
Inhibition of Endothelial Cell Tube Formation by Anti-Vascular Endothelial Growth Factor/Anti-Angiopoietin-2 RNA Nanoparticles


 and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
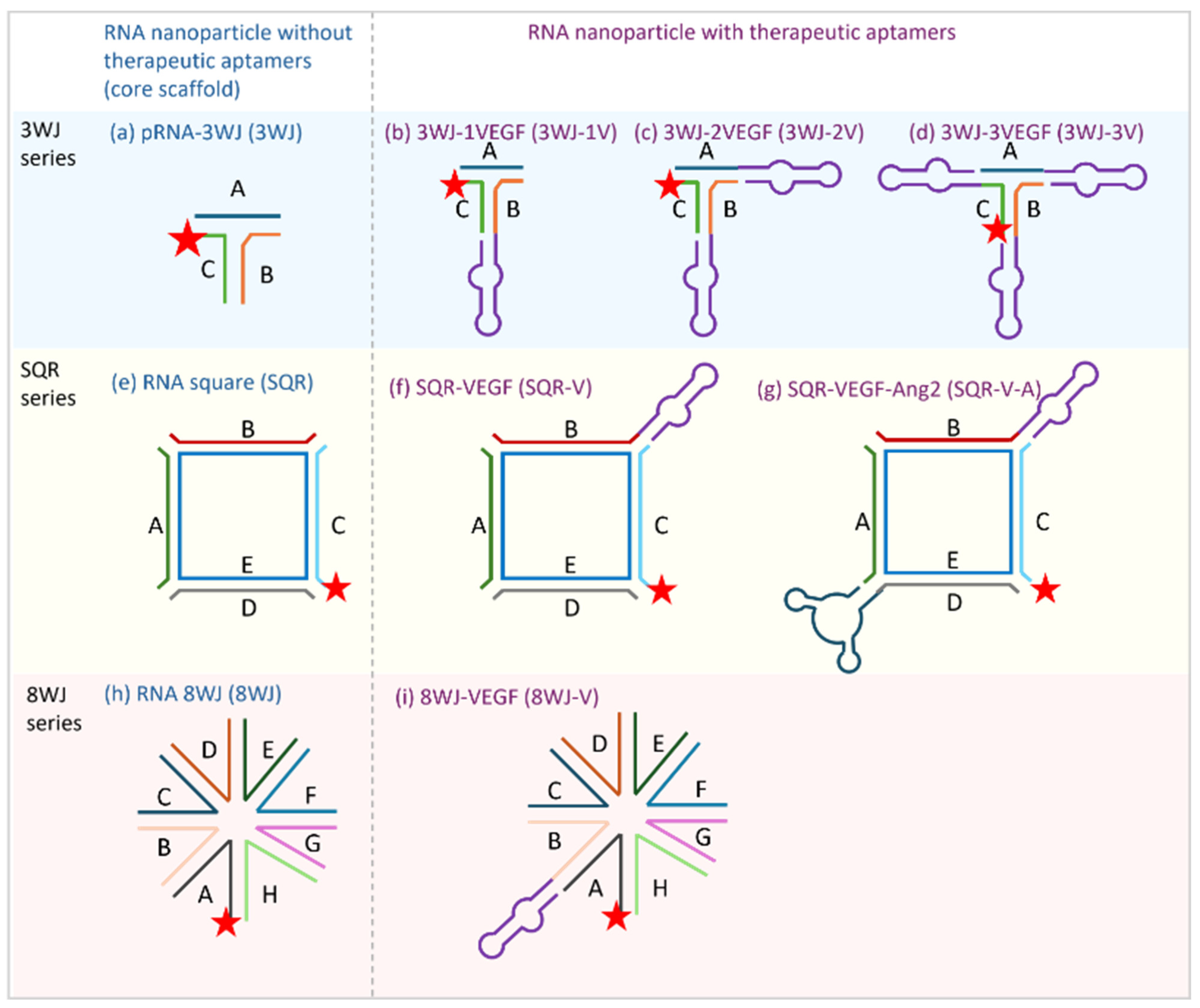
2.2. RNA Nanoparticles
2.3. Cell Culture
2.4. Three-Dimensional Spheroid Sprouting Assay
2.4.1. Spheroid Generation
2.4.2. Spheroid Sprouting Assay
2.5. Comparison Between 3D and 2D Assays
2.6. Statistical Analysis
3. Results
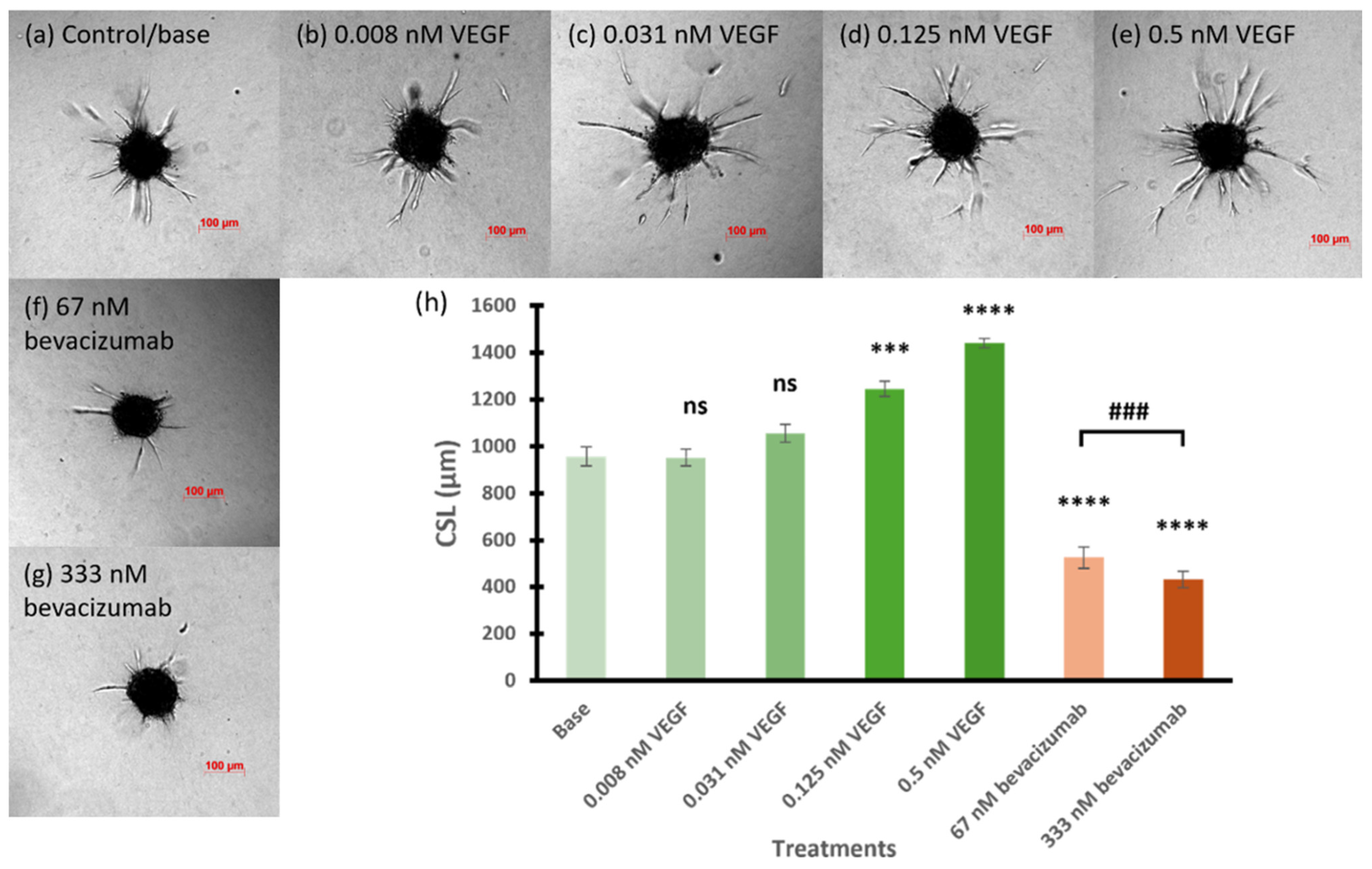
3.1. Feasibility of 3D Spheroid Sprouting Assay to Evaluate the Nanoparticles
3.2. Inhibitory Effect of RNA Nanoparticles
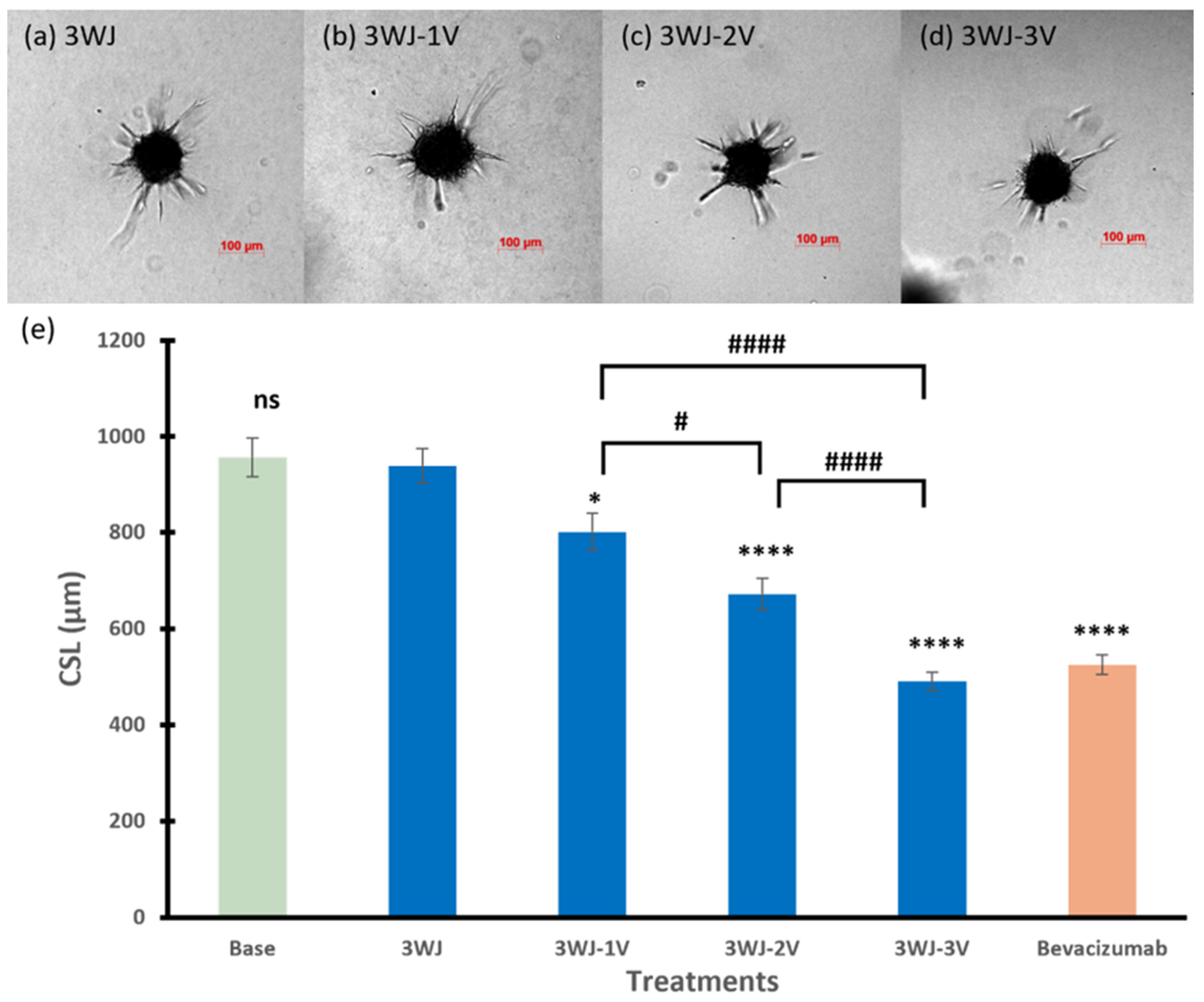
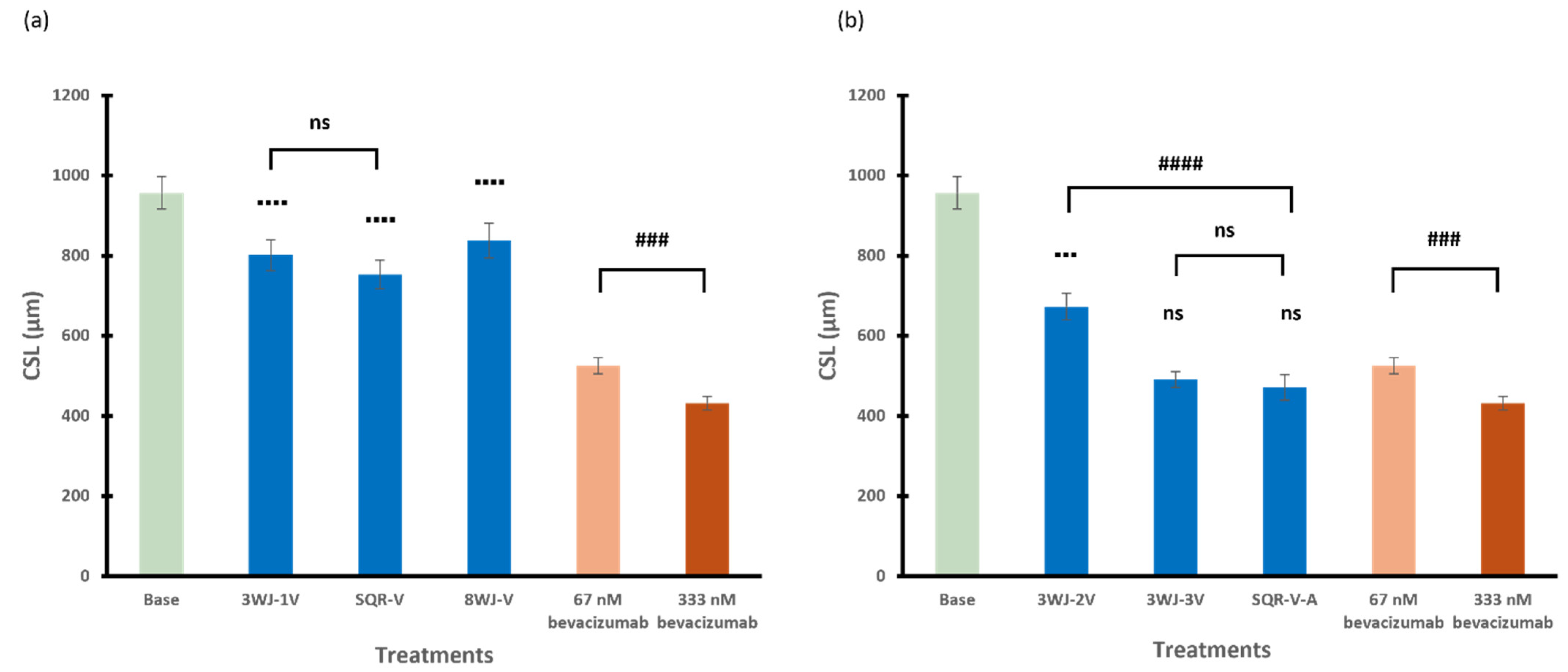
3.2.1. 3WJ Series
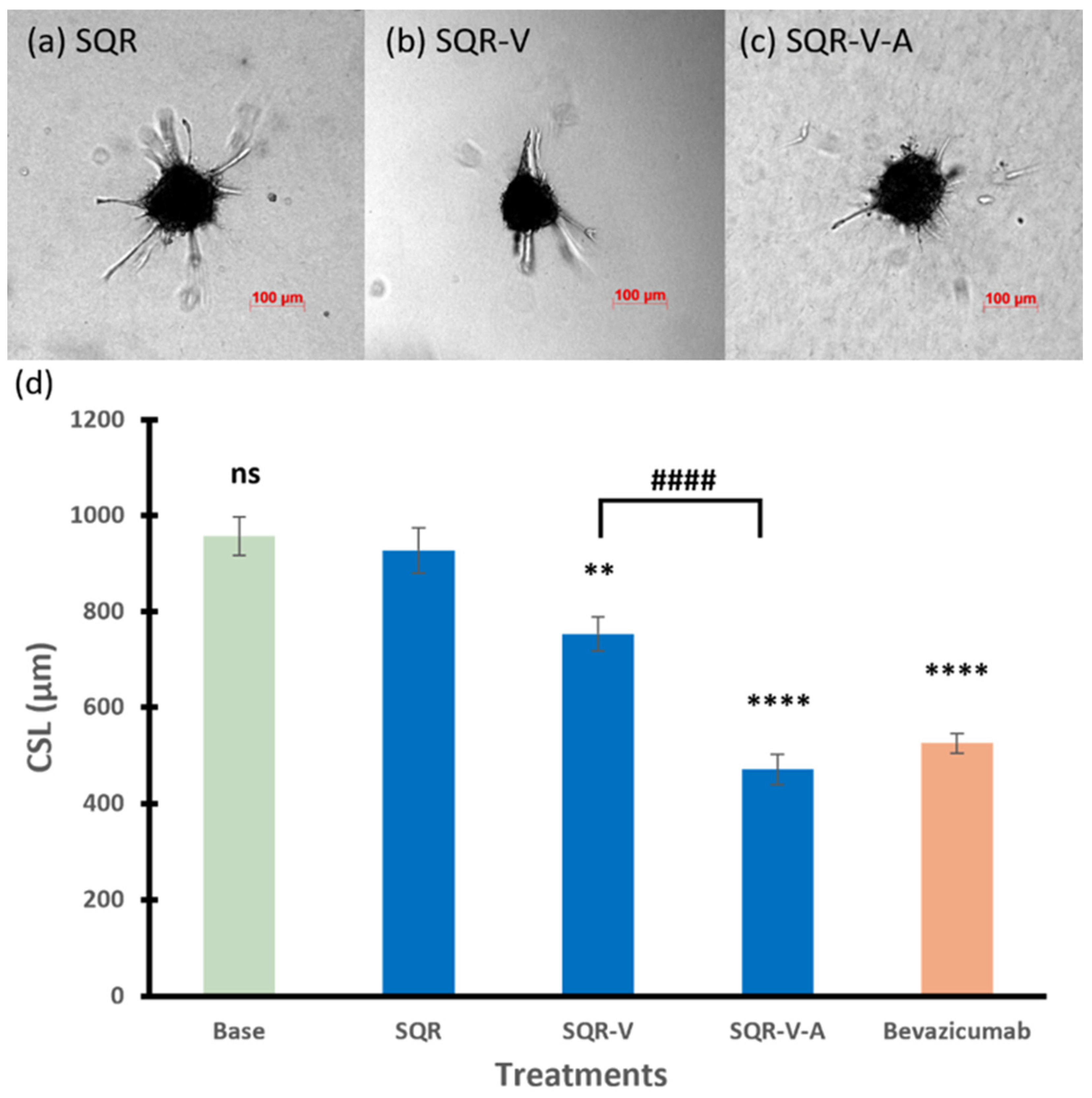
3.2.2. SQR Series
3.2.3. 8WJ Series
3.2.4. Among Series
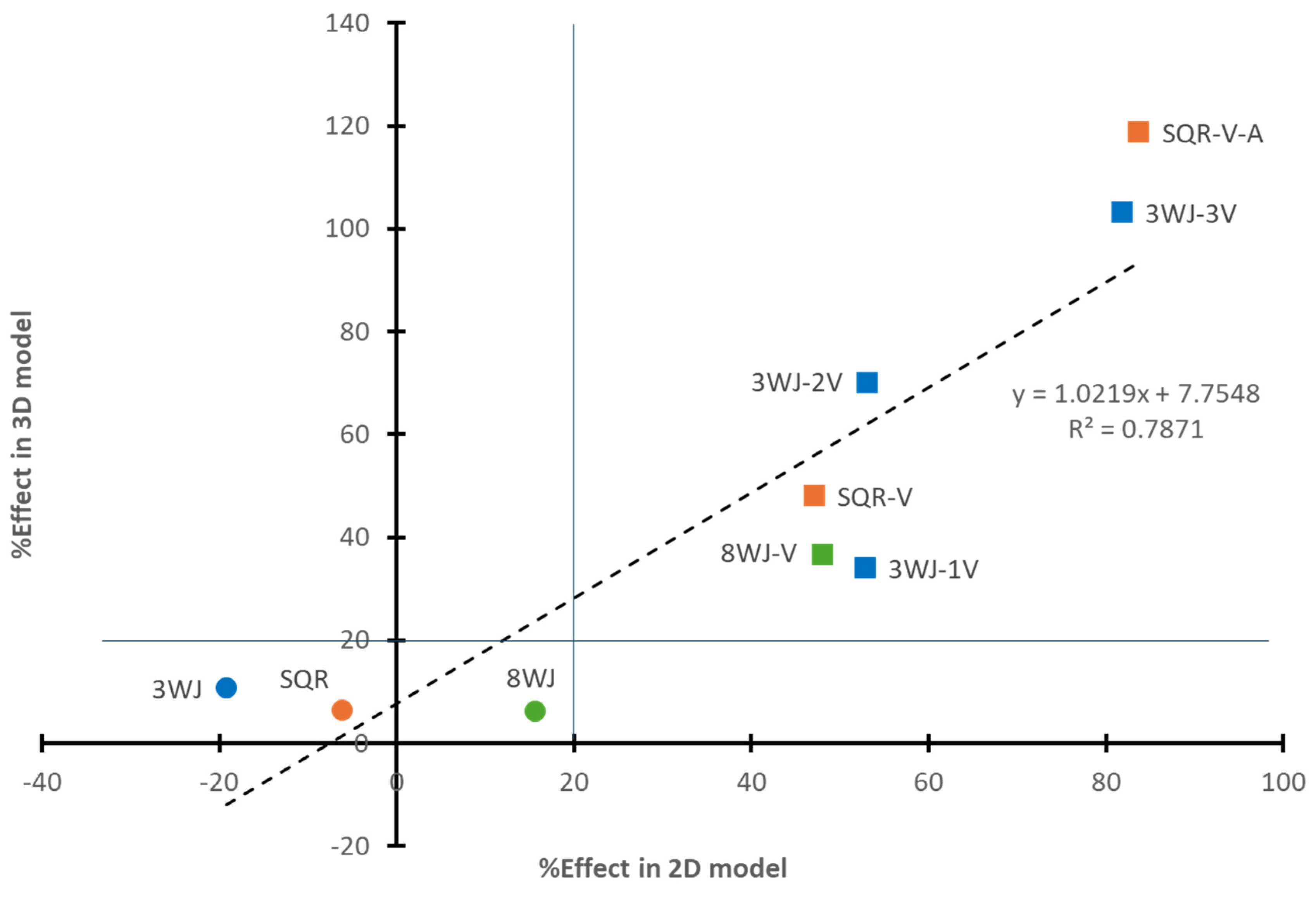
3.3. Similar Results Between 3D and 2D Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular mediators of angiogenesis. J. Burn Care Res. 2010, 31, 158–175. [Google Scholar] [CrossRef]
- White, R.R.; Shan, S.; Rusconi, C.P.; Shetty, G.; Dewhirst, M.W.; Kontos, C.D.; Sullenger, B.A. Inhibition of rat corneal angiogenesis by a nuclease-resistant RNA aptamer specific for angiopoietin-2. Proc. Natl. Acad. Sci. USA 2003, 100, 5028–5033. [Google Scholar] [CrossRef] [PubMed]
- Akwii, R.G.; Sajib, M.S.; Zahra, F.T.; Mikelis, C.M. Role of Angiopoietin-2 in vascular physiology and pathophysiology. Cells 2019, 8, 471. [Google Scholar] [CrossRef] [PubMed]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in signaling and disease: Beyond discovery and development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef]
- Cao, Y.; Langer, R.; Ferrara, N. Targeting angiogenesis in oncology, ophthalmology and beyond. Nat. Rev. Drug Discov. 2023, 22, 476–495. [Google Scholar] [CrossRef]
