
Chloroquine and Hydroxychloroquine, as Proteasome Inhibitors, Upregulate the Expression and Activity of Organic Anion Transporter 3

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Transfection
2.3. Measurement of 20S Proteasome Activity
2.4. Ubiquitination Assay
2.5. Measurement of Transport Activity
2.6. Kinetics of Estrone Sulfate Transport
2.7. Cell Surface Biotinylation
2.8. Degradation Assay
2.9. Gel Electrophoresis and Immunoblotting
2.10. Data Analysis
3. Results
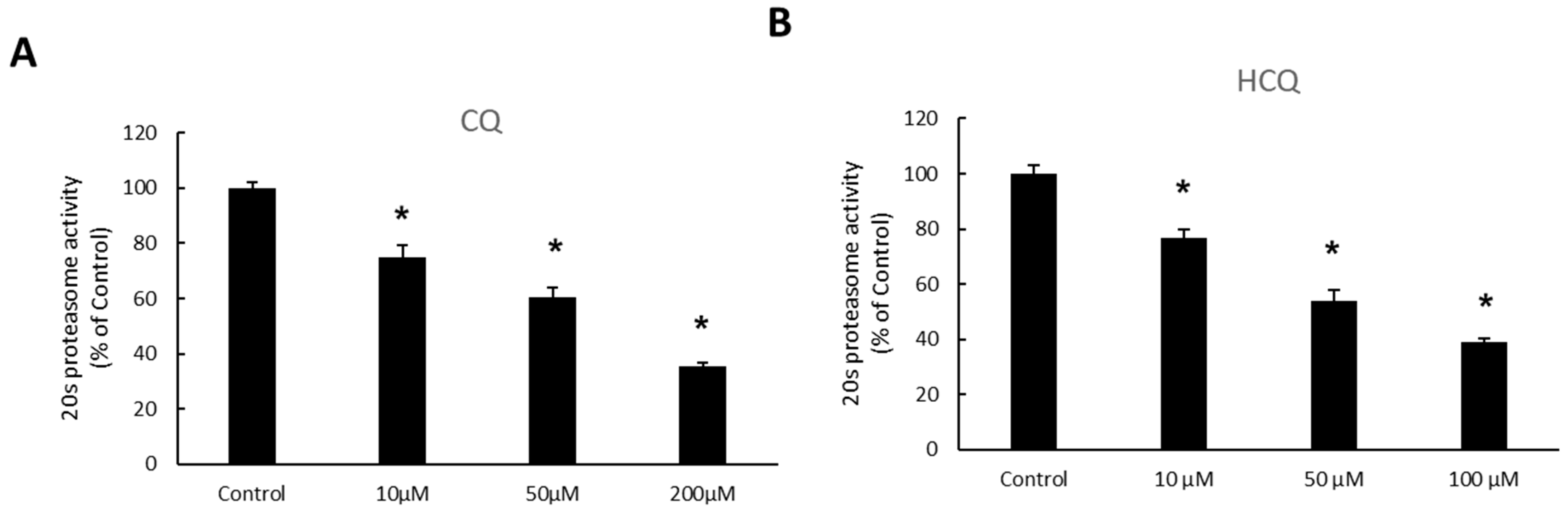
3.1. Effect of Chloroquine (CQ) and Hydroxychloroquine (HCQ) on 20S Proteasome Activity in OAT3-Expressing Cells
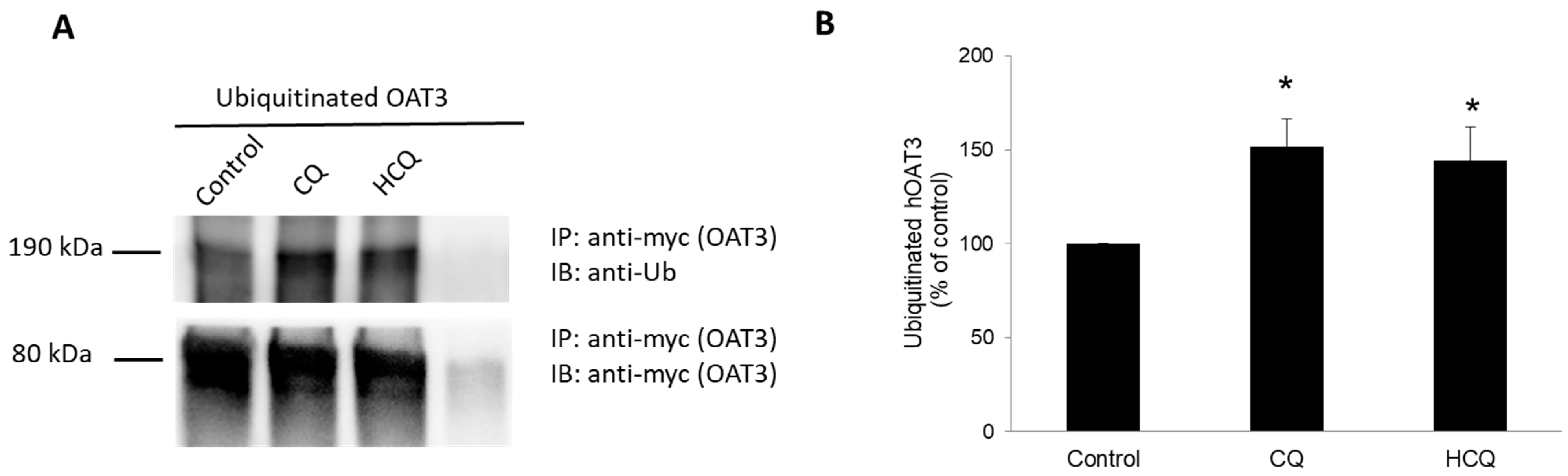
3.2. Effect of CQ and HCQ on OAT3 Ubiquitination
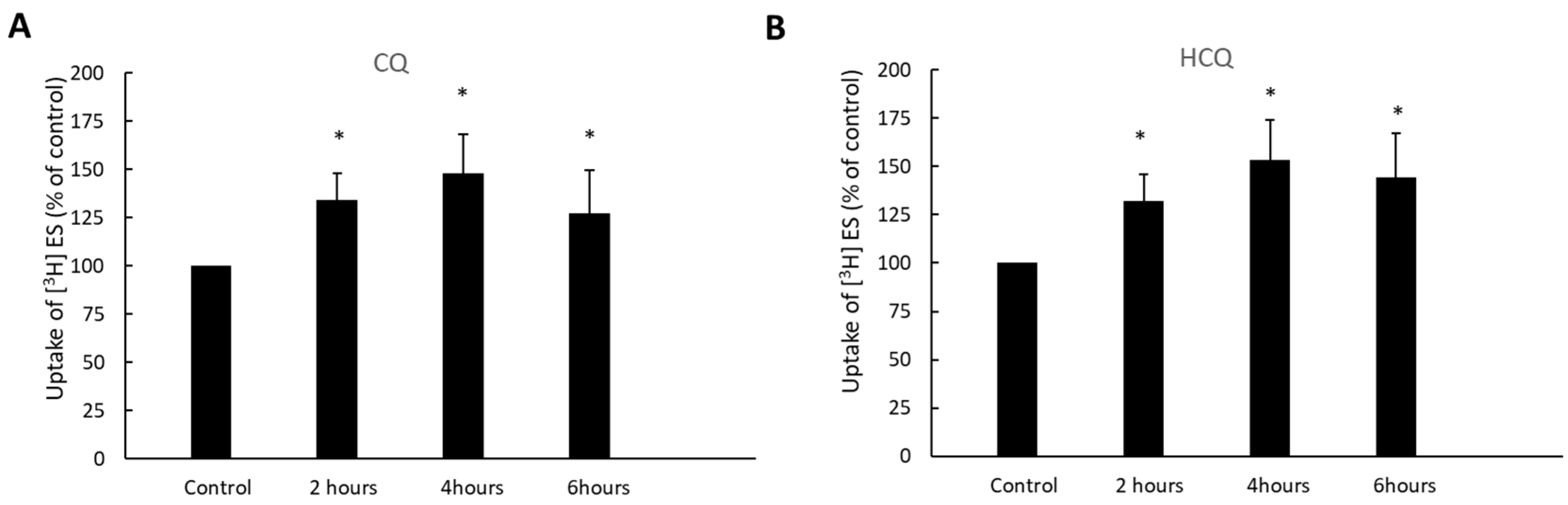
3.3. Time-Dependent Effect of CQ and HCQ on OAT3-Mediated Uptake of Estrone Sulfate
