

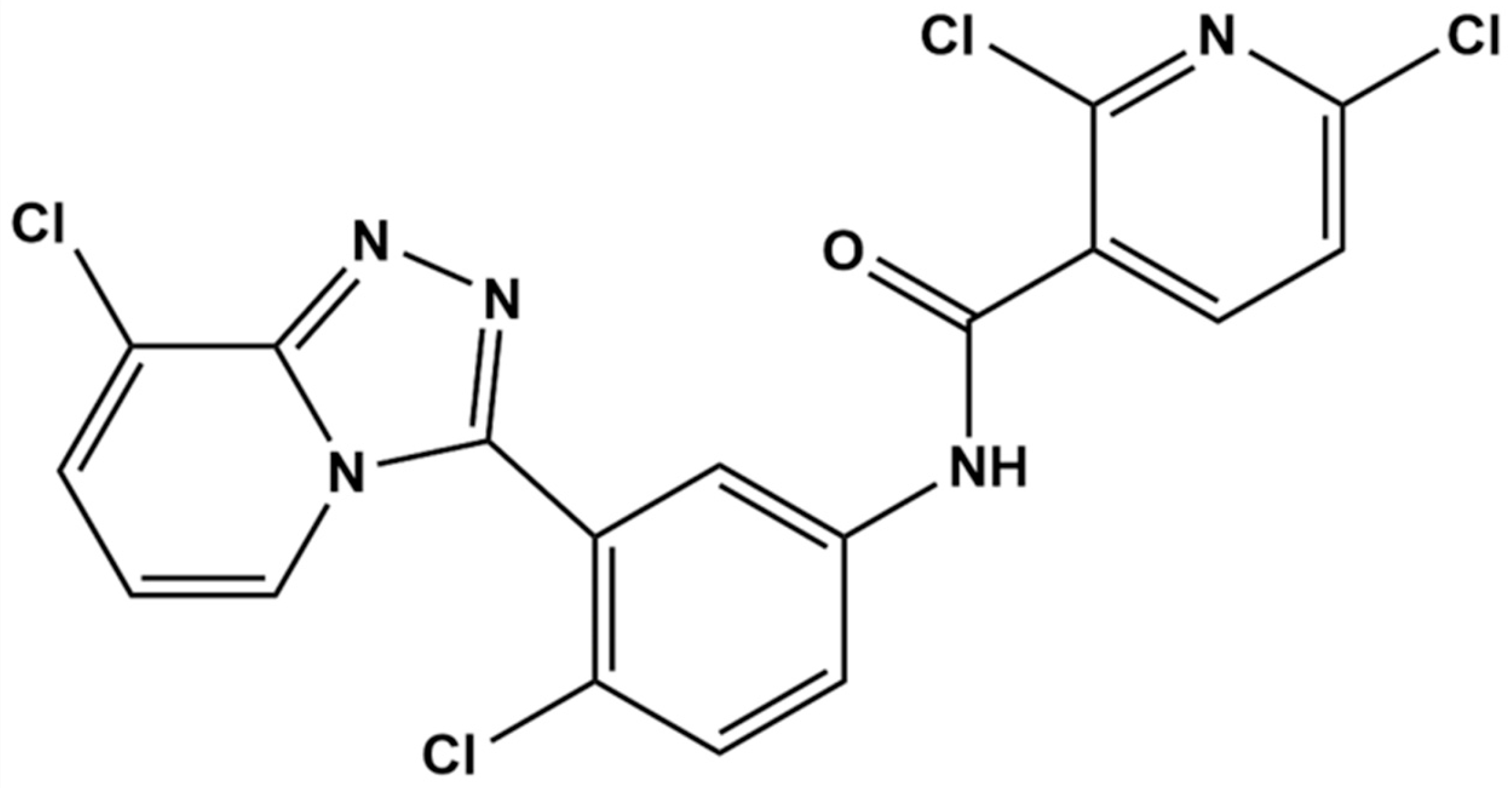
Plasma Protein Binding, Biostability, Metabolite Profiling, and CYP450 Phenotype of TPB15 Across Different Species: A Novel Smoothened Inhibitor for TNBC Therapy

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Methods
2.2.1. The Plasma Protein Binding Rates of TPB15 in Plasma from Mice, Rat, Dog, Monkey, and Human
2.2.2. Stability of TPB15 in Plasma from Mice, Rat, Dog, Monkey, and Human
2.2.3. Metabolic Stability of TPB15 in Liver Microsome from Mice, Rat, Dog, Monkey, and Human
2.2.4. Metabolic Enzyme Phenotypes in Human Liver Microsomes
2.2.5. Metabolite Identification
2.2.6. Statistical Analysis
3. Results
3.1. The Plasma Protein Binding Rates of TPB15 in Plasma from Mice, Rat, Dog, Monkey, and Human
3.1.1. The Non-Specific Adsorption Coefficient of TPB15 to the Ultrafiltration Tubes
3.1.2. The Plasma Protein Binding Rates
3.2. Stability of TPB15 in Plasma from Mice, Rat, Dog, Monkey, and Human
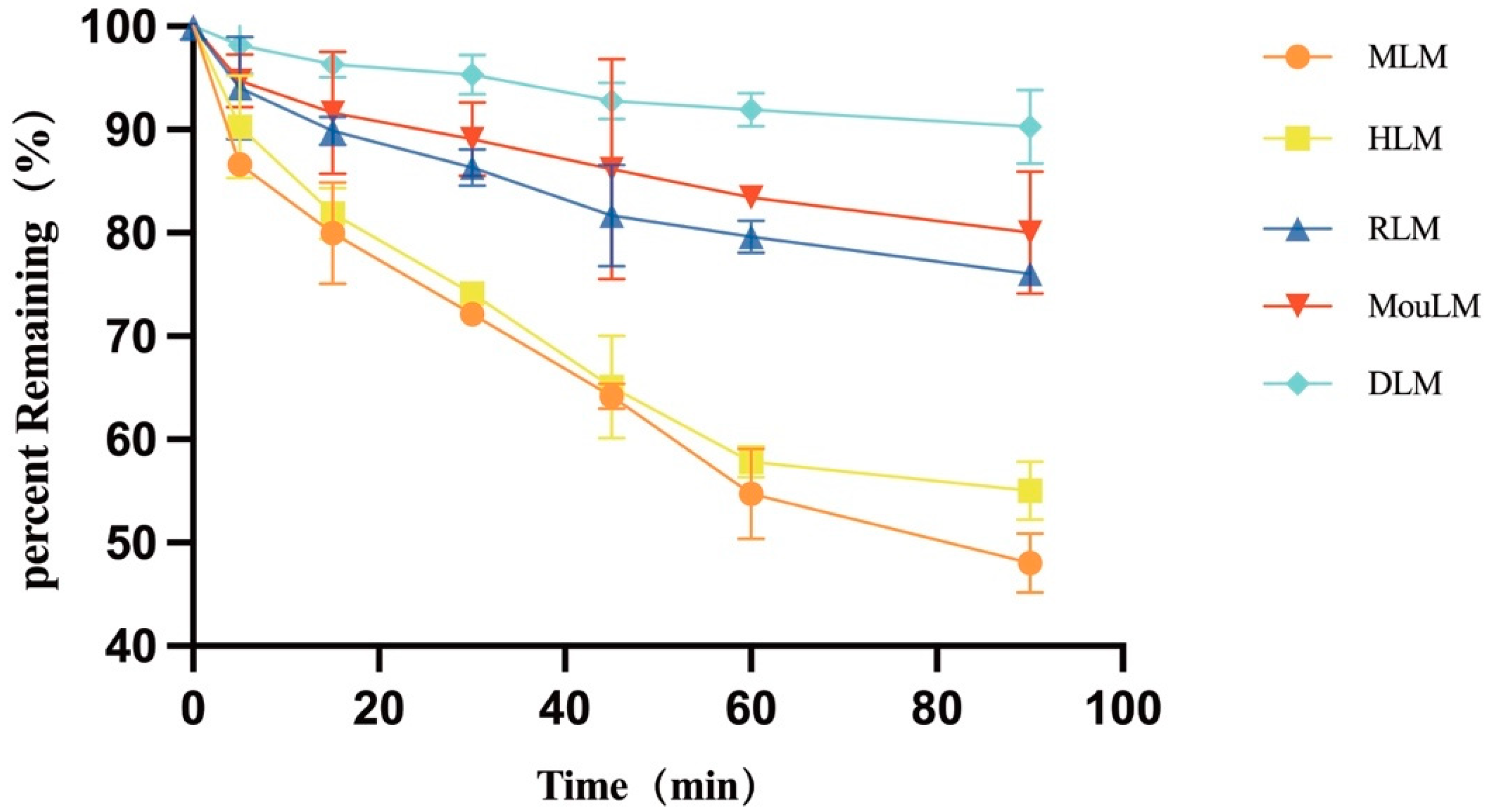
3.3. Metabolic Stability of TPB15 in Liver Microsome from Mice, Rat, Dog, Monkey, and Human
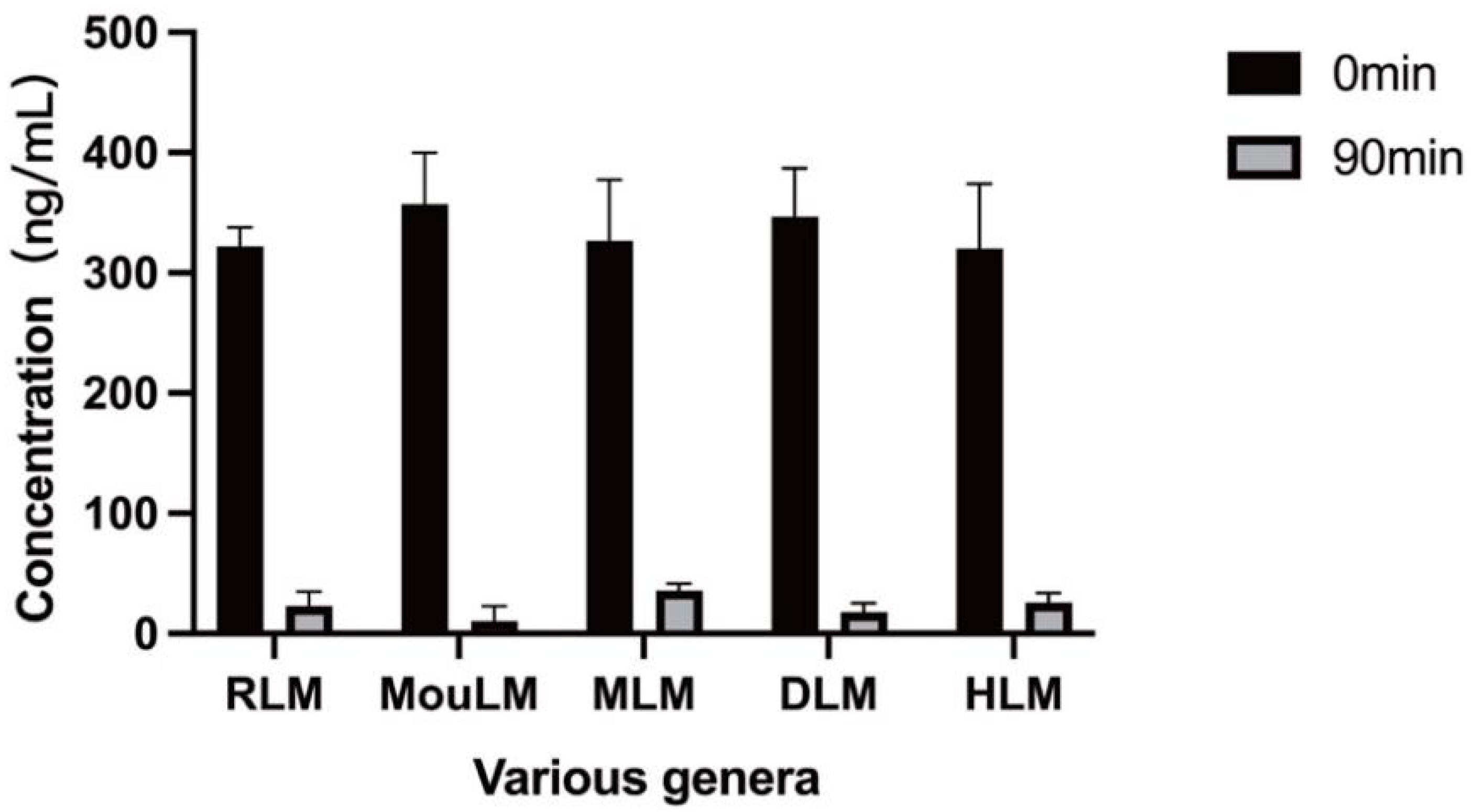
3.3.1. Positive Drug Control Groups in Liver Microsomes from Various Species