- Garcia, J.; Hurwitz, H.I.; Sandler, A.B.; Miles, D.; Coleman, R.L.; Deurloo, R.; Chinot, O.L. Bevacizumab (Avastin®) in cancer treatment: A review of 15 years of clinical experience and future outlook. Cancer Treat. Rev. 2020, 86, 102017. [Google Scholar] [CrossRef]
- Holekamp, N.M. Review of neovascular age-related macular degeneration treatment options. Am. J. Manag. Care 2019, 25, S172–S181. [Google Scholar]
- Kanwar, J.R.; Shankaranarayanan, J.S.; Gurudevan, S.; Kanwar, R.K. Aptamer-based therapeutics of the past, present and future: From the perspective of eye-related diseases. Drug Discov. Today 2014, 19, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Ng, E.W.; Shima, D.T.; Calias, P.; Cunningham, E.T., Jr.; Guyer, D.R.; Adamis, A.P. Pegaptanib, a targeted anti-VEGF aptamer for ocular vascular disease. Nat. Rev. Drug Discov. 2006, 5, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhu, L.; Wang, X.; Jin, H. RNA-based therapeutics: An overview and prospectus. Cell Death Dis. 2022, 13, 644. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.C.; Rossi, J.J. RNA-based therapeutics: Current progress and future prospects. Chem. Biol. 2012, 19, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Shi, Z.; Binzel, D.W.; Jin, K.; Li, X.; Guo, P.; Li, S.K. Posterior eye delivery of angiogenesis-inhibiting RNA nanoparticles via subconjunctival injection. Int. J. Pharm. 2024, 657, 124151. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Piao, X.; Li, H.; Guo, P. Methods for construction and characterization of simple or special multifunctional RNA nanoparticles based on the 3WJ of phi29 DNA packaging motor. Methods 2018, 143, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.X.; Bailey, S.; Bodley, J.W.; Anderson, D. Characterization of the small RNA of the bacteriophage phi 29 DNA packaging machine. Nucleic Acids Res. 1987, 15, 7081–7090. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lee, T.; Dziubla, T.; Pi, F.; Guo, S.; Xu, J.; Li, C.; Haque, F.; Liang, X.J.; Guo, P. RNA as a stable polymer to build controllable and defined nanostructures for material and biomedical applications. Nano Today 2015, 10, 631–655. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Haque, F.; Hallahan, B.; Reif, R.; Li, H. Uniqueness, advantages, challenges, solutions, and perspectives in therapeutics applying RNA nanotechnology. Nucleic Acid Ther. 2012, 22, 226–245. [Google Scholar] [CrossRef] [PubMed]
- Haque, F.; Shu, D.; Shu, Y.; Shlyakhtenko, L.S.; Rychahou, P.G.; Mark Evers, B.; Guo, P. Ultrastable synergistic tetravalent RNA nanoparticles for targeting to cancers. Nano Today 2012, 7, 245–257. [Google Scholar] [CrossRef]
- Wang, H.; Ellipilli, S.; Lee, W.-J.; Li, X.; Vieweger, M.; Ho, Y.-S.; Guo, P. Multivalent rubber-like RNA nanoparticles for targeted co-delivery of paclitaxel and MiRNA to silence the drug efflux transporter and liver cancer drug resistance. J. Control. Release 2021, 330, 173–184. [Google Scholar] [CrossRef]
- Yang, L.; Li, Z.; Binzel, D.W.; Guo, P.; Williams, T.M. Targeting oncogenic KRAS in non-small cell lung cancer with EGFR aptamer-conjugated multifunctional RNA nanoparticles. Mol. Ther. Nucleic Acids 2023, 33, 559–571. [Google Scholar] [CrossRef]
- Nowak-Sliwinska, P.; Alitalo, K.; Allen, E.; Anisimov, A.; Aplin, A.C.; Auerbach, R.; Augustin, H.G.; Bates, D.O.; van Beijnum, J.R.; Bender, R.H.F.; et al. Consensus guidelines for the use and interpretation of angiogenesis assays. Angiogenesis 2018, 21, 425–532. [Google Scholar] [CrossRef]
