
Anticancer Nanoparticle Carriers of the Proapoptotic Protein Cytochrome c


Abstract
1. Introduction
1.1. Anticancer Strategies
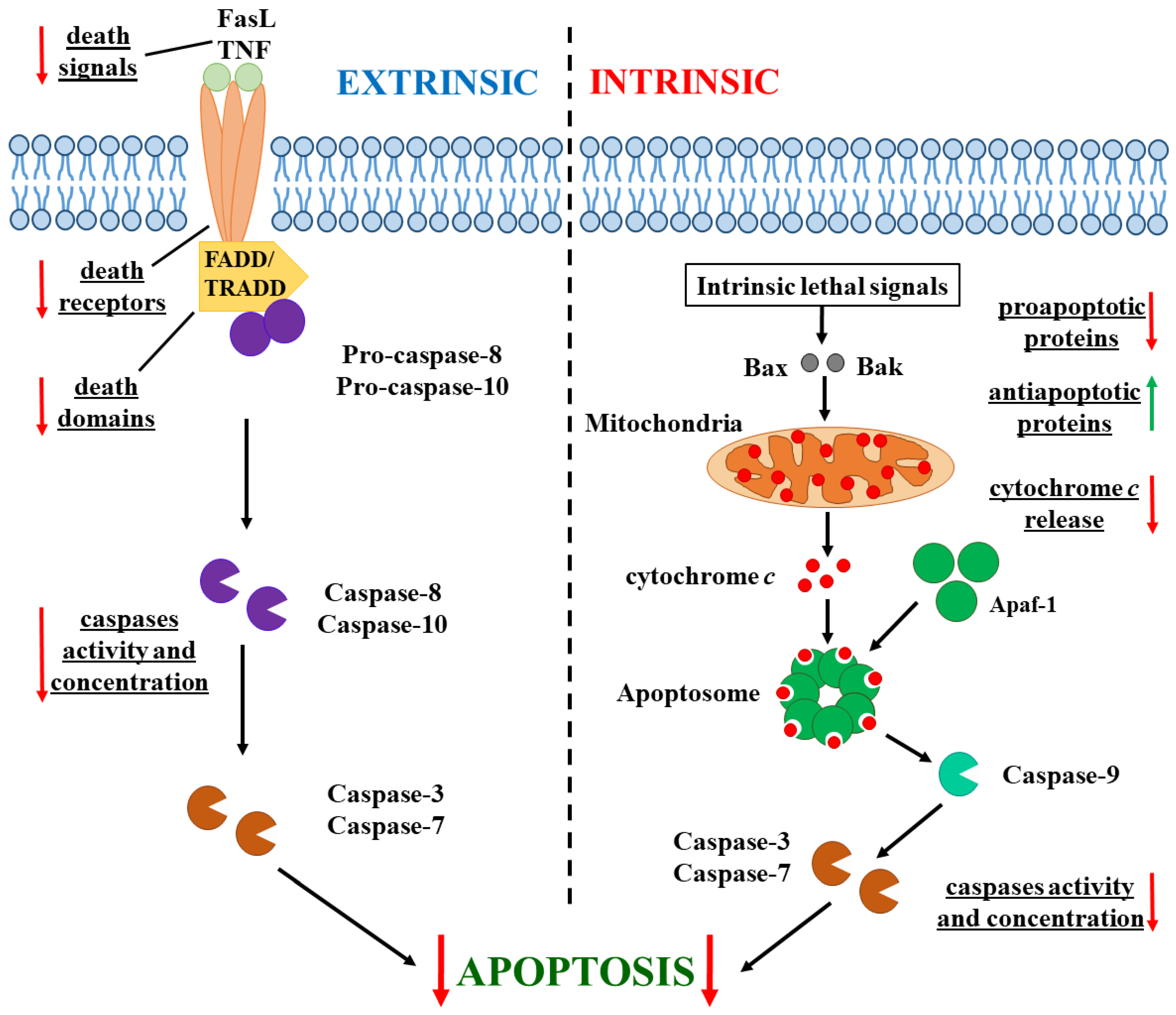
1.2. Apoptosis in Cancer Cells
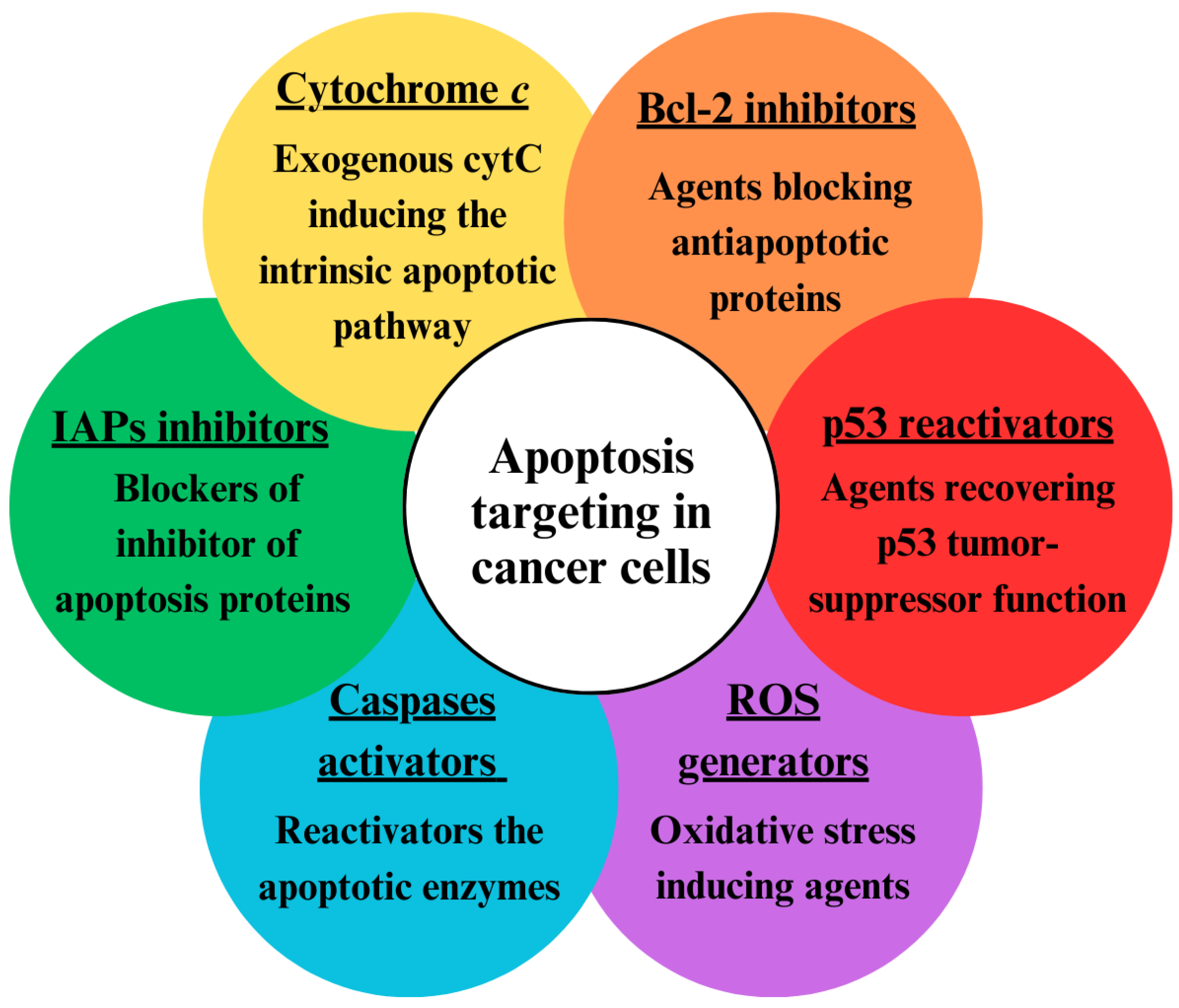
1.3. Apoptosis Targeting in Tumor Cells
2. Nanoparticle Carriers of Exogenous Cytochrome c
2.1. Anticancer Cytochrome-Bearing Particles: Chronology
2.2. Mesoporous Silica Nanoparticles (NPs)
2.3. Montmorillonite Nanoplates
2.4. Gold-ssDNA Nanoparticles
2.5. Hybrid Iron Oxide–Gold Nanoparticles
2.6. Polymeric Nanoparticles
2.7. Cytochrome c Particles
2.7.1. Oleic Acid Bearing cytC Nanoparticles
2.7.2. Hyaluronic Acid Bearing cytC Nanoparticles
2.7.3. Folic Acid Bearing cytC Nanoparticles
2.8. Ferritin Nanocapsules
2.9. Cationic Dextrin
2.10. Particles Bearing cytC: Summary
3. Required Properties for Cytochrome Carriers
3.1. Rate of Endocytosis
3.2. Particle Geometry: Surface, Size, and Shape
3.3. Electric Charge
3.4. Particle Concentration
4. Future Directions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- DeVita, J.; Vincent, T.; Chu, E. A history of cancer chemotherapy. Cancer Res. 2008, 68, 8643–8653. [Google Scholar] [CrossRef] [PubMed]
- Zhivkov, A.M.; Popov, T.T.; Hristova, S.H. Composite hydrogels with included solid-state nanoparticles bearing anticancer chemotherapeutics. Gels 2023, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse effects of cancer chemotherapy: Anything new to improve tolerance and reduce sequelae? Front Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment. Intern. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef]
- Serna, N.; Sánchez-García, L.; Unzueta, U.; Díaz, R.; Vázquez, E.; Mangues, R.; Villaverde, A. Protein-based therapeutic killing for cancer therapies. Trends Biotechnol. 2018, 36, 318–335. [Google Scholar] [CrossRef]
- Brown, J.M. Tumor microenvironment and the response to anticancer therapy. Cancer Biol. Ther. 2002, 1, 453–458. [Google Scholar] [CrossRef]
- Liu, D.; Yang, F.; Xiong, F.; Gu, N. The smart drug delivery system and its clinical potential. Theranostics 2016, 6, 1306. [Google Scholar] [CrossRef]
- Wiesenthal, A.; Hunter, L.; Wang, S.; Wickliffe, J.; Wilkerson, M. Nanoparticles: Small and mighty. Int. J. Dermatol. 2011, 50, 247–254. [Google Scholar] [CrossRef]
- Sun, L.; Liu, H.; Ye, Y.; Lei, Y.; Islam, R.; Tan, S.; Tong, R.; Miao, Y.; Cai, L. Smart nanoparticles for cancer therapy. Signal Transduct. Target. Ther. 2023, 8, 418. [Google Scholar] [CrossRef]
- Kanamala, M.; Wilson, W.R.; Yang, M.; Palmer, B.D.; Wu, Z.J.B. Mechanisms and biomaterials in pH-responsive tumour targeted drug delivery: A review. Biomaterials 2016, 85, 152–167. [Google Scholar] [CrossRef]
- De La Rica, R.; Aili, D.; Stevens, M.M. Enzyme-responsive nanoparticles for drug release and diagnostics. Adv. Drug Deliv. Rev. 2012, 64, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Matsunaga, Y.T. Thermo-responsive polymers and their application as smart biomaterials. J. Mater. Chem. B 2017, 5, 4307–4321. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, F.; Liu, Z.; Wang, L.; Zhang, N. Tumor specific delivery and therapy by double-targeted nanostructured lipid carriers with anti-VEGFR-2 antibody. Mol. Pharm. 2011, 8, 2291–2301. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.F.; Caliri, A.W.; Duex, J.E.; Theodorescu, D. Targetable pathways in advanced bladder cancer: FGFR signaling. Cancers 2021, 13, 4891. [Google Scholar] [CrossRef]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef]
- Murakami, M.; Cabral, H.; Matsumoto, Y.; Wu, S.; Kano, M.R.; Yamori, T.; Nishiyama, N.; Kataoka, K. Improving drug potency and efficacy by nanocarrier-mediated subcellular targeting. Sci. Transl. Med. 2011, 3, ra2–ra64. [Google Scholar] [CrossRef]
- Xia, Y.; Choi, H.K.; Lee, K. Recent advances in hypoxia-inducible factor (HIF)-1 inhibitors. Eur. J. Med. Chem. 2012, 49, 24–40. [Google Scholar] [CrossRef]
- Ameisen, J.C. On the origin, evolution, and nature of programmed cell death: A timeline of four billion years. Cell Death Differ. 2002, 9, 367–393. [Google Scholar] [CrossRef]
- Häcker, G. The morphology of apoptosis. Cell Tissue Res. 2000, 301, 5–17. [Google Scholar] [CrossRef]
- Arandjelovic, S.; Ravichandran, K.S. Phagocytosis of apoptotic cells in homeostasis. Nat. Immunol. 2015, 16, 907–917. [Google Scholar] [CrossRef]
- Carson, D.A.; Ribeiro, J.M. Apoptosis and disease. Lancet 1993, 341, 1251–1254. [Google Scholar] [CrossRef] [PubMed]
- Buja, L.M.; Eigenbrodt, M.L.; Eigenbrodt, E.H. Apoptosis and necrosis. Basic types and mechanisms of cell death. Arch. Pathol. Lab. Med. 1993, 117, 1208–1214. [Google Scholar] [PubMed]