3.3.2. Metabolic Stability of TPB15 in Liver Microsome from Mice, Rat, Dog, Monkey, and Human
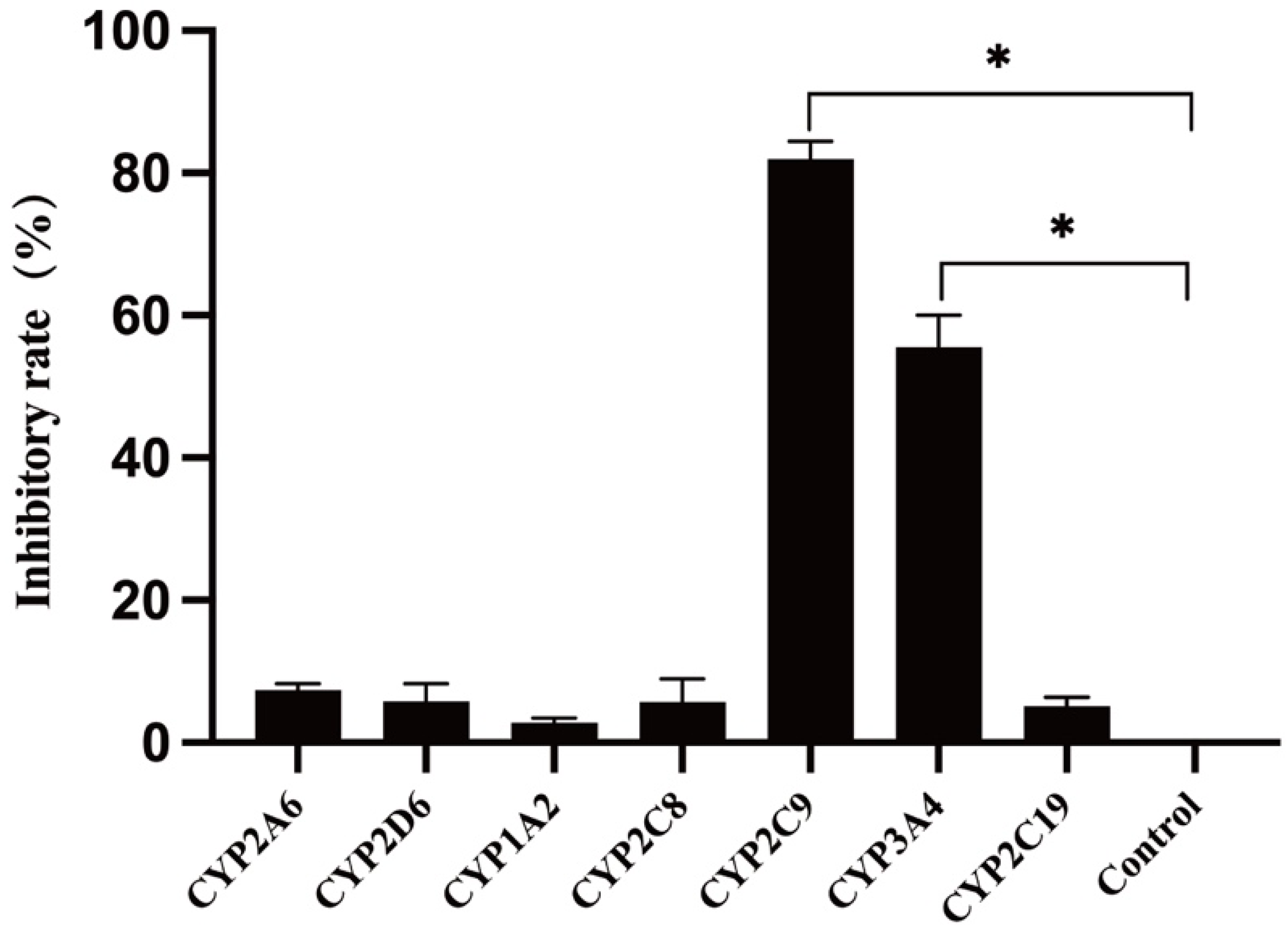
3.4. Metabolic Enzyme Phenotypes in Human Liver Microsomes
3.5. Metabolite Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TNBC | Triple-negative breast cancer |
| Hh | Hedgehog |
| CYP450 | Cytochrome P450 |
| Gli | Glioma-associated oncogene |
| GPCRs | G-protein coupled receptors |
| IC50 | Half-maximal inhibitory concentration |
| LD50 | Lethal dose for 50% of the population |
References
- Krstic, M.N.; Mijac, D.D.; Popovic, D.D.; Markovic, A.P.; Milosavljevic, T. General Aspects of Primary Cancer Prevention. Dig. Dis. 2019, 37, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Ramin, C.; Gierach, G.L.; Abubakar, M.; Veiga, L.H.S.; Vo, J.B.; Curtis, R.E.; Bowles, E.J.; Feigelson, H.S.; Buist, D.S.M.; de Gonzalez, A.B.; et al. The influence of treatment on hormone receptor subgroups and breast cancer-specific mortality within US integrated healthcare systems. Cancer Causes Control 2022, 33, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Butti, R.; Gunasekaran, V.P.; Kumar, T.V.S.; Banerjee, P.; Kundu, G.C. Breast