
Gut Microbiota and Its Repercussion in Parkinson’s Disease: A Systematic Review in Occidental Patients

 , ,
, ,
Abstract
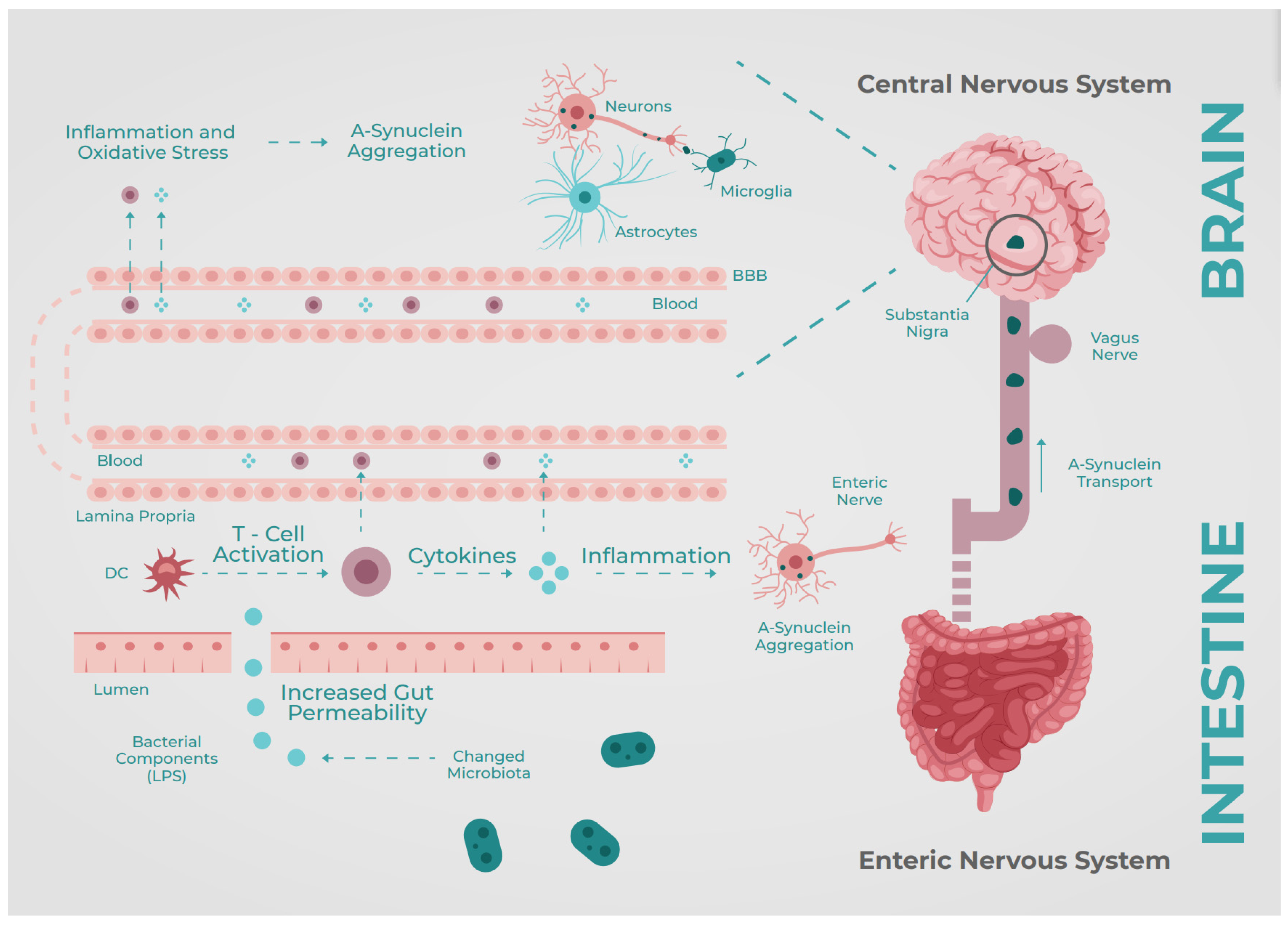
1. Introduction

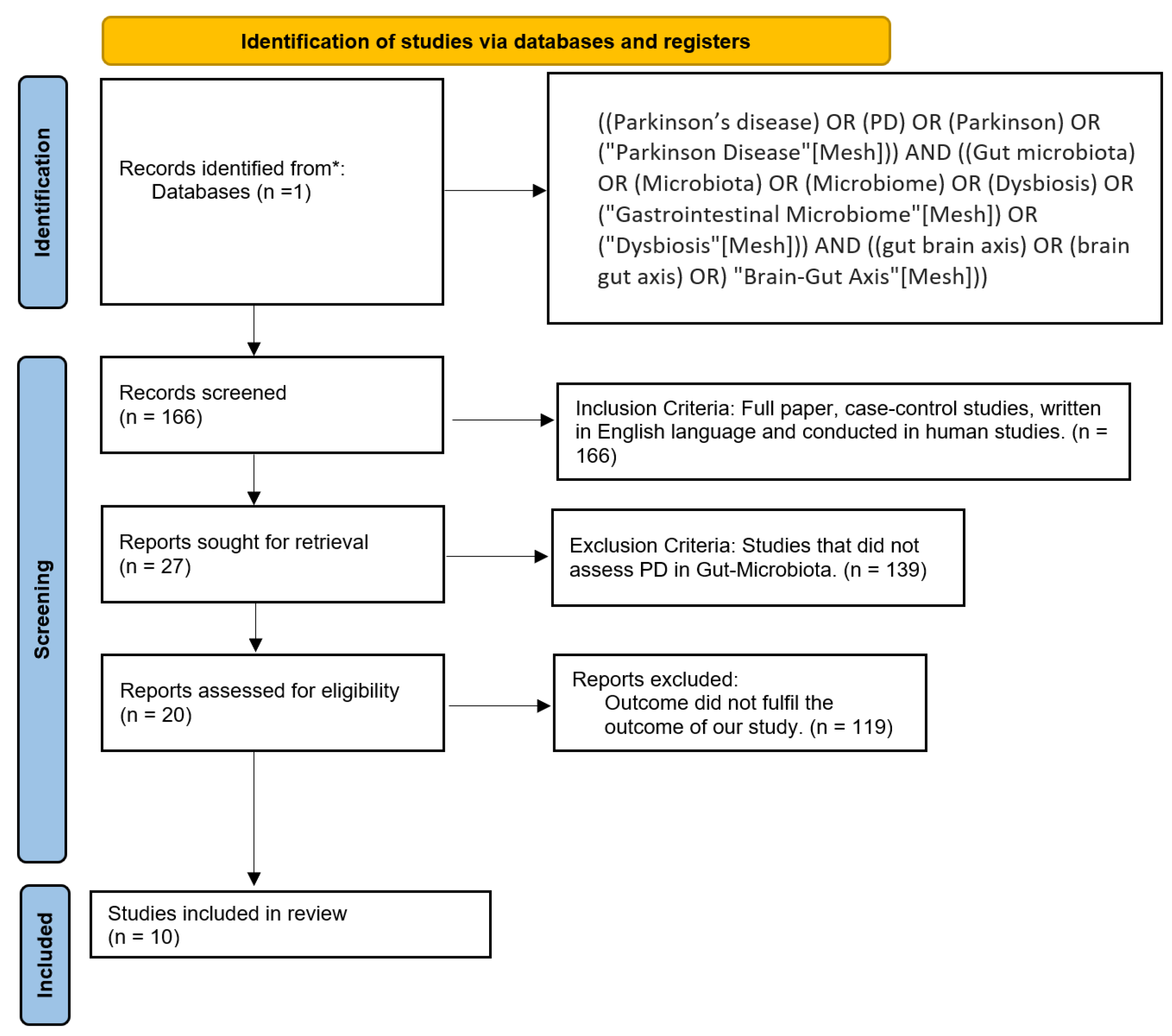
2. Materials and Methods
2.1. Protocol
2.2. Eligibility Criteria and Study Selection
2.3. Database and Search Strategy
2.4. Data Extraction and Analysis
2.5. Bias Analysis
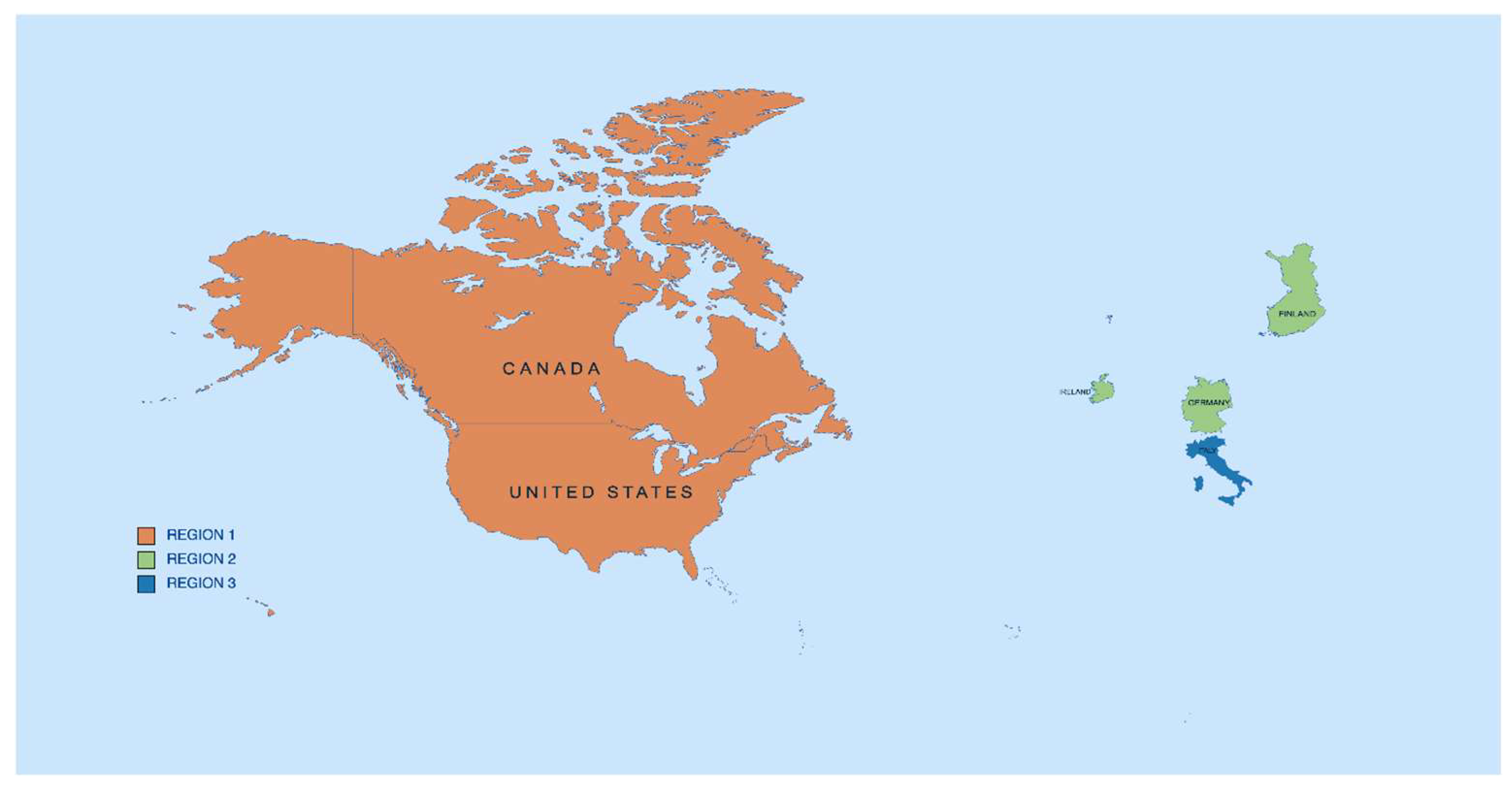
3. Results
3.1. Region 1
3.2. Region 2
3.3. Region 3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BBB | Blood–brain barrier |
| CNS | Central nervous system |
| COMTi | Catechol-O-methyltransferase inhibitors |
| GBA | Gut–Brain Axis |
| HC | Healthy controls |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| IND | Indeterminate |
| LPS | Lipopolysaccharides |
| MOOSE | Meta-analysis of observational studies in epidemiology |
| PD | Parkinson’s disease |
| PIGD | Postural Instability and Gait dysfunction |
| PRISMA | Preferred Reporting Items for Systematic Reviews and Meta-Analyses |
| SCFAs | Short-chain fatty acids |
References
- Adlerberth, I. Factors Influencing the Establishment of the Intestinal Microbiota in Infancy. In Nestlé Nutrition Workshop Series: Pediatric Program; Bier, D.M., German, J.B., Lönnerdal, B., Eds.; KARGER: Basel, Switzerland, 2008; Volume 62, pp. 13–33. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The Healthy Human Microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. The Gut Microbiota—Masters of Host Development and Physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Julian, R.Y.; Hardesty, D.A.; Lonser, R.R. Cerebral Edema. In Youmans & Winn Neurological Surgery; Winn, H.R., Ed.; Elsevier: Philadelphia, PA, USA, 2023; pp. 473.e1–473.e13. [Google Scholar]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-Analysis of Gut Microbiome Studies Identifies Disease-Specific and Shared Responses. Nat. Commun. 2017, 8, 1784. [Google Scholar] [CrossRef]
- Blacher, E.; Bashiardes, S.; Shapiro, H.; Rothschild, D.; Mor, U.; Dori-Bachash, M.; Kleimeyer, C.; Moresi, C.; Harnik, Y.; Zur, M.; et al. Potential Roles of Gut Microbiome and Metabolites in Modulating ALS in Mice. Nature 2019, 572, 474–480. [Google Scholar] [CrossRef]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The Role of Microbiota-Gut-Brain Axis in Neuropsychiatric and Neurological Disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Fasano, A.; Visanji, N.P.; Liu, L.W.C.; Lang, A.E.; Pfeiffer, R.F. Gastrointestinal Dysfunction in Parkinson’s Disease. Lancet Neurol. 2015, 14, 625–639. [Google Scholar] [CrossRef]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Oleskin, A.V.; Shenderov, B.A. Neuromodulatory Effects and Targets of the SCFAs and Gasotransmitters Produced by the Human Symbiotic Microbiota. Microb. Ecol. Health Dis. 2016, 27, 30971. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Duncan, S.H.; McCrae, S.I.; Millar, J.; Jackson, M.S.; Flint, H.J. Restricted Distribution of the Butyrate Kinase Pathway among Butyrate-Producing Bacteria from the Human Colon. J. Bacteriol. 2004, 186, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Van IJzendoorn, S.C.D.; Derkinderen, P. The Intestinal Barrier in Parkinson’s Disease: Current State of Knowledge. J. Park. Dis. 2019, 9, S323–S329. [Google Scholar] [CrossRef]
- Fayyad, M.; Salim, S.; Majbour, N.; Erskine, D.; Stoops, E.; Mollenhauer, B.; El-Agnaf, O.M.A. Parkinson’s Disease Biomarkers Based on A-synuclein. J. Neurochem. 2019, 150, 626–636. [Google Scholar] [CrossRef]
- Li, J.; Meng, P.; Zhang, J.; He, M. Effect of Berberine Hydrochloride on the Diversity of Intestinal Flora in Parkinson’s Disease Patients. Contrast Media Mol. Imaging 2022, 2022, 8381870. [Google Scholar] [CrossRef]
- MapChart. Available online: https://www.mapchart.net/ (accessed on 17 May 2023).
- Keshavarzian, A.; Green, S.J.; Engen, P.A.; Voigt, R.M.; Naqib, A.; Forsyth, C.B.; Mutlu, E.; Shannon, K.M. Colonic Bacterial Composition in Parkinson’s Disease: Colonic Microbiota in Parkinson’s Disease. Mov. Disord. 2015, 30, 1351–1360. [Google Scholar] [CrossRef]
- Hill-Burns, E.M.; Debelius, J.W.; Morton, J.T.; Wissemann, W.T.; Lewis, M.R.; Wallen, Z.D.; Peddada, S.D.; Factor, S.A.; Molho, E.; Zabetian, C.P.; et al. Parkinson’s Disease and Parkinson’s Disease Medications Have Distinct Signatures of the Gut Microbiome: PD, Medications, and Gut Microbiome. Mov. Disord. 2017, 32, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Cirstea, M.S.; Sundvick, K.; Golz, E.; Yu, A.C.; Boutin, R.C.T.; Kliger, D.; Finlay, B.B.; Appel-Cresswell, S. The Gut Mycobiome in Parkinson’s Disease. J. Park. Dis. 2021, 11, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Paul, K.C.; Jacobs, J.P.; Chou, H.-C. (Lori); Duarte Folle, A.; Del Rosario, I.; Yu, Y.; Bronstein, J.M.; Keener, A.M.; Ritz, B. Parkinson’s Disease and the Gut Microbiome in Rural California. J. Park. Dis. 2022, 12, 2441–2452. [Google Scholar] [CrossRef]
- Pietrucci, D.; Cerroni, R.; Unida, V.; Farcomeni, A.; Pierantozzi, M.; Mercuri, N.B.; Biocca, S.; Stefani, A.; Desideri, A. Dysbiosis of Gut Microbiota in a Selected Population of Parkinson’s Patients. Park. Relat. Disord. 2019, 65, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Barichella, M.; Severgnini, M.; Cilia, R.; Cassani, E.; Bolliri, C.; Caronni, S.; Ferri, V.; Cancello, R.; Ceccarani, C.; Faierman, S.; et al. Unraveling Gut Microbiota in Parkinson’s Disease and Atypical Parkinsonism. Mov. Disord. 2019, 34, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, F.; Künstner, A.; Müller, S.H.; Künzel, S.; Zeuner, K.E.; Margraf, N.G.; Deuschl, G.; Baines, J.F.; Kuhlenbäumer, G. Gut Microbiota in Parkinson Disease in a Northern German Cohort. Brain Res. 2017, 1667, 41–45. [Google Scholar] [CrossRef]