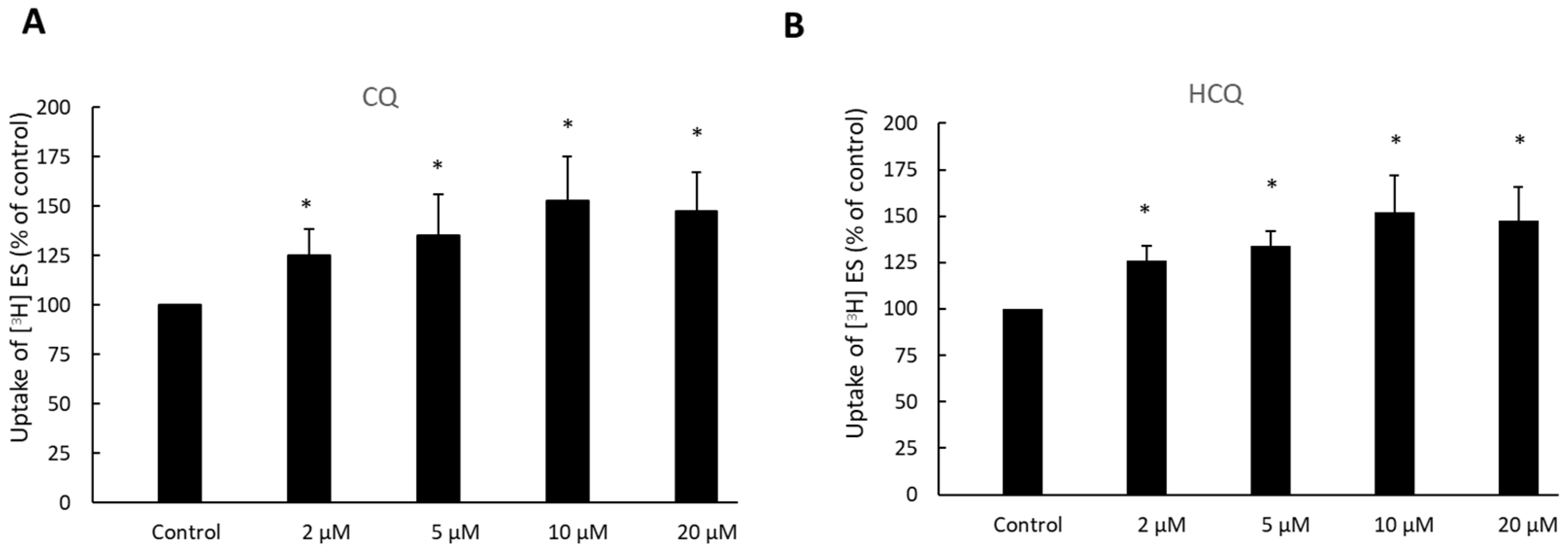
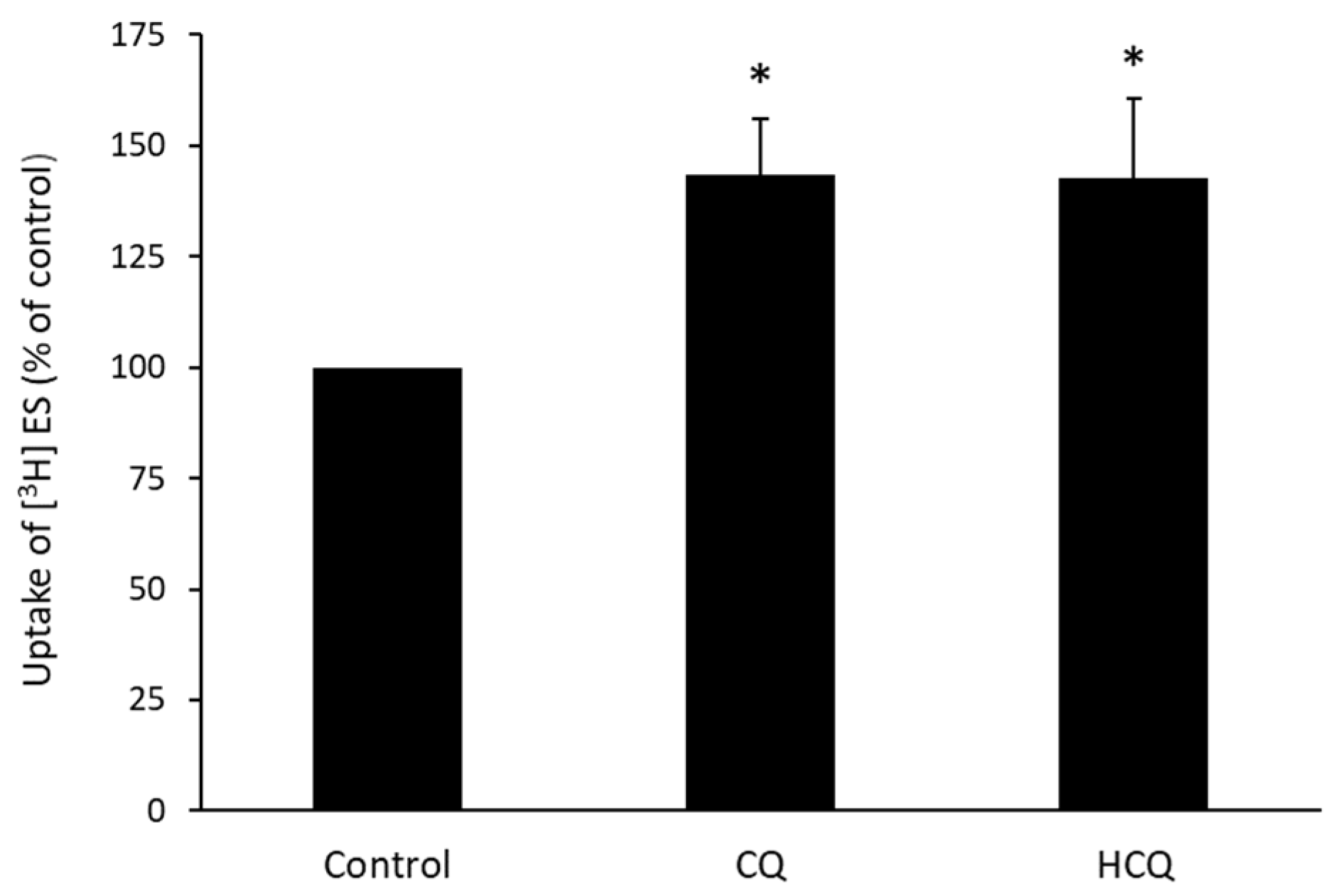
3.4. Dose-Dependent Effect of CQ and HCQ on OAT3-Mediated Uptake of Estrone Sulfate
3.5. Effect of CQ and HCQ on OAT3-Mediated Uptake of Estrone Sulfate in HEK293 Cells
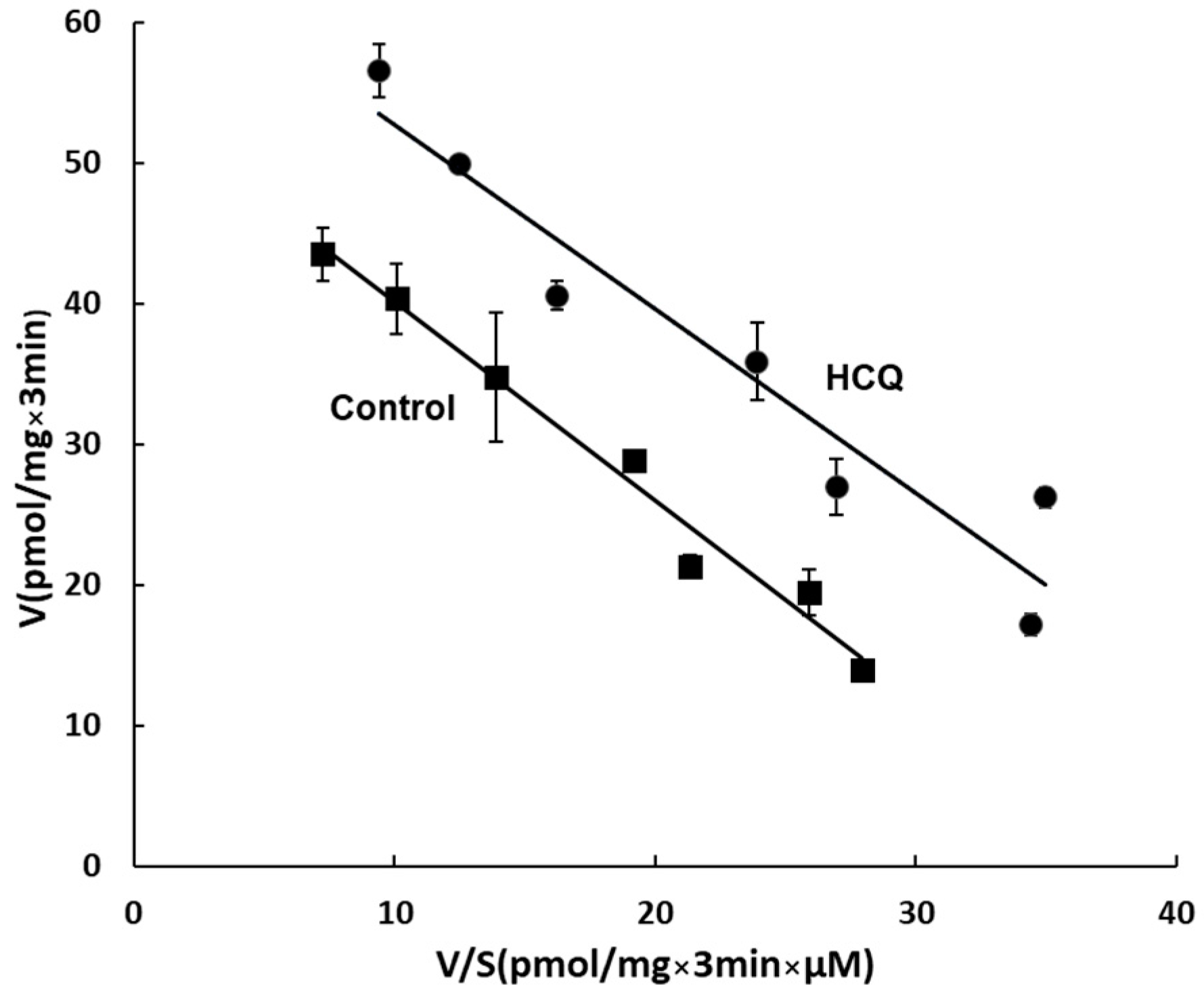
3.6. Kinetic Analysis of the Effect of HCQ on OAT3 Transport Activity
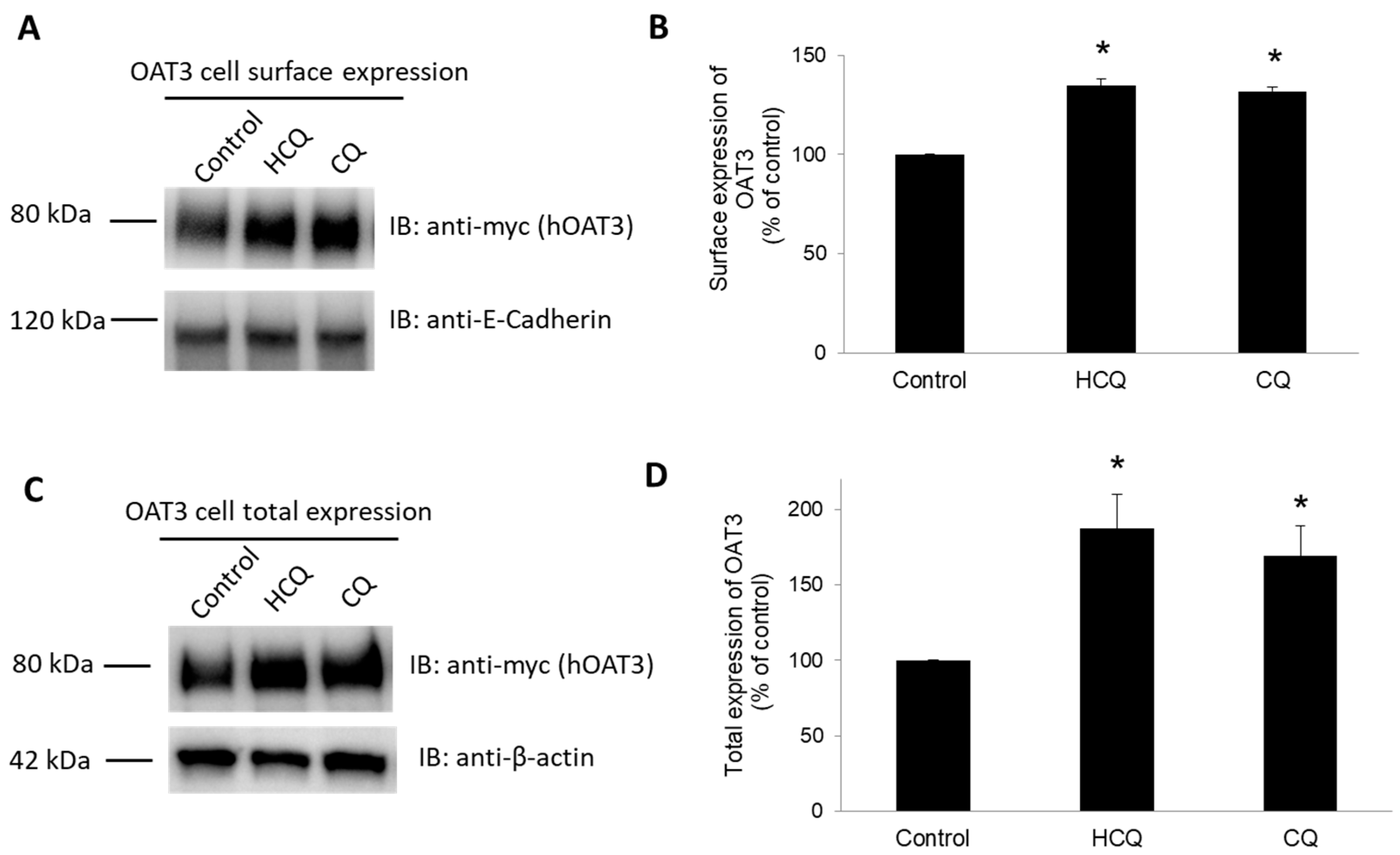
3.7. Effect of CQ and HCQ on OAT3 Expression
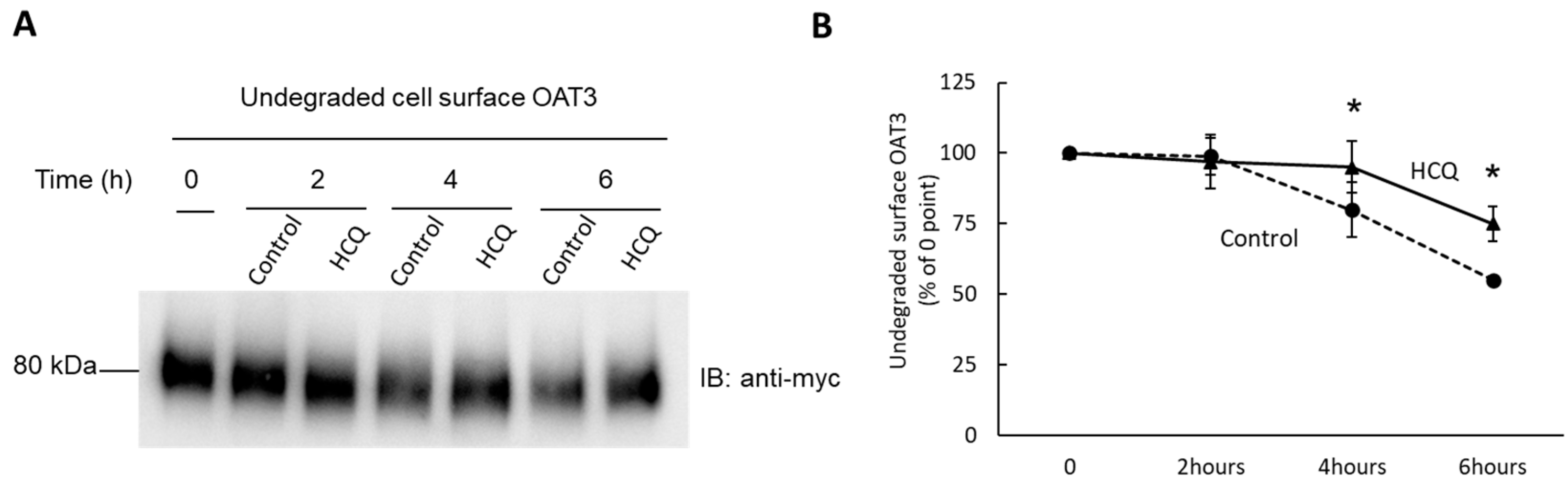
3.8. Effect of HCQ on OAT3 Stability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fan, Y.; Liang, Z.; Zhang, J.; You, G. Oral Proteasomal Inhibitors Ixazomib, Oprozomib, and Delanzomib Upregulate the Function of Organic Anion Transporter 3 (OAT3): Implications in OAT3-Mediated Drug-Drug Interactions. Pharmaceutics 2021, 13, 314. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.