cancer stem cells: Biology and therapeutic implications. Int. J. Biochem. Cell Biol. 2019, 107, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Skoda, A.M.; Simovic, D.; Karin, V.; Kardum, V.; Vranic, S.; Serman, L. The role of the Hedgehog signaling pathway in cancer: A comprehensive review. Bosn. J. Basic Med. Sci. 2018, 18, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Varjosalo, M.; Taipale, J. Hedgehog: Functions and mechanisms. Genes. Dev. 2008, 22, 2454–2472. [Google Scholar] [CrossRef] [PubMed]
- Niyaz, M.; Khan, M.S.; Mudassar, S. Hedgehog Signaling: An Achilles’ Heel in Cancer. Transl. Oncol. 2019, 12, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Du, J.; Zhao, L.; Liu, W.; Zhao, T.M.; Liang, H.; Fang, P.; Zhang, K.X.; Zeng, H. GLI1 reduces drug sensitivity by regulating cell cycle through PI3K/AKT/GSK3/CDK pathway in acute myeloid leukemia. Cell Death Dis. 2021, 12, 231. [Google Scholar] [CrossRef] [PubMed]
- Terao, T.; Minami, Y. Targeting Hedgehog (Hh) Pathway for the Acute Myeloid Leukemia Treatment. Cells 2019, 8, 312. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Z.L.; Jia, J.H. Mechanisms of Smoothened Regulation in Hedgehog Signaling. Cells 2021, 10, 2138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.W.; Fu, X.J.; Ni, L.; Liu, C.C.; Zheng, Y.X.; You, H.J.; Li, M.; Xiu, C.M.; Zhang, L.; Gong, T.T.; et al. Hedgehog Signaling Controls Bone Homeostasis by Regulating Osteogenic/Adipogenic Fate of Skeletal Stem/Progenitor Cells in Mice. J. Bone Miner. Res. 2022, 37, 559–576. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.N.; Zhao, H.X.; Wang, Y.Z.; Yu, C.D.; Liu, J.J.; Li, L.; Li, Z.H.; Zhang, J.; Dai, H.; Wang, J.F.; et al. Solubilization, purification, and ligand binding characterization of G protein-coupled receptor SMO in native membrane bilayer using styrene maleic acid copolymer. PeerJ 2022, 10, e13381. [Google Scholar] [CrossRef] [PubMed]
- Tian, N.N.; Wu, H.X.; Zhang, H.W.; Yang, D.N.; Lv, L.; Yang, Z.C.; Zhang, T.T.; Quan, D.L.; Zhou, L.; Xie, Y.; et al. Discovery of 1,2,4 triazolo 4,3-a pyridines as potent Smoothened inhibitors targeting the Hedgehog pathway with improved antitumor activity in vivo. Bioorg. Med. Chem. 2020, 28, 115584. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.Y.; Xu, J.H.; Chen, Q.Q.; Wu, H.X.; Ou, B.F.; Zhou, Z.; Xu, F.; Wu, S.Y.; Xie, S.L.; Wen, D.S. Pharmacokinetics and Bioavailability Study of a Novel Smoothened Inhibitor TPB15 for Treatment of Triple-Negative Breast Cancer in Rats by High Performance Liquid Chromatography-Mass Spectrometry. Eur. J. Drug Metab. Pharmacokinet. 2024, 49, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Marcella, M.; Geny, M.M.G.; de Kanter, R. Species differences between mouse, rat, dog, monkey and human CYP-mediated drug metabolism. Inhib. Induction 2006, 2, 875–894. [Google Scholar] [CrossRef]
- Li, D. An update on the importance of plasma protein binding in drug discovery and development. Expert Opin. Drug Discov. 2021, 16, 1453–1465. [Google Scholar] [CrossRef]
- Doohan, P.T.; Oldfield, L.D.; Arnold, J.C.; Anderson, L.L. Cannabinoid Interactions with Cytochrome P450 Drug Metabolism: A Full-Spectrum Characterization. AAPS J. 2021, 23, 91. [Google Scholar] [CrossRef] [PubMed]
- Dang, N.L.; Matlock, M.K.; Hughes, T.B.; Swamidass, S.J. The Metabolic Rainbow: Deep Learning Phase I Metabolism in Five Colors. J. Chem. Inf. Model. 2020, 60, 1146–1164. [Google Scholar] [CrossRef] [PubMed]

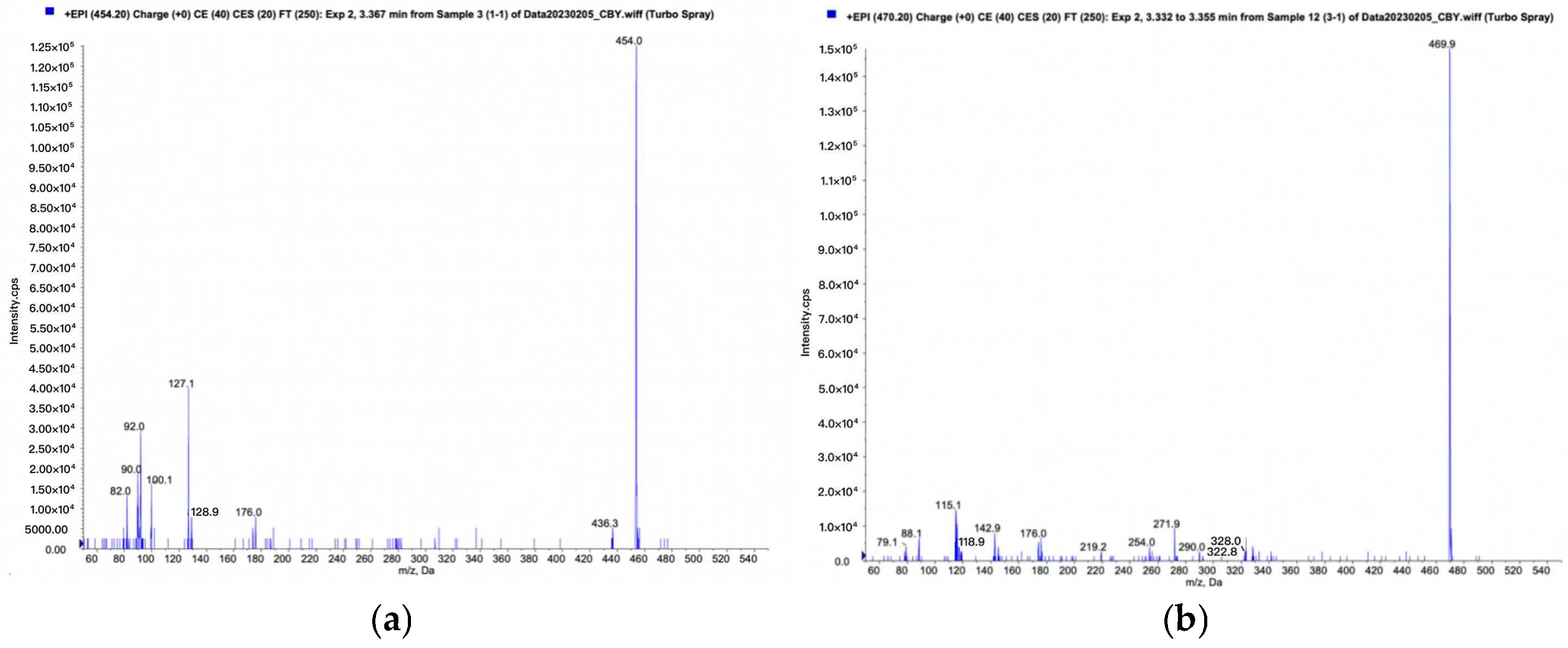