- Heintz-Buschart, A.; Pandey, U.; Wicke, T.; Sixel-Döring, F.; Janzen, A.; Sittig-Wiegand, E.; Trenkwalder, C.; Oertel, W.H.; Mollenhauer, B.; Wilmes, P. The Nasal and Gut Microbiome in Parkinson’s Disease and Idiopathic Rapid Eye Movement Sleep Behavior Disorder: Nose and Gut Microbiome in PD and IRBD. Mov. Disord. 2018, 33, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Hertel, J.; Harms, A.C.; Heinken, A.; Baldini, F.; Thinnes, C.C.; Glaab, E.; Vasco, D.A.; Pietzner, M.; Stewart, I.D.; Wareham, N.J.; et al. Integrated Analyses of Microbiome and Longitudinal Metabolome Data Reveal Microbial-Host Interactions on Sulfur Metabolism in Parkinson’s Disease. Cell Rep. 2019, 29, 1767–1777.e8. [Google Scholar] [CrossRef] [PubMed]
- Aho, V.T.E.; Pereira, P.A.B.; Voutilainen, S.; Paulin, L.; Pekkonen, E.; Auvinen, P.; Scheperjans, F. Gut Microbiota in Parkinson’s Disease: Temporal Stability and Relations to Disease Progression. EBioMedicine 2019, 44, 691–707. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Study | Newcastle–Ottawa Scale | Overall Risk of Bias | ||
|---|---|---|---|---|
| Selection (Total 4) | Comparability (Total 2) | Outcome/Exposure (Total 3) | (Total score) | |
| Keshavarzian et al., 2015, USA [17]. | *** | ** | * | Moderate (6) |
| Hill-Burns et al., 2017, USA [18]. | ** | ** | ** | Moderate (6) |
| Appel-Cresswell et al., 2020, Canada [19]. | - | * | * | High (2) |
| Zhang et al., 2020, USA [20]. | **** | ** | *** | Low (9) |
| Pietrucci et al., 2019, Italy [21]. | *** | * | *** | Moderate (6) |
| Cereda et al., 2018, Italy [22]. | *** | * | ** | Moderate (6) |
| Hopfner et al., 2017, Germany [23]. | **** | * | ** | Low (7) |
| Heintz-Buschart et al., 2017, Germany [24]. | *** | ** | ** | Low (7) |
| Hertel, J. et al., 2019, Ireland [25]. | *** | ** | ** | Low (7) |
| Aho et al., 2019, Finland [26]. | *** | * | *** | Moderate (6) |
| Author/Country/Year * | Sample Number | Methodology | Microbiota Alteration in PD | Mechanisms (Theories) | Association | Limitations |
|---|---|---|---|---|---|---|
