag, A.M.; Santos-Silva, A.R. Clinical and epidemiological profile of burning mouth syndrome patients following the International Headache Society classification: A systematic review and meta-analysis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2024, 137, 119–135. [Google Scholar] [CrossRef]
- Fitzcharles, M.-A.; Cohen, S.P.; Clauw, D.J.; Littlejohn, G.; Usui, C.; Häuser, W. Nociplastic pain: Towards an understanding of prevalent pain conditions. Lancet 2021, 397, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T. Burning mouth syndrome needs to consider the gut-brain axis from three types of pain: Nociceptive, neuropathic, and nociplastic pain. J. Gastrointest. Liver Dis. 2023, 32, 558–559. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T. Chronic orofacial pain. Br. Dent. J. 2024, 237, 153. [Google Scholar] [CrossRef]
- Kosek, E.; Clauw, D.; Nijs, J.; Baron, R.; Gilron, I.; Harris, R.E.; Mico, J.-A.; Rice, A.S.C.; Sterling, M. Chronic nociplastic pain affecting the musculoskeletal system: Clinical criteria and grading system. Pain 2021, 162, 2629–2634. [Google Scholar] [CrossRef]
- Herrero, J.F.; Laird, J.M.; Lopez-Garcia, J.A. Wind-up of spinal cord neurones and pain sensation: Much ado about something? Prog. Neurobiol. 2000, 61, 169–203. [Google Scholar] [CrossRef]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef]
- Nagamine, T. Pathogenesis of Orofacial Pain Based on Brain Circuits. Oral Dis. 2025. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T. Two-hit theory by estrogen in burning mouth syndrome. J. Dent. Sci. 2022, 17, 1833–1834. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Sugimura, M.; Yoshida, M.; Sekine, S.; Kawano, A.; Oyamaguchi, A.; Maegawa, H.; Niwa, H. Estrogens Exacerbate Nociceptive Pain via Up-Regulation of TRPV1 and ANO1 in Trigeminal Primary Neurons of Female Rats. Endocrinology 2016, 157, 4309–4317. [Google Scholar] [CrossRef] [PubMed]
- Stratton, H.; Lee, G.; Dolatyari, M.; Ghetti, A.; Cotta, T.; Mitchell, S.; Yue, X.; Ibrahim, M.; Dumaire, N.; Salih, L.; et al. Nociceptors are functionally male or female: From mouse to monkey to man. Brain 2024, 147, 4280–4291. [Google Scholar] [CrossRef]
- Seol, S.-H.; Chung, G. Estrogen-dependent regulation of transient receptor potential vanilloid 1 (TRPV1) and P2X purinoceptor 3 (P2X3): Implication in burning mouth syndrome. J. Dent. Sci. 2022, 17, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Lee, H.-J.; Lee, I.H.; Lee, J.-J. Risk of Burning Mouth Syndrome in Patients with Migraine: A Nationwide Cohort Study. J. Pers. Med. 2022, 12, 620. [Google Scholar] [CrossRef]
- Fileccia, E.; Incensi, A.; Ventruto, F.; Rizzo, G.; Galimberti, D.; Rao, G.; Salvi, F.; Liguori, R.; Donadio, V. Small Fiber Neuropathy in Patients with Chronic Pain and a Previous Diagnosis of Multiple Chemical Sensitivity Syndrome. J. Neuropathol. Exp. Neurol. 2021, 80, 868–874. [Google Scholar] [CrossRef]
- Taubøll, E.; Sveberg, L.; Svalheim, S. Interactions between hormones and epilepsy. Seizure 2015, 28, 3–11. [Google Scholar] [CrossRef]
- Ji, Y.; Bai, G.; Cao, D.; Traub, R.J. Estradiol modulates visceral hyperalgesia by increasing thoracolumbar spinal GluN2B subunit activity in female rats. Neurogastroenterol. Motil. 2015, 27, 775–786. [Google Scholar] [CrossRef]
- Traub, R.J.; Ji, Y. Sex differences and hormonal modulation of deep tissue pain. Front. Neuroendocr. 2013, 34, 350–366. [Google Scholar] [CrossRef]
- Shen, Z.; Li, W.; Chang, W.; Yue, N.; Yu, J. Sex differences in chronic pain-induced mental disorders: Mechanisms of cerebral circuitry. Front. Mol. Neurosci. 2023, 16, 1102808. [Google Scholar] [CrossRef] [PubMed]
- Horii, K.; Ehara, Y.; Shiina, T.; Naitou, K.; Nakamori, H.; Horii, Y.; Shimaoka, H.; Saito, S.; Shimizu, Y. Sexually dimorphic response of colorectal motility to noxious stimuli in the colorectum in rats. J. Physiol. 2021, 599, 1421–1437. [Google Scholar] [CrossRef]
- Horii, K.; Sawamura, T.; Onishi, A.; Yuki, N.; Naitou, K.; Shiina, T.; Shimizu, Y. Contribution of sex hormones to the sexually dimorphic response of colorectal motility to noxious stimuli in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 323, G1–G8. [Google Scholar] [CrossRef]