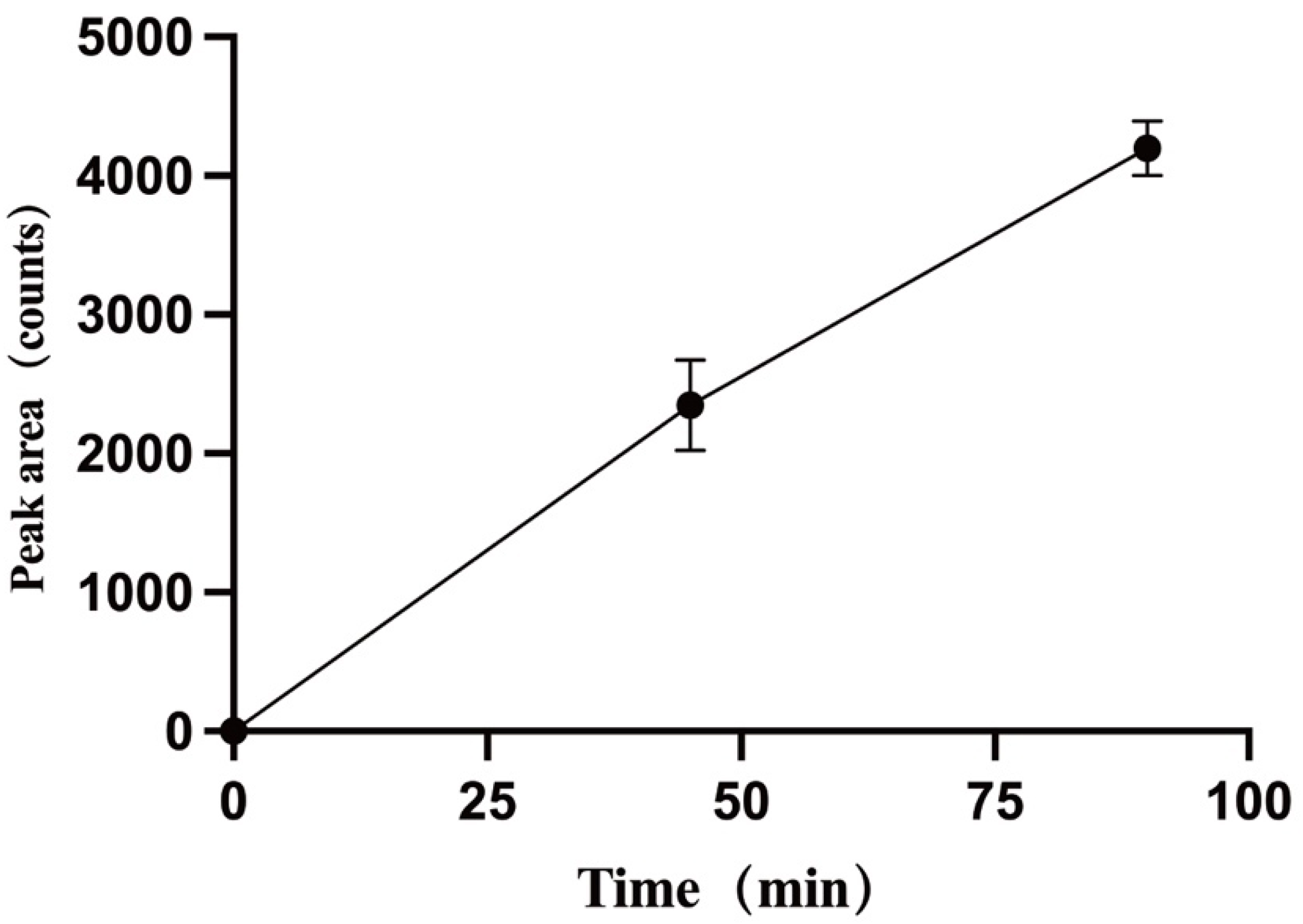

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Systemic Concentration | |
|---|---|---|
| Incubation system | Liver microsomes | 0.5 mg/mL |
| Drug (TPB15) | 5 μM | |
| NADPH | 1 mM | |
| MgCl2 | 3 mM | |
| PBS | 100 mM |
| Inhibitor | Corresponding CYP Metabolic Enzymes | System Concentration (μM) |
|---|---|---|
| Coumarin | CYP2A6 | 25 |
| Quinidine | CYP2D6 | 10 |
| Quercetin | CYP2C8 | 20 |
| α-Naphthoflavone | CYP1A2 | 10 |
| Sulfaphenazole | CYP2C9 | 20 |
| Ketoconazole | CYP3A4 | 1 |
| Thioridazine hydrochloride | CYP2C19 | 25 |
| Indicated Concentration (μg/mL) | Cpre (μg/mL) | Cpost (μg/mL) | NSB (%) |
|---|---|---|---|
| 4 | 4.01 ± 0.03 | 3.94 ± 0.05 | 1.75 ± 0.16 |
| 8 | 7.97 ± 0.09 | 7.75 ± 0.02 | 2.76 ± 0.27 |
| 16 | 16.09 ± 0.11 | 15.66 ± 0.13 | 2.67 ± 0.34 |
| Indicated Concentration (μg/mL) | Plasma Protein Binding (%) | ||||
|---|---|---|---|---|---|
| Human | SD Rat | KM Mice | Cynomolgus Monkey | Beagle Dog | |
| 4 | 81.51 ± 0.44 | 81.70 ± 0.10 | 81.80 ± 0.10 | 81.48 ± 2.00 | 81.48 ± 2.22 |
| 8 | 81.67 ± 0.15 | 80.87 ± 1.08 | 81.21 ± 0.34 | 81.24 ± 0.39 | 81.03 ± 0.51 |