- Reed, J.C. Dysregulation of apoptosis in cancer. J. Clin. Oncol. 1999, 17, 2941. [Google Scholar] [CrossRef] [PubMed]
- Fadeel, B.; Orrenius, S.; Zhivotovsky, B. Apoptosis in human disease: A new skin for the old ceremony. Biochem. Biophys. Res. Commun. 1999, 266, 699–717. [Google Scholar] [CrossRef]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326, 1–16. [Google Scholar] [CrossRef]
- Lavrik, I.; Golks, A.; Krammer, P.H. Death receptor signaling. J. Cell Sci. 2005, 118, 265–267. [Google Scholar] [CrossRef]
- Flusberg, D.A.; Sorger, P.K. Surviving apoptosis: Life–death signaling in single cells. Trends Cell Biol. 2015, 25, 446–458. [Google Scholar] [CrossRef]
- Shen, X.G.; Wang, C.; Li, Y.; Wang, L.; Zhou, B.; Xu, B.; Jiang, X.; Zhou, Z.G.; Sun, X.F. Downregulation of caspase-9 is a frequent event in patients with stage II colorectal cancer and correlates with poor clinical outcome. Color. Dis. 2010, 12, 1213–1218. [Google Scholar] [CrossRef]
- Pepper, C.; Hoy, T.; Bentley, D.P. Bcl-2/Bax ratios in chronic lymphocytic leukaemia and their correlation with in vitro apoptosis and clinical resistance. Br. J. Cancer 1997, 76, 935–938. [Google Scholar] [CrossRef]
- Lopes, R.B.; Gangeswaran, R.; McNeish, I.A.; Wang, Y.; Lemoine, N.R. Expression of the IAP protein family is dysregulated in pancreatic cancer cells and is important for resistance to chemotherapy. Int. J. Cancer 2007, 120, 2344–2352. [Google Scholar] [CrossRef]
- Waterhouse, N.J.; Green, D.R. Mitochondria and apoptosis: HQ or high-security prison? J. Clin. Immunol. 1999, 19, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Hassan, M.I.; Islam, A.; Ahmad, F. The role of key residues in structure, function, and stability of cytochrome-c. Cell. Mol. Life Sci. 2014, 71, 229–255. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, D.H.; Neupert, W. Biogenesis of mitochondrial c-type cytochromes. J. Bioenerg. Biomembr. 1990, 22, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef]
- Kang, M.H.; Reynolds, C.P. Bcl-2 inhibitors: Targeting mitochondrial apoptotic pathways in cancer therapy. Clin. Cancer Res. 2009, 15, 1126–1132. [Google Scholar] [CrossRef]
- Philchenkov, A.; Zavelevich, M.; Kroczak, T.J.; Los, M.J. Caspases and cancer: Mechanisms of inactivation and new treatment modalities. Exp. Oncol. 2004, 26, 82–97. [Google Scholar]
- Chène, P. p53 as a drug target in cancer therapy. Expert Opin. Ther. Pat. 2001, 11, 923–935. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Shiraki, K.; Fuke, H.; Inoue, T.; Miyashita, K.; Yamanaka, Y.; Saitou, Y.; Sugimoto, K.; Nakano, T. Targeting of X-linked inhibitor of apoptosis protein or survivin by short interfering RNAs sensitize hepatoma cells to TNF-related apoptosis-inducing ligand-and chemotherapeutic agent-induced cell death. Oncol. Rep. 2005, 14, 1311–1316. [Google Scholar] [CrossRef]
- Dong, Q.; Jiang, Z. Platinum–Iron Nanoparticles for Oxygen-Enhanced Sonodynamic Tumor Cell Suppression. Inorganics 2024, 12, 331. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, L.; Guo, Z.; Sun, Y.; Yan, J. A pH-sensitive imidazole grafted polymeric micelles nanoplatform based on ROS amplification for ferroptosis-enhanced chemodynamic therapy. Colloids Surf. B Biointerfaces 2024, 237, 113871. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, J. Cytochrome c in cancer therapy and prognosis. Biosci. Rep. 2022, 42, BSR20222171. [Google Scholar] [CrossRef] [PubMed]
- Purring-Koch, C.; McLendon, G. Cytochrome c binding to Apaf-1: The effects of dATP and ionic strength. Proc. Natl. Acad. Sci. USA 2000, 97, 11928–11931. [Google Scholar] [CrossRef]
- Yuan, S.; Topf, M.; Reubold, T.F.; Eschenburg, S.; Akey, C.W. Changes in Apaf-1 conformation that drive apoptosome assembly. Biochemistry 2013, 52, 2319–2327. [Google Scholar] [CrossRef]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef]
- Zou, H.; Li, Y.; Liu, X.; Wang, X. An APAF-1: Cytochrome c multimeric complex is a functional apoptosome that activates procaspase-9. J. Biol. Chem. 1999, 274, 11549–11556. [Google Scholar] [CrossRef]
- Monian, P.; Jiang, X. Clearing the final hurdles to mitochondrial apoptosis: Regulation post cytochrome c release. Exp. Oncol. 2012, 7, 185–191. [Google Scholar]
- Zhivotovsky, B.; Orrenius, S.; Brustugun, O.T.; Døskeland, S.O. Injected cytochrome c induces apoptosis. Nature 1998, 391, 449–450. [Google Scholar] [CrossRef]
- Kole, A.J.; Knight, E.R.W.; Deshmukh, M. Activation of apoptosis by cytoplasmic microinjection of cytochrome c. J. Vis. Exp. 2011, 1, 2773. [Google Scholar]
- Chang, S.H.; Phelps, P.C.; Berezesky, I.K.; Ebersberger, M.L.; Trump, B.F. Studies on the mechanisms and kinetics of apoptosis induced by microinjection of cytochrome c in rat kidney tubule epithelial cells (NRK-52E). Am. J. Pathol. 2000, 156, 637–649. [Google Scholar] [CrossRef]