| Keshavarzian et al. [17], 2015, USA | Cases: (n = 38) Controls: (n = 34) | Fecal and colonic mucosa samples: Microbial 16S rRNA gene on genomic DNA. | - Less abundant in fecal samples: Family: Lachnospiraceae Genus: Blautia, Coprococcus, Roseburia - Less abundant in mucosa samples: Family: Coprobacillaceae Genus: Dorea, Faecalibacterium - More abundant in fecal samples: Phylum: Bacteroidetes, Proteobacteria, Verrucomicrobiota Genus: Akkermansia, Oscillospira, Bacteroides - More abundant in mucosa samples: Family: Oxalobacteraceae Genus: Ralstonia. | Family Lachnospiraceae contains several butyrate-producing bacteria, butyrate has anti-inflammatory properties. Thus, the low abundance of butyrate-producing bacteria in feces from PD subjects may be one mechanism contributing to intestinal leakiness and inflammation in PD. | Yes | Environmental factors associated with PD (age, BMI, medication use, and PD duration) might explain differences between cases and control. The study found significant differences in age between PD and controls, but its impact is uncertain. |
| Hill-Burns et al. [18], 2017, USA | Cases: (n = 197) Controls PD-household: (n = 130) | Fecal samples: 16S rRNA amplicon sequencing on genomic DNA. | -Less abundant in fecal samples: Family: Lachnospiraceae - More abundant in fecal samples: Genus: Akkermansia, Lactobacillus, Bifidobacterium. | SCFA are made by bacteria in the gut; notably, Lachnospiraceae is consistent with its depletion. Butyrate kinase, which catalyzes a reversible reaction between butyrate and butanoylphosphate, was reduced in PD, and acetyl-CoA synthetase, which converts acetate to acetyl-CoA, was elevated. | Yes | Most relevant potential confounders: PD medications, disease duration, marital status, and geographic site. |
| Appel-Cresswell et al. [19], 2020, Canada | Cases: (n = 95) Controls: (n = 57) | Fecal samples: Microbial DNA was extracted using QIAGEN and amplicon base sequenced. | -Mycobiome found in fecal samples: Phylum: 86% Ascomycota,12% Basidiomycota Genus: Saccharomyces (most common 58.7%) in 94% of participants, Candida 35%, Cladosporium 23%, Penicillium 23%. | Found no correlation between PD patients and control fecal samples. | No | - Only one ITS primer pair was used for sequencing. - Additional data on environmental factors were not collected. |
| Zhang et al. [20], 2022, USA | Cases: (n = 96) Controls PD-household: (n = 74) | Fecal samples: 16S rRNA gene sequencing, rarefaction, and feature filtering. | - More abundant in fecal samples: Phylum: Proteobacteria, Verrucomicrobiota, and Actinobacteriota. Genus: UBA1819 (Ruminococcaceae), DTU089 (Ruminococcaceae), Akkermansia, Enterococcus, and Hungatella. | Increased level of Proteobacteria has been associated with potential immunoregulation ability via the production of LPS. They initiate the immune process, stimulating microglia and leading to dopaminergic neuron necrosis. | Yes | - Gastrointestinal comorbidities. - The recent use of antibiotics was not specified. - Size, confounders control, and statistical power. - The samples were collected at a single time; this does not allow for establishing temporality or causality. - The species-level resolution of the sequencing and annotation pipeline. |