- Korff, T.; Augustin, H.G. Integration of endothelial cells in multicellular spheroids prevents apoptosis and induces differentiation. J. Cell Biol. 1998, 143, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Korff, T.; Augustin, H.G. Tensional forces in fibrillar extracellular matrices control directional capillary sprouting. J. Cell Sci. 1999, 112 Pt 19, 3249–3258. [Google Scholar] [CrossRef] [PubMed]
- Heiss, M.; Hellström, M.; Kalén, M.; May, T.; Weber, H.; Hecker, M.; Augustin, H.G.; Korff, T. Endothelial cell spheroids as a versatile tool to study angiogenesis in vitro. FASEB J. 2015, 29, 3076–3084. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Menger, M.D. Spheroids as vascularization units: From angiogenesis research to tissue engineering applications. Biotechnol. Adv. 2017, 35, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Nogués, C.; O’Brien, T. In vitro models for assessing therapeutic angiogenesis. Drug Discov. Today 2016, 21, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Shu, D.; Haque, F.; Guo, P. Fabrication of pRNA nanoparticles to deliver therapeutic RNAs and bioactive compounds into tumor cells. Nat. Protoc. 2013, 8, 1635–1659. [Google Scholar] [CrossRef]
- Tetzlaff, F.; Fischer, A. Human endothelial cell spheroid-based sprouting angiogenesis assay in collagen. Bio Protoc. 2018, 8, e2995. [Google Scholar] [CrossRef] [PubMed]
- Pfisterer, L.; Korff, T. Spheroid-based in vitro angiogenesis model. In Angiogenesis Protocols; Martin, S.G., Hewett, P.W., Eds.; Springer: New York, NY, USA, 2016; pp. 167–177. [Google Scholar]
- Liu, Y.; Tian, H.; Blobe, G.C.; Theuer, C.P.; Hurwitz, H.I.; Nixon, A.B. Effects of the combination of TRC105 and bevacizumab on endothelial cell biology. Investig. New Drugs 2014, 32, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Biel, N.M.; Siemann, D.W. Targeting the Angiopoietin-2/Tie-2 axis in conjunction with VEGF signal interference. Cancer Lett. 2016, 380, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Daly, C.; Eichten, A.; Castanaro, C.; Pasnikowski, E.; Adler, A.; Lalani, A.S.; Papadopoulos, N.; Kyle, A.H.; Minchinton, A.I.; Yancopoulos, G.D.; et al. Angiopoietin-2 functions as a Tie2 agonist in tumor models, where it limits the effects of VEGF inhibition. Cancer Res. 2013, 73, 108–118. [Google Scholar] [CrossRef]
- Brown, J.L.; Cao, Z.A.; Pinzon-Ortiz, M.; Kendrew, J.; Reimer, C.; Wen, S.; Zhou, J.Q.; Tabrizi, M.; Emery, S.; McDermott, B.; et al. A human monoclonal anti-ANG2 antibody leads to broad antitumor activity in combination with VEGF inhibitors and chemotherapy agents in preclinical models. Mol. Cancer Ther. 2010, 9, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Li, S.K.; Liu, H.; Liu, C.Y.; LaSance, K.; Haque, F.; Shu, D.; Guo, P. Ocular delivery of pRNA nanoparticles: Distribution and clearance after subconjunctival injection. Pharm. Res. 2014, 31, 1046–1058. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, S.K.; Charoenputtakun, P.; Liu, C.Y.; Jasinski, D.; Guo, P. RNA nanoparticle distribution and clearance in the eye after subconjunctival injection with and without thermosensitive hydrogels. J. Control. Release 2018, 270, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Abdelmawla, S.; Guo, S.; Zhang, L.; Pulukuri, S.M.; Patankar, P.; Conley, P.; Trebley, J.; Guo, P.; Li, Q.X. Pharmacological characterization of chemically synthesized monomeric phi29 pRNA nanoparticles for systemic delivery. Mol. Ther. 2011, 19, 1312–1322. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.; Shu, Y.; Haque, F.; Abdelmawla, S.; Guo, P. Thermodynamically stable RNA three-way junction for constructing multifunctional nanoparticles for delivery of therapeutics. Nat. Nanotechnol. 2011, 6, 658–667. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA Nanoparticles | Sequences |