- Sell, M.; Lopes, A.R.; Escudeiro, M.; Esteves, B.; Monteiro, A.R.; Trindade, T.; Cruz-Lopes, L. Application of nanoparticles in cancer treatment: A concise review. Nanomaterials 2023, 13, 2887. [Google Scholar] [CrossRef]
- Yao, Y.; Zhou, Y.; Liu, L.; Xu, Y.; Chen, Q.; Wang, Y.; Wu, S.; Deng, Y.; Zhang, J.; Shao, A. Nanoparticle-based drug delivery in cancer therapy and its role in overcoming drug resistance. Front. Mol. Biosci. 2020, 7, 193. [Google Scholar] [CrossRef]
- Slowing, I.I.; Trewyn, B.G.; Lin, V.S.Y. Mesoporous silica nanoparticles for intracellular delivery of membrane-impermeable proteins. J. Am. Chem. Soc. 2007, 129, 8845–8849. [Google Scholar] [CrossRef]
- Santra, S.; Kaittanis, C.; Perez, J.M. Cytochrome c encapsulating theranostic nanoparticles: A novel bifunctional system for targeted delivery of therapeutic membrane-impermeable proteins to tumors and imaging of cancer therapy. Mol. Pharm. 2010, 7, 185–191. [Google Scholar] [CrossRef]
- Kim, S.K.; Foote, M.B.; Huang, L. The targeted intracellular delivery of cytochrome c protein to tumors using lipid-apolipoprotein nanoparticles. Biomaterials 2012, 33, 3959–3966. [Google Scholar] [CrossRef]
- Imesch, P.; Scheiner, D.; Szabo, E.; Fink, D.; Fedier, A. Conjugates of cytochrome c and antennapedia peptide activate apoptosis and inhibit proliferation of HeLa cancer cells. Exp. Ther. Med. 2013, 6, 786–790. [Google Scholar] [CrossRef]
- Ng, D.Y.W.; Fahrer, J.; Wu, Y.; Eisele, K.; Kuan, S.L.; Barth, H.; Weil, T. Efficient delivery of p53 and cytochrome c by supramolecular assembly of a dendritic multi-domain delivery system. Adv. Healthc. Mater. 2013, 2, 1620–1629. [Google Scholar] [CrossRef]
- Morales-Cruz, M.; Figueroa, C.M.; González-Robles, T.; Delgado, Y.; Molina, A.; Méndez, J.; Morales, M.; Griebenow, K. Activation of caspase-dependent apoptosis by intracellular delivery of cytochrome c-based nanoparticles. J. Nanobiotechnol. 2014, 12, 1–11. [Google Scholar] [CrossRef]
- Malekigorji, M.; Hoskins, C.; Curtis, T.; Varbiro, G. Enhancement of the cytotoxic effect of anticancer agent by cytochrome c functionalized hybrid nanoparticles in hepatocellular cancer cells. J. Nanomed. Res. 2014, 1, 00010. [Google Scholar]
- Barnett, C.M.; Gueorguieva, M.; Lees, M.R.; McGarvey, D.J.; Darton, R.J.; Hoskins, C. Effect of hybrid composition on the physicochemical properties and morphology of iron oxide-gold nanoparticles. J. Nanopart. Res. 2012, 14, 1170. [Google Scholar] [CrossRef]
- Zhang, B.; Luo, Z.; Liu, J.; Ding, X.; Li, J.; Cai, K. Cytochrome c end-capped mesoporous silica nanoparticles as redox-responsive drug delivery vehicles for liver tumor-targeted triplex therapy in vitro and in vivo. J. Control. Release 2014, 192, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Teng, Z.; Liu, Y.; Tian, Y.; Sun, J.; Wang, S.; Wang, C.; Wang, J.; Lu, G. Cytochrome C capped mesoporous silica nanocarriers for pH-sensitive and sustained drug release. J. Mater. Chem. B 2014, 2, 4356–4362. [Google Scholar] [CrossRef] [PubMed]
- Méndez, J.; Morales Cruz, M.; Delgado, Y.; Figueroa, C.M.; Orellano, E.A.; Morales, M.; Monteagudo, A.; Griebenow, K. Delivery of chemically glycosylated cytochrome c immobilized in mesoporous silica nanoparticles induces apoptosis in HeLa cancer cells. Mol. Pharm. 2014, 11, 102–111. [Google Scholar] [CrossRef]
- Yeh, T.H.; Wu, F.L.L.; Shen, L.J. Intracellular delivery of cytochrome c by galactosylated albumin to hepatocarcinoma cells. J. Drug Target. 2014, 22, 528–535. [Google Scholar] [CrossRef]
- Koo, A.N.; Min, K.H.; Lee, H.J.; Jegal, J.H.; Lee, J.W.; Lee, S.C. Calcium carbonate mineralized nanoparticles as an intracellular transporter of cytochrome c for cancer therapy. Chem Asian J. 2015, 10, 2380–2387. [Google Scholar] [CrossRef]
- Delgado, Y.; Morales-Cruz, M.; Hernández-Román, J.; Hernández, G.; Griebenow, K. Development of HAMLET-like cytochrome c–oleic acid nanoparticles for cancer therapy. J. Nanomed. Nanotechnol. 2015, 6, 1. [Google Scholar]
- Morales-Cruz, M.; Cruz-Montañez, A.; Figueroa, C.M.; González-Robles, T.; Davila, J.; Inyushin, M.; Loza-Rosas, S.A.; Molina, A.M.; Muñoz-Perez, L.; Kucheryavykh, L.Y.; et al. Combining stimulus-triggered release and active targeting strategies improves cytotoxicity of cytochrome c nanoparticles in tumor cells. Mol. Pharm. 2016, 13, 2844–2854. [Google Scholar] [CrossRef]
- Vladimirov, Y.A.; Sarisozen, C.; Vladimirov, G.K.; Filipczak, N.; Polimova, A.M.; Torchilin, V.P. The cytotoxic action of cytochrome C/Cardiolipin nanocomplex (Cyt-CL) on cancer cells in culture. Pharm. Res. 2017, 34, 1264–1275. [Google Scholar] [CrossRef]