| # | Genus | Family | Order | Class | Phylum | |
|---|---|---|---|---|---|---|
| Actinobacteriota | ↑ | ✔ | ||||
| Akkermansia | ↑ | ✔ | Akkermansiaceae | Verrucomicrobiales | Verrucomicrobiae | Verrucomicrobiota |
| Bacteroides | ↑ | ✔ | Bacteroidaceae | Bacteroidales | Bacteroidia | Bacteroidetes |
| Bacteroidetes | ↑ | ✔ | ||||
| Bifidobacterium | ↑ | ✔ | Bifidobacteriaceae | Bifidobacteriales | Actinomycetia | Actinobacteriota |
| Blautia | ↓ | ✔ | Lachnospiraceae | Eubacteriales | Clostridia | Firmicutes |
| Coprococcus | ↓ | ✔ | Lachnospiraceae | Eubacteriales | Clostridia | Firmicutes |
| Enterococcus | ↑ | ✔ | Enterococcaceae | Lactobacillales | Bacilli | Firmicutes |
| Hungatella | ↑ | ✔ | Clostridiaceae | Clostridiales | Clostridia | Firmicutes |
| Lachnospiraceae | ↓ | ✔ | Eubacteriales | Clostridia | Firmicutes | |
| Lactobacillus | ↑ | ✔ | Lactobacillaceae | Lactobacillales | Bacilli | Firmicutes |
| Oscillospira | ↑ | ✔ | Oscillospiraceae | Eubacteriales | Clostridia | Firmicutes |
| Proteobacteria | ↑ | ✔ | ||||
| Roseburia | ↓ | ✔ | Lachnospiraceae | Eubacteriales | Clostridia | Firmicutes |
| Ruminococcaceae | ↑ | ✔ | Clostridiales | Clostridia | Firmicutes | |
| Verrucomicrobiota | ↑ | ✔ |
| Author/Country/Year * | Sample Number | Methodology | Microbiota Alteration in PD | Mechanisms (Theories) | Association | Limitations |
|---|---|---|---|---|---|---|
| Hopfner et al., 2017, Germany [23]. | Cases: (n = 29) Controls: (n = 29) | Samples: Next-generation sequencing of the 16S rRNA gene. | - Significant increase in PD patients of Family: Lactobacillaceae - Higher abundance in PD cases of: Family: Barnesiellaceae. | The GI microbiome influences the enteric nervous system and through the vagal nerve reaches the central nervous system. | Yes | - Previous dietary habits, also cardiovascular comorbidities were present in the control group. - Cases were using PD treatment. |
| Heintz-buschart et al., 2017, Germany [24]. ** | Cases: (n = 99) Controls: (n = 76) | Samples: 16S and 18S ribosomal RNA amplicon sequencing from flash-frozen stool and nasal swab. | - Relative abundances in the PD patients of: Genus: Akkermansia Family: Verrucomicrobiaceae - In addition, 75% showed the same changes in the gut microbiome between the PD patients and RBD vs. the HC: Genus: Anaerotruncus, Clostridium XIVb. Phylum: Bacteroidetes. | Akkermansia spp. abundance is related to higher susceptibility to a pathogen due to the depletion of the mucus layer. | Yes | - Comorbidity of diabetes and coronary artery disease were found to affect the prokaryotic taxonomic profiles. - Oral intake of diabetes medication was considered a potential confounder. |