|---|---|
| 3WJ series: | |
| 3WJ | A, 18 nt: 5′-uuG ccA uGu GuA uGu GGG-3′ B, 20 nt: 5′-ccc AcA uAc uuu Guu GAu cc-3′ C-AF647, 16 nt: 5′-GGA ucA Auc AuG GcA A-AF647-3′ |
| 3WJ-1V | A, 18 nt: 5′-uuG ccA uGu GuA uGu GGG-3′ B-VEGF, 47 nt: 5′-ccc AcA uAc uuu Guu GAu cc-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ C-AF647, 16 nt: 5′-GGA ucA Auc AuG GcA A-AF647-3′ |
| 3WJ-2V | A-VEGF, 45 nt: 5′-uuG ccA uGu GuA uGu GGG-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ B-VEGF, 47 nt: 5′-ccc AcA uAc uuu Guu GAu cc-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ C-AF647, 16 nt: 5′-GGA ucA Auc AuG GcA A-AF647-3′ |
| 3WJ-3V | A-VEGF, 45 nt: 5′-uuG ccA uGu GuA uGu GGG-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ B-VEGF, 47 nt: 5′-ccc AcA uAc uuu Guu GAu cc-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ AF647-C-VEGF, 43 nt: 5′-AF647-GGA ucA Auc AuG GcA A-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ |
| SQR series: | |
| SQR | A, 46 nt: 5′-GAG ccG ucA Auc AuG GcA AGu Guc cGc cAu Acu uuG uuG cAc GcA c-3′ B, 46 nt: 5′-GAG cGu GcA Auc AuG GcA AGc GcA ucG cAu Acu uuG uuG cGA ccA A-3′ C-AF647, 46 nt: 5′-GAG Guc GcA Auc AuG GcA AcG AuA GAG cAu Acu uuG uuG Gcu GGA G-AF647-3′ D, 46 nt: 5′-GAc cAG ccA Auc AuG GcA AuA uAc AcG cAu Acu uuG uuG AcG GcG G-3′ E, 88 nt: 5′-GGA cAc uuG ucA uGu GuA uGc GuG uAu Auu Guc AuG uGu AuG cuc uAu cGu uGu cAu GuG uAu GcG AuG cGc uuG ucA uGu GuA uGG c-3′ |
| SQR-V | A, 46 nt: 5′-GAG ccG ucA Auc AuG GcA AGu Guc cGc cAu Acu uuG uuG cAc GcA c-3′ B-VEGF, 73 nt: 5′-GAG cGu GcA Auc AuG GcA AGc GcA ucG cAu Acu uuG uuG cGA ccA A-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ C-AF647, 46 nt: 5′-GAG Guc GcA Auc AuG GcA AcG AuA GAG cAu Acu uuG uuG Gcu GGA G-AF647-3′ D, 46 nt: 5′-GAc cAG ccA Auc AuG GcA AuA uAc AcG cAu Acu uuG uuG AcG GcG G-3′ E, 88 nt: 5′-GGA cAc uuG ucA uGu GuA uGc GuG uAu Auu Guc AuG uGu AuG cuc uAu cGu uGu cAu GuG uAu GcG AuG cGc uuG ucA uGu GuA uGG c-3′ |