- Figueroa, C.M.; Suárez, B.N.; Molina, A.M.; Fernández, J.C.; Torres, Z.; Griebenow, K. Smart release nano-formulation of cytochrome c and hyaluronic acid induces apoptosis in cancer cells. J. Nanomed. Nanotechnol. 2017, 8, 1. [Google Scholar]
- Al-Shakarchi, W.; Alsuraifi, A.; Abed, M.; Abdullah, M.; Richardson, A.; Curtis, A.; Hoskins, C. Combined effect of anticancer agents and cytochrome c decorated hybrid nanoparticles for liver cancer therapy. Pharmaceutics 2018, 10, 48. [Google Scholar] [CrossRef]
- Pei, Y.; Li, M.; Hou, Y.; Hu, Y.; Chu, G.; Dai, L.; Li, K.; Xing, Y.; Tao, B.; Yu, Y.; et al. An autonomous tumor-targeted nanoprodrug for reactive oxygen species-activatable dual-cytochrome c/doxorubicin antitumor therapy. Nanoscale 2018, 10, 11418–11429. [Google Scholar] [CrossRef] [PubMed]
- Macone, A.; Masciarelli, S.; Palombarini, F.; Quaglio, D.; Boffi, A.; Trabuco, M.C.; Baiocco, P.; Fazi, F. Ferritin nanovehicle for targeted delivery of cytochrome c to cancer cells. Sci. Rep. 2019, 9, 11749. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, W.J.; Park, S.; Choi, D.; Kim, S.; Park, N. Reversibly pH-responsive gold nanoparticles and their applications for photothermal cancer therapy. Sci. Rep. 2019, 9, 20180. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; Lim, D.K.; Kim, S. Hydrolytic surface erosion of mesoporous silica nanoparticles for efficient intracellular delivery of cytochrome c. J. Colloid Interface Sci. 2020, 560, 416–425. [Google Scholar] [CrossRef]
- Hristova, S.H. Electric Properties and Anticancer Effect of Cytochrome c Adsorbed on Colloid Particles from Montmorillonite. Ph.D. Thesis, Institute of Physical Chemistry, Bulgarian Academy of Sciences, Sofia, Bulgaria, 2019. [Google Scholar]
- Hristova, S.H.; Zhivkov, A.M. Cytotoxic effect of exogenous cytochrome c adsorbed on montmorillonite colloid particles on colon cancer cell culture. Compt. Rend. Acad. Bulg. Sci. 2019, 72, 198–203. [Google Scholar]
- Hristova, S.H.; Zhivkov, A.M. Montmorillonite colloid plates with adsorbed cytochrome c: In vitro cytotoxic effect on colon cancer cell culture. Cancer Nanotechnol. 2021, 12, 23. [Google Scholar] [CrossRef]
- Hristova, S.H.; Zhivkov, A.M. Protein–mineral composite particles with logarithmic dependence of anticancer cytotoxicity on concentration of montmorillonite nanoplates with adsorbed cytochrome c. Pharmaceutics 2023, 15, 386. [Google Scholar] [CrossRef]
- Barcelo-Bovea, V.; Dominguez-Martinez, I.; Joaquin-Ovalle, F.; Amador, L.A.; Castro-Rivera, E.; Medina-Álvarez, K.; McGoron, A.; Griebenow, K.; Ferrer-Acosta, Y. Optimization and characterization of protein nanoparticles for the targeted and smart delivery of cytochrome c to non-small cell lung carcinoma. Cancers 2020, 12, 1215. [Google Scholar] [CrossRef]
- Sun, X.S.; Jang, M.S.; Fu, Y.; Lee, J.H.; Lee, D.S.; Li, Y.; Yang, H.Y. Intracellular delivery of cytochrome C using hypoxia-responsive polypeptide micelles for efficient cancer therapy. Mater. Sci. Eng. C 2020, 114, 111069. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, F.; Wen, H.; Shi, W.; Huang, Q.; Huang, Y.; Xie, J.; Li, P.; Chen, J.; Qin, L. Tumor- and mitochondria-targeted nanoparticles eradicate drug-resistant lung cancer through mitochondrial pathway of apoptosis. J. Nanobiotechnol. 2020, 18, 8. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, Y.; Li, Y.; Zhang, L.; Jiang, H.; Tao, J.; Zhu, J. Gold nanoparticle-guarded large-pore mesoporous silica nanocomposites for delivery and controlled release of cytochrome c. J. Colloid Interface Sci. 2021, 589, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Martinez, I.; Joaquin-Ovalle, F.; Ferrer-Acosta, Y.; Griebenow, K.H. Folate-decorated cross-linked cytochrome c nanoparticles for active targeting of non-small cell lung carcinoma (NSCLC). Pharmaceutics 2022, 14, 490. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Jang, M.S.; Liu, C.; Lee, J.H.; Li, Y.; Yang, H.Y. Hypoxia-responsive hyaluronic acid nanogels with improved endo/lysosomal escape ability for tumor-targeted cytochrome c delivery. Eur. Polym. J. 2022, 173, 111259. [Google Scholar] [CrossRef]
- Sarkar, A.; Sarkhel, S.; Bisht, D.; Jaiswal, A. Cationic dextrin nanoparticles for effective intracellular delivery of cytochrome C in cancer therapy. RSC Chem. Biol. 2024, 5, 249–261. [Google Scholar] [CrossRef]
- Zhivkov, A.M.; Hristova, S.H.; Popov, T.T. Composition of Proapoptotic Montmorillonite-Cytochrome c Nanoplates with High Anticancer Cytotoxicity: Monolayer Adsorption Approach. In Interdisciplinary Cancer Research; Springer Nature: Cham, Switzerland, 2024. [Google Scholar]