- Damascelli, M.; Woodward, T.S.; Sanford, N.; Zahid, H.B.; Lim, R.; Scott, A.; Kramer, J.K. Multiple Functional Brain Networks Related to Pain Perception Revealed by fMRI. Neuroinformatics 2022, 20, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.G.; Price, S.J.; Suckling, J. Functional connectivity networks for preoperative brain mapping in neurosurgery. J. Neurosurg. 2017, 126, 1941–1950. [Google Scholar] [CrossRef]
- Da Silva, J.T.; Tricou, C.; Zhang, Y.; Seminowicz, D.A.; Ro, J.Y. Brain networks and endogenous pain inhibition are modulated by age and sex in healthy rats. Pain 2020, 161, 1371–1380. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Keaser, M.L.; Meiller, T.F.; Seminowicz, D.A. Altered structure and function in the hippocampus and medial prefrontal cortex in patients with burning mouth syndrome. Pain 2014, 155, 1472–1480. [Google Scholar] [CrossRef]
- Ong, W.-Y.; Stohler, C.S.; Herr, D.R. Role of the Prefrontal Cortex in Pain Processing. Mol. Neurobiol. 2019, 56, 1137–1166. [Google Scholar] [CrossRef]
- Buckner, R.L. The brain’s default network: Origins and implications for the study of psychosis. Dialogues Clin. Neurosci. 2013, 15, 351–358. [Google Scholar] [CrossRef]
- Chung, Y.S.; Calhoun, V.; Stevens, M.C. Adolescent sex differences in cortico-subcortical functional connectivity during response inhibition. Cogn. Affect. Behav. Neurosci. 2020, 20, 1–18. [Google Scholar] [CrossRef]
- Shvetcov, A.; Ruitenberg, M.J.; Delerue, F.; Gold, W.A.; Brown, D.A.; Finney, C.A. The neuroprotective effects of estrogen and estrogenic compounds in spinal cord injury. Neurosci. Biobehav. Rev. 2023, 146, 105074. [Google Scholar] [CrossRef]
- Baudry, M.; Bi, X.; Aguirre, C. Progesterone–estrogen interactions in synaptic plasticity and neuroprotection. Neuroscience 2013, 239, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.; Hodgetts, S.; Hausmann, M. Sex differences and menstrual cycle effects in cognitive and sensory resting state networks. Brain Cogn. 2019, 131, 66–73. [Google Scholar] [CrossRef]
- Mueller, J.M.; Pritschet, L.; Santander, T.; Taylor, C.M.; Grafton, S.T.; Jacobs, E.G.; Carlson, J.M. Dynamic community detection reveals transient reorganization of functional brain networks across a female menstrual cycle. Netw. Neurosci. 2021, 5, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Chimenos-Küstner, E.; de Luca-Monasterios, F.; Schemel-Suárez, M.; de Rivera-Campillo, M.E.R.; Pérez-Pérez, A.M.; López-López, J. Burning mouth syndrome and associated factors: A case-control retrospective study. Med. Clin. 2017, 148, 153–157. [Google Scholar] [CrossRef]
- Koren, T.; Yifa, R.; Amer, M.; Krot, M.; Boshnak, N.; Ben-Shaanan, T.L.; Azulay-Debby, H.; Zalayat, I.; Avishai, E.; Hajjo, H.; et al. Insular cortex neurons encode and retrieve specific immune responses. Cell 2021, 184, 5902–5915.e17. [Google Scholar] [CrossRef] [PubMed]
- Wada, A.; Shizukuishi, T.; Kikuta, J.; Yamada, H.; Watanabe, Y.; Imamura, Y.; Shinozaki, T.; Dezawa, K.; Haradome, H.; Abe, O. Altered structural connectivity of pain-related brain network in burning mouth syndrome—Investigation by graph analysis of probabilistic tractography. Neuroradiology 2017, 59, 525–532. [Google Scholar] [CrossRef]
- Dugan, C.; Parlatescu, I.; Dobre, M.; Pîrvu, R.E.; Milanesi, E. Insights on brain functions in burning mouth syndrome. Front. Syst. Neurosci. 2022, 16, 975126. [Google Scholar] [CrossRef]
- McBenedict, B.; Petrus, D.; Pires, M.P.; Pogodina, A.; Agbor, D.B.A.; Ahmed, Y.A.; Ceron, J.I.C.; Balaji, A.; Abrahão, A.; Pessôa, B.L.; et al. The Role of the Insula in Chronic Pain and Associated Structural Changes: An Integrative Review. Cureus 2024, 16, e58511. [Google Scholar] [CrossRef]
- Öz, F.; Acer, N.; Katayifçi, N.; Aytaç, G.; Karaali, K.; Sindel, M. The role of lateralisation and sex on insular cortex: 3D volumetric analysis. Turk. J. Med. Sci. 2021, 51, 1240–1248. [Google Scholar] [CrossRef]