| Hertel et al., 2019, Ireland [25]. | Cases: (n = 30) Controls: (n = 30) | - Targeted metabolomic analysis. - Computational analysis methods and the Virtual Metabolic Human database. | - Significant increase in abundance of: Genus: Akkermansia muciniphila and Bilophila wadsworthia. | A. muciniphila produces hydrogen sulfide, which is pro-inflammatory and harmful to the gut. This correlates to gastrointestinal motility dysfunction and higher absorption of bacterial toxins through the gut barrier in PD patients. B. Wadsworthia produces sulfite, which is pro-inflammatory to the gut and a neurotoxin. | Yes | - The effects of exercise were not monitored. - The study results could be influenced by dietary variance. - The study results cannot describe the mechanics involved in PD presentation and metabolic alterations. |
| Aho et al., 2019, Finland [26]. | Cases: (n = 64) Controls: (n = 64) | Stool samples: twice on average 2–2.5 years apart: 16s rRNA gene amplicon sequencing. | - Increased in fecal samples in PD patients at baseline and follow-up: Genus: Bifidobacterium, Lactobacillus, Roseburia—progressed PD patients had a Firmicutes-dominated enterotype. - Less abundant in PD patients and faster progression patients: Genus: Prevotella. | The early nonmotor symptoms of PD led to the hypothesis that it could originate outside the CNS, in the enteric nervous system. | Yes | - Deficient follow-up. - PD intake medication stroke or TIA was common among the controls. |
| # | Spp | Genus | Family | Order | Class | Phylum | |
|---|---|---|---|---|---|---|---|
| Akkermansia | ↑ | ✔ | Akkermansiaceae | Verrucomicrobiales | Verrucomicrobiae | Verrucomicrobiota | |
| Akkermansia Muciniphila | ↑ | ✔ | Akkermansia | Akkermansiaceae | Verrucomicrobiales | Verrucomicrobiae | Verrucomicrobiota |
| Barnesiellaceae | ↑ | ✔ | Bacteroidales | Bacteroidia | Bacteroidetes | ||
| Bifidobacterium | ↑ | ✔ | Bifidobacteriaceae | Bifidobacteriales | Actinomycetia | Actinobacteriota | |
| Bilophila wadsworthia | ↑ | ✔ | Bilophila | Desulfovibrionaceae | Desulfovibrionales | Desulfovibrionia | Thermodesulfobacteriota |
| Lactobacillaceae | ↑ | ✔ | Lactobacillales | Bacilli | Firmicutes | ||
| Lactobacillus | ↑ | ✔ | Lactobacillaceae | Lactobacillales | Bacilli | Firmicutes | |
| Prevotella | ↓ | ✔ | Prevotellaceae | Bacteroidales | Bacteroidia | Bacteroidetes | |
| Roseburia | ↑ | ✔ | Lachnospiraceae | Eubacteriales | Clostridia | Firmicutes | |