- Hristova, S.H.; Zhivkov, A.M. Asorption of cytochrome c on montmorillonite nanoplates: Protein concentration dependence. J. Colloid Interface Sci. 2015, 446, 252–262. [Google Scholar] [CrossRef]
- Hristova, S.H.; Zhivkov, A.M. Isoelectric point of free and adsorbed cytochrome c determined by various methods. Colloids Surf. B Biointerfaces 2019, 174, 87–94. [Google Scholar] [CrossRef]
- Hristova, S.H.; Zhivkov, A.M. Electrooptical determination of the isoelectric point of globular proteins: Cytochrome c adsorbed on montmorillonite nanoplates. Colloids Surf. B Biointerfaces 2019, 176, 480–487. [Google Scholar] [CrossRef]
- Hristova, S.H.; Zhivkov, A.M. Intermolecular Electrostatic Interactions in Cytochrome c Protein Monolayer on Montmorillonite Alumosilicate Surface: A Positive Cooperative Effect. Int. J. Mol. Sci. 2024, 25, 6834. [Google Scholar] [CrossRef]
- Hoskins, C.; Min, Y.; Gueorguieva, M.; McDougall, C.; Volovick, A.; Prentice, P.; Wang, L. Hybrid gold-iron oxide nanoparticles as a multifunctional platform for biomedical application. J. Nanobiotechnology 2012, 10, 27. [Google Scholar] [CrossRef]
- Barnett, C.M.; Gueorguieva, M.; Lees, M.R.; McGarvey, D.J.; Hoskins, C. Physical stability, biocompatibility and potential use of hybrid iron oxide-gold nanoparticles as drug carriers. J. Nanopart. Res. 2013, 15, 1706. [Google Scholar] [CrossRef]
- Ma, W.; Saccardo, A.; Roccatano, D.; Aboagye-Mensah, D.; Alkaseem, M.; Jewkes, M.; Di Nezza, F.; Baron, M.; Soloviev, M.; Ferrari, E. Modular assembly of proteins on nanoparticles. Nat. Commun. 2018, 9, 1489. [Google Scholar] [CrossRef] [PubMed]
- Collinson, M.; Bowden, E.F.; Tarlov, M.J. Voltammetry of covalently immobilized cytochrome c on self-assembled monolayer electrodes. Langmuir 1992, 8, 1247–1250. [Google Scholar] [CrossRef]
- Boulos, S.P.; Davis, T.A.; Yang, J.A.; Lohse, S.E.; Alkilany, A.M.; Holland, L.A.; Murphy, C.J. Nanoparticle-protein interactions: A thermodynamic and kinetic study of the adsorption of bovine serum albumin to gold nanoparticle surfaces. Langmuir 2013, 29, 14984–14996. [Google Scholar] [CrossRef] [PubMed]
- Cumberland, S.L.; Strouse, G.F. Analysis of the nature of oxyanion adsorption on gold nanomaterial surfaces. Langmuir 2002, 18, 269–276. [Google Scholar] [CrossRef]
- Enustun, B.V.; Turkevick, J. Coagulation of colloidal gold. J. Am. Chem. Soc. 1963, 85, 3317–3328. [Google Scholar] [CrossRef]
- Lee, H.H.; Yamaoka, S.; Murayama, N.; Shibata, J. Dispersion of Fe3O4 suspensions using sodium dodecylbenzene sulphonate as dispersant. Mater. Lett. 2007, 61, 3974–3977. [Google Scholar] [CrossRef]
- Andre, N.; Braguer, D.; Brasseur, G.; Goncalves, A.; Lemesle-Meunier, D.; Guise, S.; Jordan, M.A.; Briand, C. Paclitaxel induces release of cytochrome c from mitochondria isolated from human neuroblastoma cells’. Cancer Res. 2000, 60, 5349–5353. [Google Scholar]
- Méndez, J.; Monteagudo, A.; Griebenow, K. Stimulus-responsive controlled release system by covalent immobilization of an enzyme into mesoporous silica nanoparticles. Bioconjug Chem. 2012, 23, 698–704. [Google Scholar] [CrossRef]
- Delgado, Y.; Morales-Cruz, M.; Hernández-Román, J.; Martínez, Y.; Griebenow, K. Chemical glycosylation of cytochrome c improves physical and chemical protein stability. BMC Biochem. 2014, 15, 16. [Google Scholar] [CrossRef]
- Delgado, Y.; Morales-Cruz, M.; Figueroa, C.M.; Hernández-Román, J.; Hernández, G.; Griebenow, K. The cytotoxicity BAMLET complexes is due to oleic acid and independent of the α-lactalbumin component. FEBS Open Bio. 2015, 5, 397–404. [Google Scholar] [CrossRef]
- Maeda, H.; Nakamura, H.; Fang, J. The EPR effect for macromolecular drug delivery to solid tumors: Improvement of tumor uptake, lowering of systemic toxicity, and distinct tumor imaging in vivo. Adv. Drug Deliv. Rev. 2013, 65, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H. Macromolecular therapeutics in cancer treatment: The EPR effect and beyond. J. Control Release 2012, 164, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Sankar, C.; Rajasree, P.H. Human serum albumin nanoparticles for enhanced drug delivery to treat breast cancer: Preparation and in vitro assessment. Int. J. Pharm. Life Sci. 2012, 3, 2055–2063. [Google Scholar]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A multifunctional cell surface adhesion receptor is a regulator of progression and metastases of cancer cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef]
- Figueroa, C.M.; Morales-Cruz, M.; Suárez, B.N.; Fernández, J.C.; Molina, A.M.; Quiñones, C.M.; Griebenow, K. Induction of cancer cell death by hyaluronic acid-mediated uptake of cytochrome c. J. Nanomed. Nanotechnol. 2015, 6, 316. [Google Scholar]