| 16 | 82.15 ± 0.21 | 82.40 ± 0.14 | 82.40 ± 0.42 | 82.20 ± 0.85 | 82.28 ± 1.86 |
| Species * | Regression Equation | R2 | T1/2 (min) | CLint (mL/min/mg) |
|---|---|---|---|---|
| RLM | Y = −0.0028 * X + 4.560 | 0.9218 | 305.15 | 0.0344 |
| MouLM | Y = −0.0022 * X + 4.569 | 0.9346 | 241.55 | 0.0273 |
| MLM | Y = −0.0078 * X + 4.526 | 0.9666 | 88.34 | 0.0941 |
| DLM | Y = −0.0011 * X + 4.591 | 0.9377 | 630.00 | 0.0132 |
| HLM | Y = −0.0067 * X + 4.529 | 0.9307 | 103.25 | 0.0805 |
| Inhibitor | CYP Isoform | Remaining Proportion of Substrate TPB15 (μg/mL) | RSD (%) | Inhibitor Rate (%) |
|---|---|---|---|---|
| Coumarin | CYP2A6 | 1.66 ± 0.12 | 7.32 | 6.60 |
| Quinidine | CYP2D6 | 1.63 ± 0.04 | 2.88 | 4.82 |
| α-Naphthoflavone | CYP1A2 | 1.64 ± 0.05 | 3.55 | 5.58 |
| Quercetin | CYP2C8 | 1.68 ± 0.08 | 4.79 | 8.63 |
| Sulfaphenazole | CYP2C9 | 2.64 ± 0.10 | 3.93 | 81.22 |
| Ketoconazole | CYP3A4 | 2.23 ± 0.15 | 6.83 | 50.25 |
| Thioridazine hydrochloride | CYP2C19 | 1.61 ± 0.10 | 6.11 | 2.79 |
| Positive control group | — | 1.57 ± 0.09 | 1.32 | 0.00 |