| Verrucomicrobiaceae | ↑ | ✔ | Verrucomicrobiales | Verrucomicrobiae | Verrucomicrobiota | ||
| Verrucomicrobiota | ↑ | ✔ |
| Author/Country/Year * | Sample Number | Methodology | Microbiota Alteration in PD | Mechanisms (Theories) | Association | Limitations |
|---|---|---|---|---|---|---|
| Pietrucci et al., 2019, Italy [21]. | Cases: (n = 80) Controls: (n = 72) | Fecal samples: 16S rRNA amplicon (V3–V4 regions) sequencing analysis PD-COMTi takers: 8. | - Higher levels in the feces compared to controls: Phylum: Proteobacteria, Firmicutes Family: Enterobacteriaceae, Lactobacillaceae, Enterococcaceae - Lower levels in the feces compared to controls: Phylum: Firmicutes Family: Lachnospiraceae - More abundant in PD- COMTi: Genus: Citrobacter, Enterococcus, Lactococcus, Klebsiella, Salmonella, Shigella, and an unclassified Enterobacteriaceae - Less abundant in PD- COMTi: Genus: Roseburia. | Gut microorganisms metabolize phenylalanine, tryptophan, and tyrosine. This study found they were reduced in PD patients. An increase in the presence of Gram-negative bacteria leads to the elevation of LPS content. Thus, increasing intestinal permeability. | Yes | - Unbalanced cases and controls. - Age and sex. - Loss of 5 kg in the last year. |
| Cereda et al., 2018, Italy [22]. | Cases: (n = 193) Controls: (n = 113) | Fecal samples: 16S gene ribosomal RNA sequencing. | - Higher levels in fecal samples of PD compared to controls: Phylum: Verrucomicrobiota, Proteobacteria Family: Enterobacteriaceae, Lactobacillaceae, Coriobacteriaceae, Bifidobacteriaceae Genus: Akkermansia, Christensenellaceae, ParaBacteroides, Oscillospira - Lower levels in fecal samples of PD compared to controls: Family: Lachnospiraceae Genus: Roseburia, Ruminococcus. | Production of SCFA such as butyrate, propionate, and acetate. Additionally, Lactobacillaceae is involved in the activity of the enteric nervous system and its secretion of α-synuclein and brain chemistry through the vagal nerve. | Yes | - Lack of sigmoid mucosal biopsies. - Lack of characterization of microbial functions through metagenomic analyses. - Presence of irritable bowel syndrome. |
| # | Genus | Family | Order | Class | Phylum | |
|---|---|---|---|---|---|---|
| Akkermansia | ↑ | ✔ | Akkermansiaceae | Verrucomicrobiales | Verrucomicrobiae | Verrucomicrobiota |
| Bifidobacteriaceae | ↑ | ✔ | Bifidobacteriales | Actinomycetia | Actinobacteriota | |
| Christensenellaceae | ↑ | ✔ | Eubacteriales | Clostridia | Firmicutes | |
| Citrobacter | ↑ | ✔ | Enterobacteriaceae | Enterobacterales | Gammaproteobacteria | Proteobacteria |
| Coriobacteriaceae | ↑ | ✔ | Coriobacteriales | Coriobacteriia | Actinobacteriota | |