- Naor, D. Encyclopedia of Immunology; Elsevier: Amsterdam, The Netherlands, 1998; ISBN 0-12-226765-6. [Google Scholar]
- Croce, M.A.; Boraldi, F.; Quaglino, D.; Tiozzo, R.; Pasquali-Ronchetti, I. Hyaluronan uptake by adult human skin fibroblasts in vitro. Europ. J. Histochem. 2003, 47, 63–74. [Google Scholar] [CrossRef]
- Kucheryavykh, Y.V.; Davila, J.; Ortiz-Rivera, J.; Inyushin, M.; Almodovar, L.; Mayol, M.; Morales-Cruz, M.; Cruz-Montañez, A.; Barcelo-Bovea, V.; Griebenow, K.; et al. Targeted delivery of nanoparticulate cytochrome C into glioma cells through the proton-coupled folate transporter. Biomolecules 2019, 9, 154. [Google Scholar] [CrossRef]
- Scaranti, M.; Cojocaru, E.; Banerjee, S.; Banerji, U. Exploiting the folate receptor α in oncology. Nat. Rev. Clin. Oncol. 2020, 17, 349–359. [Google Scholar] [CrossRef]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA methylation: A review of molecular mechanisms and the evidence for folate’s role. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef]
- O’Shannessy, D.J.; Yu, G.; Smale, R.; Fu, Y.-S.; Singhal, S.; Thiel, R.P.; Somers, E.B.; Vachani, A. Folate receptor alpha expression in lung cancer: Diagnostic and prognostic significance. Oncotarget 2012, 3, 414. [Google Scholar] [CrossRef]
- Shulpekova, Y.; Nechaev, V.; Kardasheva, S.; Sedova, A.; Kurbatova, A.; Bueverova, E.; Kopylov, A.; Malsagova, K.; Dlamini, J.C.; Ivashkin, V. The Concept of Folic Acid in Health and Disease. Molecules 2021, 26, 3731. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Harashima, H. Mitochondrial drug delivery systems for macromolecules and their therapeutic application to mitochondrial diseases. Adv. Drug Deliv. Rev. 2008, 60, 1439–1462. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.; Gogada, R.; O’Malley, J.; Gundampati, R.K.; Jayanthi, S.; Hashmi, S.; Lella, R.; Zhang, D.; Wang, J.; Kumar, R.; et al. Molecular insights on cytochrome c and nucleotide regulation of apoptosome function and its implication in cancer. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118573. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Torchilin, V.P. Nanopreparations for organelle-specific delivery in cancer. Adv. Drug Deliv. Rev. 2014, 66, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Jennifer, B.; Berg, V.; Modak, M.; Puck, A.; Seyerl-Jiresch, M.; Künig, S.; Zlabinger, G.J.; Steinberger, P.; Chou, J.; Geha, R.S.; et al. Transferrin receptor 1 is a cellular receptor for human heme-albumin. Commun Biol. 2020, 3, 621. [Google Scholar] [CrossRef]
- Candelaria, P.V.; Leoh, L.S.; Penichet, M.L.; Daniels-Wells, T.R. Antibodies targeting the transferrin receptor 1 (TfR1) as direct anti-cancer agents. Front. Immunol. 2021, 12, 607692. [Google Scholar] [CrossRef]
- Dickerson, R.E.; Timkovich, R. Cytochromes c. Enzym. 1975, 11, 397–547. [Google Scholar]
- Paul, K.G. The stability of cytochrome c at extreme pH values. Acta Chem. Scand. 1948, 10, 430–439. [Google Scholar] [CrossRef]
- Kuroda, Y.; Kidokoro, S.I.; Wada, A. Thermodynamic characterization of cytochrome c at low pH: Observation of the molten globule state and of the cold denaturation process. J. Mol. Biol. 1992, 223, 1139–1153. [Google Scholar] [CrossRef]
- Cinelli, S.; Spinozzi, F.; Itri, R.; Finet, S.; Carsughi, F.; Onori, G.; Mariani, P. Structural characterization of the pH-denatured states of ferricytochrome-c by synchrotron small angle X-Ray scattering. Biophys. J. 2001, 81, 3522–3533. [Google Scholar] [CrossRef]
- Endo, S.; Nagayama, K.; Wada, A. Probing stability and dynamics of proteins by protease digestion I: Comparison of protease susceptibility and thermal stability of cytochrome c. J. Biomol. Struct. Dyn. 1985, 3, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Wang, W.; Zhao, B.; Zhang, S.; Miao, J. Chloroquine inhibits cell growth and induces cell death in A549 lung cancer cells. Bioorg. Med. Chem. 2006, 14, 3218–3222. [Google Scholar] [CrossRef] [PubMed]
- Varkouhi, A.K.; Scholte, M.; Storm, G.; Haisma, H.J. Endosomal escape pathways for delivery of biologicals. J. Control. Release 2011, 151, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Al-Bari, M.A.A. Targeting endosomal acidification by chloroquine analogs as a promising strategy for the treatment of emerging viral diseases. Pharmacol. Res. Perspect. 2017, 5, e00293. [Google Scholar] [CrossRef]
- Hajimolaali, M.; Mohammadian, H.; Torabi, A.; Shirini, A.; Shal, M.K.; Nezhad, H.B.; Iranpour, S.; Eftekhari, R.B.; Dorkoosh, F. Application of chloroquine as an endosomal escape enhancing agent: New frontiers for an old drug. Expert Opin. Drug Delivery 2021, 18, 877–889. [Google Scholar] [CrossRef]