- Jørgensen, M.R.; Pedersen, A.M. Analgesic effect of topical oral capsaicin gel in burning mouth syndrome. Acta Odontol. Scand. 2017, 75, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Kuten-Shorrer, M.; Treister, N.; Stock, S.; Kelley, J.; Ji, Y.; Woo, S.-B.; Lerman, M.A.; Palmason, S.; Sonis, S.T.; Villa, A. Topical Clonazepam Solution for the Management of Burning Mouth Syndrome: A Retrospective Study. J. Oral Facial Pain Headache 2017, 31, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Gramacy, A.; Villa, A. Topical gabapentin solution for the management of burning mouth syndrome: A retrospective study. PLoS ONE 2023, 18, e0295559. [Google Scholar] [CrossRef]
- Cui, Y.; Xu, H.; Chen, F.M.; Liu, J.L.; Jiang, L.; Zhou, Y.; Chen, Q.M. Efficacy evaluation of clonazepam for symptom remission in burning mouth syndrome: A meta-analysis. Oral Dis. 2016, 22, 503–511. [Google Scholar] [CrossRef]
- Noma, N.; Watanabe, Y.; Shimada, A.; Usuda, S.; Iida, T.; Shimada, A.; Tanaka, Y.; Oono, Y.; Sasaki, K. Effects of cognitive behavioral therapy on orofacial pain conditions. J. Oral Sci. 2020, 63, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, C.; de Klerk, C.C.J.M.; Richards, J.E.; Southgate, V.; Hamilton, A.; Blasi, A. The developmental trajectory of fronto-temporoparietal connectivity as a proxy of the default mode network: A longitudinal fNIRS investigation. Hum. Brain Mapp. 2020, 41, 2717–2740. [Google Scholar] [CrossRef]
- Cheung, D.; Trudgill, N. Managing a patient with burning mouth syndrome. Front. Gastroenterol. 2015, 6, 218–222. [Google Scholar] [CrossRef]
- Brauwers, K.G.; Bueno, V.M.; Calcia, T.B.B.; Daroit, N.B. Sexual hormones changes in burning mouth syndrome: A systematic review. J. Oral Rehabil. 2024, 51, 2475–2483. [Google Scholar] [CrossRef]
- Nagamine, T. Molecular Mechanisms of Pain Regulation by Sex Hormones in Individuals With Burning Mouth Syndrome. J. Oral Rehabil. 2025. [Google Scholar] [CrossRef]
- Athnaiel, O.; Cantillo, S.; Paredes, S.; Knezevic, N.N. The Role of Sex Hormones in Pain-Related Conditions. Int. J. Mol. Sci. 2023, 24, 1866. [Google Scholar] [CrossRef]
- Cao, B.; Xu, Q.; Shi, Y.; Zhao, R.; Li, H.; Zheng, J.; Liu, F.; Wan, Y.; Wei, B. Pathology of pain and its implications for therapeutic interventions. Signal Transduct. Target. Ther. 2024, 9, 155. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author (Year) | Pain Circuit | Estrogen-Mediated Mechanisms | Study Phase |
|---|---|---|---|
| Seol et al., 2022 [15] | Peripheral pain | Regulation of expression and intracellular trafficking of transient receptor potential cation channel subfamily V member 1 (TRPV1) | Preclinical |
| Taubøll et al., 2015 [18] | Peripheral pain | Changes in the sensitivity of N-Methyl-D-aspartic acid (NMDA) receptors and are involved in the wind-up phenomenon | Preclinical |
| Shen et al., 2023 [21] | Peripheral pain | Affects the structure of the bed nucleus of the stria terminalis (BNST) and alters emotional responses | Preclinical |
| Horii et al., 2023 [23] | Peripheral pain | Regulation of neurotransmitter expression in the descending pain inhibitory pathway | Preclinical |
| Ong et al., 2019 [28] | Brain network pain | Hypofunction of the medial prefrontal cortex (mPFC) and altered connectivity of the default mode network | Clinical |
| McBenedict et al., 2024 [39] | Memorized pain | Involved in activation of the anterior insular cortex and anterior cingulate cortex | Clinical |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagamine, T. Estrogen-Mediated Neural Mechanisms of Sex Differences in Burning Mouth Syndrome. Neurol. Int. 2025, 17, 61. https://doi.org/10.3390/neurolint17040061
Nagamine T. Estrogen-Mediated Neural Mechanisms of Sex Differences in Burning Mouth Syndrome. Neurology International. 2025; 17(4):61. https://doi.org/10.3390/neurolint17040061
Chicago/Turabian StyleNagamine, Takahiko. 2025. "Estrogen-Mediated Neural Mechanisms of Sex Differences in Burning Mouth Syndrome" Neurology International 17, no. 4: 61. https://doi.org/10.3390/neurolint17040061
APA StyleNagamine, T. (2025). Estrogen-Mediated Neural Mechanisms of Sex Differences in Burning Mouth Syndrome. Neurology International, 17(4), 61. https://doi.org/10.3390/neurolint17040061