| Enterobacteriaceae | ↑ | ✔ | Enterobacterales | Gammaproteobacteria | Proteobacteria | |
| Enterococcaceae | ↑ | ✔ | Lactobacillales | Bacilli | Firmicutes | |
| Enterococcus | ↑ | ✔ | Enterococcaceae | Lactobacillales | Bacilli | Firmicutes |
| Firmicutes * | ↑↓ | ✔ | ||||
| Klebsiella | ↑ | ✔ | Enterobacteriaceae | Enterobacterales | Gammaproteobacteria | Proteobacteria |
| Lachnospiraceae | ↓ | ✔ | Eubacteriales | Clostridia | Firmicutes | |
| Lactobacillaceae | ↑ | ✔ | Lactobacillales | Bacilli | Firmicutes | |
| Lactococcus | ↑ | ✔ | Streptococcaceae | Lactobacillales | Bacilli | Firmicutes |
| Oscillospira | ↑ | ✔ | Oscillospiraceae | Eubacteriales | Clostridia | Firmicutes |
| ParaBacteroides | ↑ | ✔ | Tannerellaceae | Bacteroidales | Bacteroidia | Bacteroidetes |
| Proteobacteria | ↑ | ✔ | ||||
| Roseburia | ↓ | ✔ | Lachnospiraceae | Eubacteriales | Clostridia | Firmicutes |
| Ruminococcus | ↓ | ✔ | Oscillospiraceae | Eubacteriales | Clostridia | Firmicutes |
| Salmonella | ↑ | ✔ | Enterobacteriaceae | Enterobacterales | Gammaproteobacteria | Proteobacteria |
| Shigella | ↑ | ✔ | Enterobacteriaceae | Enterobacterales | Gammaproteobacteria | Proteobacteria |
| Verrucomicrobiota | ↑ | ✔ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proano, A.C.; Viteri, J.A.; Orozco, E.N.; Calle, M.A.; Costa, S.C.; Reyes, D.V.; German-Montenegro, M.; Moncayo, D.F.; Tobar, A.C.; Moncayo, J.A. Gut Microbiota and Its Repercussion in Parkinson’s Disease: A Systematic Review in Occidental Patients. Neurol. Int. 2023, 15, 750-763. https://doi.org/10.3390/neurolint15020047
Proano AC, Viteri JA, Orozco EN, Calle MA, Costa SC, Reyes DV, German-Montenegro M, Moncayo DF, Tobar AC, Moncayo JA. Gut Microbiota and Its Repercussion in Parkinson’s Disease: A Systematic Review in Occidental Patients. Neurology International. 2023; 15(2):750-763. https://doi.org/10.3390/neurolint15020047
Chicago/Turabian StyleProano, Ana Cristina, Javier A. Viteri, Eileen N. Orozco, Marco Antonio Calle, Stefany Carolina Costa, Daniel V. Reyes, Melissa German-Montenegro, David F. Moncayo, Andrea C. Tobar, and Juan A. Moncayo. 2023. "Gut Microbiota and Its Repercussion in Parkinson’s Disease: A Systematic Review in Occidental Patients" Neurology International 15, no. 2: 750-763. https://doi.org/10.3390/neurolint15020047
APA StyleProano, A. C., Viteri, J. A., Orozco, E. N., Calle, M. A., Costa, S. C., Reyes, D. V., German-Montenegro, M., Moncayo, D. F., Tobar, A. C., & Moncayo, J. A. (2023). Gut Microbiota and Its Repercussion in Parkinson’s Disease: A Systematic Review in Occidental Patients. Neurology International, 15(2), 750-763. https://doi.org/10.3390/neurolint15020047