- Vinu, A.; Murugesan, V.; Tangermann, O.; Hartmann, M. Adsorption of cytochrome c on mesoporous molecular sieves: Influence of pH, pore diameter, and aluminum incorporation. Chem. Mater. 2004, 16, 3056–3065. [Google Scholar] [CrossRef]
- Vinu, A.; Streb, C.; Murugesan, V.; Hartmann, M. Adsorption of cytochrome C on new mesoporous carbon molecular sieves. J. Phys. Chem. B 2003, 107, 8297–8299. [Google Scholar] [CrossRef]
- Moosavi, M. Bentonite clay as a natural remedy: A brief review. Iran. J. Public Health 2017, 46, 1176. [Google Scholar]
- Couvreur, P.; Puisieux, F. Nano- and microparticles for the delivery of polypeptides and proteins. Adv. Drug Deliv. Rev. 1993, 10, 141–162. [Google Scholar] [CrossRef]
- Cortez, C.; Tomaskovic-Crook, E.; Johnston, A.P.R.; Scott, A.M.; Nice, E.C.; Heath, J.K.; Caruso, F. Influence of size, surface, cell line, and kinetic properties on the specific binding of A33 antigen-targeted multilayered particles and capsules to colorectal cancer cells. ACS Nano. 2007, 1, 93–102. [Google Scholar] [CrossRef]
- Yin Win, K.; Feng, S.S. Effects of particle size and surface coating on cellular uptake of polymeric nanoparticles for oral delivery of anticancer drugs. Biomaterials 2005, 26, 2713–2722. [Google Scholar] [CrossRef] [PubMed]
- Sakhtianchi, R.; Minchin, R.F.; Lee, K.-B.; Alkilany, A.M.; Serpooshan, V.; Mahmoudi, M. Exocytosis of nanoparticles from cells: Role in cellular retention and toxicity. Adv. Colloid Interface Sci. 2013, 201, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Gratton, S.E.A.; Ropp, P.A.; Pohlhaus, P.D.; Luft, J.C.; Madden, V.J.; Napier, M.E.; DeSimone, J.M. The effect of particle design on cellular internalization pathways. PNAS 2008, 105, 11613–11618. [Google Scholar] [CrossRef]
- Banerjee, A.; Qi, J.; Gogoi, R.; Wong, J.; Mitragotri, S. Role of nanoparticle size, shape and surface chemistry in oral drug delivery. J. Control. Release 2016, 238, 176–185. [Google Scholar] [CrossRef]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef]
- Chung, T.H.; Wu, S.H.; Yao, M.; Lu, C.W.; Lin, Y.S.; Hung, Y.; Mou, C.Y.; Chen, Y.C.; Huang, D.M. The effect of surface charge on the uptake and biological function of mesoporous silica nanoparticles in 3T3-L1 cells and human mesenchymal stem cells. Biomaterials 2007, 28, 2959–2966. [Google Scholar] [CrossRef]
- Foged, C.; Brodin, B.; Frokjaer, S.; Sundblad, A. Particle size and surface charge affect particle uptake by human dendritic cells in an in vitro model. Int. J. Pharm. 2005, 298, 315–322. [Google Scholar] [CrossRef]
- Chen, L.; McCrate, J.M.; Lee, J.C.-M.; Li, H. The role of surface charge on the uptake and biocompatibility of hydroxyapatite nanoparticles with osteoblast cells. Nanotechnology 2011, 22, 105708. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, M.; Portney, N.G.; Cui, D.; Budak, G.; Ozbay, E.; Ozkan, M.; Ozkan, C.S. Zeta potential: A surface electrical characteristic to probe the interaction of nanoparticles with normal and cancer human breast epithelial cells. Biomed. Microdevices 2008, 10, 321–328. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Substance | Shape | Size [nm] | Zeta Potential [mV] | Type Connection | Year | Reference | |
|---|---|---|---|---|---|---|---|
| 1 | Silica (SiO2) | spherical | 265 | – | noncovalent | 2007 | [53] |
| 2 | Hyperbranched polyhydroxyl polymer | spherical | 63 ± 2 | – | noncovalent | 2010 | [54] |
| 3 | Apolipoprotein | spherical | 20–30 | +2–3 | – | 2012 | [55] |
| 4 | Antennapedia (peptide) | – | – | – | covalent | 2013 | [56] |
| 5 | Streptavidin, Biotin | spherical | 7.06 | – | covalent | 2013 | [57] |
| 6 | Poly(lactic-co-glycolic) acid | spherical | 100–300 | – | covalent | 2014 | [58] |
| 7 | Iron oxide and Gold | spherical | 115–200 | +10.0 | covalent | 2014 | [59,60] |
| 8 | Silica (SiO2) | spherical | 160–170 | – | covalent | 2014 | [61] |
| 9 | Silica (SiO2) | spherical | 117 ± 10 | –14.4 (pH 7.4) –0.5 (pH 5) | – | 2014 | [62] |
| 10 | Silica (SiO2) | spherical | 356 ± 56 | – | covalent | 2014 | [63] |
| 11 | Galactosylated albumin | spherical | 13.6 ± 2 | – | covalent | 2014 | [64] |
| 12 | Calcium carbonate, PEG | spherical | 200–250 | −13.7 | noncovalent | 2015 | [65] |
| 13 | cytC, Oleic acid | spherical | 122.9 ± 5.5 | −0.489 ± 3.2 | noncovalent | 2015 | [66] |
| 14 | cytC, Folic acid | spherical | 338 ± 8 | +47.5 | covalent | 2016 | [67] |
| 15 | Cardiolipin | spherical | 93.8 ± 1.35 | −61.1 ± 1.2 | noncovalent | 2017 | [68] |
| 16 | cytC, Hyaluronic acid | spherical | 542 ± 9 | −28.7 ± 0.6 | covalent | 2017 | [69] |
| 17 | Iron oxide, Gold | spherical | 115–200 | +10.0 | covalent | 2018 | [70] |