-Y.; Nigam, S.K. Toward a systems level understanding of organic anion and other multispecific drug transporters: A remote sensing and signaling hypothesis. Mol. Pharmacol. 2009, 76, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Kaminski, T.; Karbowska, M.; Pawlak, D. New insight into organic anion transporters from the perspective of potentially important interactions and drugs toxicity. J. Physiol. Pharmacol. 2018, 69. [Google Scholar] [CrossRef]
- Erdman, A.R.; Mangravite, L.M.; Urban, T.J.; Lagpacan, L.L.; Castro, R.A.; de la Cruz, M.; Chan, W.; Huang, C.C.; Johns, S.J.; Kawamoto, M. The human organic anion transporter 3 (OAT3; SLC22A8): Genetic variation and functional genomics. Am. J. Physiol.-Ren. Physiol. 2006, 290, F905–F912. [Google Scholar] [CrossRef]
- Wu, W.; Jamshidi, N.; Eraly, S.A.; Liu, H.C.; Bush, K.T.; Palsson, B.O.; Nigam, S.K. Multispecific drug transporter Slc22a8 (Oat3) regulates multiple metabolic and signaling pathways. Drug Metab. Dispos. 2013, 41, 1825–1834. [Google Scholar] [CrossRef]
- Fan, Y.; You, G. Proteasome Inhibitors Bortezomib and Carfilzomib Stimulate the Transport Activity of Human Organic Anion Transporter 1. Mol Pharm. 2020, 97, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Dantzler, W.H.; Wright, S.H. The molecular and cellular physiology of basolateral organic anion transport in mammalian renal tubules. Biochim. Biophys. Acta (BBA)-Biomembr. 2003, 1618, 185–193. [Google Scholar] [CrossRef]
- Wang, L.; Sweet, D.H. Renal organic anion transporters (SLC22 family): Expression, regulation, roles in toxicity, and impact on injury and disease. AAPS J. 2013, 15, 53–69. [Google Scholar] [CrossRef]