| SQR-V-A | A, 46 nt: 5′-GAG ccG ucA Auc AuG GcA AGu Guc cGc cAu Acu uuG uuG cAc GcA c-3′ B-VEGF, 73 nt: 5′-GAG cGu GcA Auc AuG GcA AGc GcA ucG cAu Acu uuG uuG cGA ccA A-cGG AAu cAG uGA AuG cuu AuA cAu ccG-3′ C-AF647, 46 nt: 5′-GAG Guc GcA Auc AuG GcA AcG AuA GAG cAu Acu uuG uuG Gcu GGA G–AF647-3′ D-Ang2, 87 nt: 5′-GAc cAG ccA Auc AuG GcA AuA uAc AcG cAu Acu uuG uuG AcG GcG G-GAGGAc GAu GcG GAc uAG ccu cAu cAG cuc AuG uGc ccc uc-3′ E, 88 nt: 5′-GGA cAc uuG ucA uGu GuA uGc GuG uAu Auu Guc AuG uGu AuG cuc uAu cGu uGu cAu GuG uAu GcG AuG cGc uuG ucA uGu GuA uGG c-3′ |
| 8WJ series: | |
| 8WJ | AF647-A, 42 nt: 5′-AF647-GAG uAu AuG uuA GGc cuG GGu GAG ucc uuG cGu cuu cuA ccG-3′ B, 42 nt: 5′-cGG uAG AAG AcG cAA GGA cuu Gcu AGu uGu GGu Acu Guu ccc-3′ C, 42 nt: 5′-GGG AAc AGu Acc AcA Acu AGu Guc ccG GGA uAG GGA cAu AcA-3′ D, 42 nt: 5′-uGu AuG ucc cuA ucc cGG GAu Gcu ccG cAu GAu GAA uAc AGc-3′ E, 42 nt: 5′-Gcu GuA uuc Auc AuG cGG AGu GGG cAu uGG GAu cGu AuG AGc-3′ F, 42 nt: 5′-Gcu cAu AcG Auc ccA AuG ccu GAA cAA AcA GAG cAA Gcc ucc-3′ G, 42 nt: 5′-GGA GGc uuG cuc uGu uuG uuu GcG cGA uuu ccG cGu uAc AcA-3′ H, 42 nt: 5′-uGu GuA AcG cGG AAA ucG cGu Gcc cAG Gcc uAA cAu AuA cuc-3′ |
| 8WJ-V | AF647-A, 42 nt: 5′-AF647-GAG uAu AuG uuA GGc cuG GGu GAG ucc uuG cGu cuu cuA ccG-3′ VEGF-B, 69 nt: 5′-cGG AAu cAG uGA AuG cuu AuA cAu ccG-cGG uAG AAG AcG cAA GGA cuu Gcu AGu uGu GGu Acu Guu ccc-3′ C, 42 nt: 5′-GGG AAc AGu Acc AcA Acu AGu Guc ccG GGA uAG GGA cAu AcA-3′ D, 42 nt: 5′-uGu AuG ucc cuA ucc cGG GAu Gcu ccG cAu GAu GAA uAc AGc-3′ E, 42 nt: 5′-Gcu GuA uuc Auc AuG cGG AGu GGG cAu uGG GAu cGu AuG AGc-3′ F, 42 nt: 5′-Gcu cAu AcG Auc ccA AuG ccu GAA cAA AcA GAG cAA Gcc ucc-3′ G, 42 nt: 5′-GGA GGc uuG cuc uGu uuG uuu GcG cGA uuu ccG cGu uAc AcA-3′ H, 42 nt: 5′-uGu GuA AcG cGG AAA ucG cGu Gcc cAG Gcc uAA cAu AuA cuc-3′ |