| Negative control group | — | 2.88 ± 0.04 | 1.44 | — |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, D.; Chen, B.; Deng, M.; Wu, S.; Xie, S. Plasma Protein Binding, Biostability, Metabolite Profiling, and CYP450 Phenotype of TPB15 Across Different Species: A Novel Smoothened Inhibitor for TNBC Therapy. Pharmaceutics 2025, 17, 423. https://doi.org/10.3390/pharmaceutics17040423
Wen D, Chen B, Deng M, Wu S, Xie S. Plasma Protein Binding, Biostability, Metabolite Profiling, and CYP450 Phenotype of TPB15 Across Different Species: A Novel Smoothened Inhibitor for TNBC Therapy. Pharmaceutics. 2025; 17(4):423. https://doi.org/10.3390/pharmaceutics17040423
Chicago/Turabian StyleWen, Dingsheng, Boyu Chen, Mingtong Deng, Shaoyu Wu, and Shuilin Xie. 2025. "Plasma Protein Binding, Biostability, Metabolite Profiling, and CYP450 Phenotype of TPB15 Across Different Species: A Novel Smoothened Inhibitor for TNBC Therapy" Pharmaceutics 17, no. 4: 423. https://doi.org/10.3390/pharmaceutics17040423
APA StyleWen, D., Chen, B., Deng, M., Wu, S., & Xie, S. (2025). Plasma Protein Binding, Biostability, Metabolite Profiling, and CYP450 Phenotype of TPB15 Across Different Species: A Novel Smoothened Inhibitor for TNBC Therapy. Pharmaceutics, 17(4), 423. https://doi.org/10.3390/pharmaceutics17040423