| 18 | Silica (SiO2) | spherical | 163.3 ± 26.84 | – | covalent | 2018 | [71] |
| 19 | Ferritin | spherical | – | – | covalent | 2019 | [72] |
| 20 | Gold, ssDNA | spherical | 12 (pH 7.4) 600 (pH 5.5) | −25 (pH 7.4) +3 (pH 5.5) | noncovalent | 2019 | [73] |
| 21 | Silica (SiO2) | spherical | 73.7 ± 9.2 | −16.3 ± 1.0 | – | 2019 | [74] |
| 22 | Montmorillonite | monolayer | 430 | +20.0 (pH 6) | noncovalent | 2019 2021 | [75,76,77,78] |
| 23 | cytC, Folic acid | spherical | 253 ± 55 354 ± 11 | +26.9 ± 5.03 +22.4 ± 6.36 | covalent | 2020 | [79] |
| 24 | Methoxy-PEG-block- -copolymer | spherical | 96.3 | +15 | – | 2020 | [80] |
| 25 | Hyaluronic acid, Triphenylphosphonium | spherical | 140 | −24.65 | covalent | 2020 | [81] |
| 26 | Silica, DMMA, Gold | spherical | 200 | −15.4 ± 0.3 | noncovalent | 2021 | [82] |
| 27 | cytC, Folic acid | spherical | 169 ± 9 | +17.7 ± 1.7 | covalent | 2022 | [83] |
| 28 | PEG, Hyaluronic acid | spherical | 50 | −10 | noncovalent | 2022 | [84] |
| 29 | Cationic dextrin | spherical | 100 ± 30 | +6.3 | noncovalent | 2024 | [85] |
| Nanoparticles | Cancer | Concentration [µg/mL] | Additional Agent(s) | Cytotoxicity (72nd h) | Reference | ||
|---|---|---|---|---|---|---|---|
| Type | Cell Line | ||||||
| 1 | Mesoporous silica | Cervical cancer | HeLa | – | – | – | [53] |
| 2 | Polymeric | Breast cancer | MCF-7 | 3200 | – | ≈65% | [54] |
| 3 | Lipid | Lung carcinoma | H460 | – | – | in vivo | [55] |
| 4 | Peptide | Cervical cancer | HeLa | >1.3 | – | clonogenic potential block | [56] |
| 5 | Dendritic multidomain | Lung adenocarcinoma | A549 | 95 | – | ≈60% 1 | [57] |
| 6 | cytC-polymer | Cervical cancer | HeLa | 100 | – | ≈75–80% 2 | [58] |
| 7 | Hybrid iron oxide–gold | Liver cancer Hepatocellular carcinoma | HepG2 | 25 | – | ≈59% | [59,60] |
| 8 | Mesoporous silica | Hepatocellular carcinoma | HL-7702 HepG2 | – | – | ≈80% 1 | [61] |
| 9 | Mesoporous silica | Breast cancer | MCF-7 | – | doxorubicin | in vivo | [62] |
| 10 | Mesoporous silica | Cervical cancer | HeLa | 37.5 | Lactose | ≈55% | [63] |
| 11 | Galactosylated albumin | Liver cancer Hepatocellular carcinoma | HepG2 Hep3B | 10 | – | ≈35% 1 | [64] |
| 12 | CaCO3 | Breast cancer | MCF-7 | 50 | – | ≈35% 3 | [65] |
| 13 | cytC | Cervical cancer Lung adenocarcinoma | HeLa A549 | 120 | Oleic acid | ≈95% 2 ≈90% 2 | [66] |
| 14 | cytC | Cervical cancer | HeLa | 50 | Folate | 80% 2 | [67] |
| 15 | Cardiolipin | Ovarian cancer | A2780 | 1 | – | ≈70% | [68] |
| 16 | cytC | Lung adenocarcinoma | A549 | 164 | Hyaluronic acid | ≈90% 2 | [69] |
| 17 | Hybrid iron oxide–gold | Liver cancers | HepG2 Huh-7D SK-hep-1 | 12 | doxorubicinpaclitaxel oxaliplatin vinblastine vincristine | ≈60–75% 4 (maximal with vinblastine) | [70] |
| 18 | Mesoporous silica | Hepatocellular carcinoma | HepG2 | 150 | doxorubicin | ≈80% 1 | [71] |
| 19 | Ferritin | Acute promyelocytic leukemia (APL) | NB4 | 150 | – | – | [72] |
| 20 | Gold-ssDNA | Melanoma (skin) | B16F10 | 20 nM | – | ≈75–80% | [73] |
| 21 | Mesoporous silica NPs (degraded) | Ovarian cancer | SKOV3 | 40 | – | ≈80% 1 | [74] |
| 22 | Montmorillonite nanoplates | Colon cancer | CCL-227 | 3 | – | 97% | [75,76,77,78] |
| 23 | cytC | Cervical cancer Lewis lung carcinoma | HeLa LLC | 180 | Folate | ≈85% 2 ≈80% 2 | [79] |
| 24 | Polymeric nanomicelles | Liver cancer Hepatocellular carcinoma | HepG2 | 100 | – | ≈75% 1 | [80] |
| 25 | Polymeric nanomicelles | In vivo mice Lung adenocarcinoma | A549 | 50 | Paclitaxel | 95% | [81] |
| 26 | Gold–silica | Mice body tumor | HeLa | 50 | – | ≈80% | [82] |
| 27 | cytC | Lewis lung carcinoma | LLC | 300 | Folate | ≈90% | [83] |
| 28 | PEG, Hyaluronic acid nanogels | Lung adenocarcinoma | A549 | 100 | – | ≈70% 1 | [84] |
| 29 | Cationic dextrin | Cervical cancer | HeLa | 1500 | chloroquine | ≈88% 5 | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhivkov, A.M.; Hristova, S.H.; Popov, T.T. Anticancer Nanoparticle Carriers of the Proapoptotic Protein Cytochrome c. Pharmaceutics 2025, 17, 305. https://doi.org/10.3390/pharmaceutics17030305
Zhivkov AM, Hristova SH, Popov TT. Anticancer Nanoparticle Carriers of the Proapoptotic Protein Cytochrome c. Pharmaceutics. 2025; 17(3):305. https://doi.org/10.3390/pharmaceutics17030305
Chicago/Turabian StyleZhivkov, Alexandar M., Svetlana H. Hristova, and Trifon T. Popov. 2025. "Anticancer Nanoparticle Carriers of the Proapoptotic Protein Cytochrome c" Pharmaceutics 17, no. 3: 305. https://doi.org/10.3390/pharmaceutics17030305
APA StyleZhivkov, A. M., Hristova, S. H., & Popov, T. T. (2025). Anticancer Nanoparticle Carriers of the Proapoptotic Protein Cytochrome c. Pharmaceutics, 17(3), 305. https://doi.org/10.3390/pharmaceutics17030305