| Treatment | Sprout Number | Branching Point Number | Spheroid Size (Diameter, µm) |
|---|---|---|---|
| Control/base | 11.8 ± 0.5 | 2.3 ± 0.3 | 112.1 ± 1.0 |
| 0.5 nM VEGF | 15.9 ± 0.7 | 5.7 ± 0.6 | 114.0 ± 0.9 |
| 67 nM bevacizumab | 8.2 ± 0.3 | 0.47 ± 0.12 | 112.0 ± 1.0 |
| 333 nM bevacizumab | 7.7 ± 0.3 | 0.50 ± 0.12 | 111.4 ± 1.0 |
| 3WJ-3V | 8.5 ± 0.4 | 0.68 ± 0.13 | 113.0 ± 1.3 |
| SQR-V-A | 8.1 ± 0.5 | 0.51 ± 0.13 | 114.0 ± 1.0 |
| 8WJ-V | 11.6 ± 0.4 | 1.00 ± 0.18 | 113.2 ± 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, C.; Shi, Z.; Liu, C.-Y.; Binzel, D.W.; Jin, K.; Li, X.; Guo, P.; Li, S.K. Inhibition of Endothelial Cell Tube Formation by Anti-Vascular Endothelial Growth Factor/Anti-Angiopoietin-2 RNA Nanoparticles. Pharmaceutics 2025, 17, 55. https://doi.org/10.3390/pharmaceutics17010055
Zhong C, Shi Z, Liu C-Y, Binzel DW, Jin K, Li X, Guo P, Li SK. Inhibition of Endothelial Cell Tube Formation by Anti-Vascular Endothelial Growth Factor/Anti-Angiopoietin-2 RNA Nanoparticles. Pharmaceutics. 2025; 17(1):55. https://doi.org/10.3390/pharmaceutics17010055
Chicago/Turabian StyleZhong, Cheng, Zhanquan Shi, Chia-Yang Liu, Daniel W. Binzel, Kai Jin, Xin Li, Peixuan Guo, and S. Kevin Li. 2025. "Inhibition of Endothelial Cell Tube Formation by Anti-Vascular Endothelial Growth Factor/Anti-Angiopoietin-2 RNA Nanoparticles" Pharmaceutics 17, no. 1: 55. https://doi.org/10.3390/pharmaceutics17010055
APA StyleZhong, C., Shi, Z., Liu, C.-Y., Binzel, D. W., Jin, K., Li, X., Guo, P., & Li, S. K. (2025). Inhibition of Endothelial Cell Tube Formation by Anti-Vascular Endothelial Growth Factor/Anti-Angiopoietin-2 RNA Nanoparticles. Pharmaceutics, 17(1), 55. https://doi.org/10.3390/pharmaceutics17010055