- Xu, D.; Wang, H.; You, G. An Essential Role of Nedd4-2 in the Ubiquitination, Expression, and Function of Organic Anion Transporter-3. Mol. Pharm. 2016, 13, 621–630. [Google Scholar] [CrossRef]
- Zhang, Q.; Suh, W.; Pan, Z.; You, G. Short-term and long-term effects of protein kinase C on the trafficking and stability of human organic anion transporter 3. Int. J. Biochem. Mol. Biol. 2012, 3, 242. [Google Scholar]
- Fan, Y.; Wang, H.; Yu, Z.; Liang, Z.; Li, Y.; You, G. Inhibition of proteasome, but not lysosome, upregulates organic anion transporter 3 in vitro and in vivo. Biochem. Pharmacol. 2023, 208, 115387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, S.; Patterson, C.; You, G. Lysine 48-Linked Polyubiquitination of Organic Anion Transporter-1 Is Essential for Its Protein Kinase C–Regulated Endocytosis. Mol. Pharmacol. 2013, 83, 217–224. [Google Scholar] [CrossRef]
- Sprangers, R.; Li, X.; Mao, X.; Rubinstein, J.L.; Schimmer, A.D.; Kay, L.E. TROSY-Based NMR Evidence for a Novel Class of 20S Proteasome Inhibitors. Biochemistry 2008, 47, 6727–6734. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, R.; Xu, M.; Wang, X.; Zhang, H.; Hu, H.; Li, Y.; Hu, Z.; Zhong, W.; Wang, M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020, 6, 16. [Google Scholar] [CrossRef]
- McChesney, E.W. Animal toxicity and pharmacokinetics of hydroxychloroquine sulfate. Am. J. Med. 1983, 75, 11–18. [Google Scholar] [CrossRef]
- Shippey, E.A.; Wagler, V.D.; Collamer, A.N. Hydroxychloroquine: An old drug with new relevance. Clevel. Clin. J. Med. 2018, 85, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Stokkermans, T.J.; Goyal, A.; Bansal, P.; Trichonas, G. Chloroquine and Hydroxychloroquine Toxicity; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Chen, Y.; Traore, Y.L.; Li, A.; Fowke, K.R.; Ho, E.A. Development of polyether urethane intravaginal rings for the sustained delivery of hydroxychloroquine. Drug Des. Dev. Ther. 2014, 8, 1801–1815. [Google Scholar]
- Schrezenmeier, E.; Dörner, T. Mechanisms of action of hydroxychloroquine and chloroquine: Implications for rheumatology. Nat. Rev. Rheumatol. 2020, 16, 155–166. [Google Scholar] [CrossRef]
- “Aralen Phosphate”. The American Society of Health-System Pharmacists. Archived from the Original on 8 December 2015. Retrieved 2 December 2015. Available online: https://www.drugs.com/monograph/chloroquine.html (accessed on 29 November 2021).
- Fox, R.I. Mechanism of action of hydroxychloroquine as an antirheumatic drug. Semin. Arthritis Rheum. 1993, 23, 82–91. [Google Scholar] [CrossRef]
- Rangwala, R.; Chang, Y.C.; Hu, J.; Algazy, K.M.; Evans, T.L.; Fecher, L.A.; Schuchter, L.M.; Torigian, D.A.; Panosian, J.T.; Troxel, A.B. Combined MTOR and autophagy inhibition: Phase I trial of hydroxychloroquine and temsirolimus in patients with advanced solid tumors and melanoma. Autophagy 2014, 10, 1391–1402. [Google Scholar] [CrossRef]
- Manic, G.; Obrist, F.; Kroemer, G.; Vitale, I.; Galluzzi, L. Chloroquine and hydroxychloroquine for cancer therapy. Mol. Cell. Oncol. 2014, 1, e29911. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.-T.; Yu, X.-X.; Yan, L.-J.; Xiao, H.-T. Research progress of hydroxychloroquine and autophagy inhibitors on cancer. Cancer Chemother. Pharmacol. 2017, 79, 287–294. [Google Scholar] [CrossRef]
- Maycotte, P.; Aryal, S.; Cummings, C.T.; Thorburn, J.; Morgan, M.J.; Thorburn, A. Chloroquine sensitizes breast cancer cells to chemotherapy independent of autophagy. Autophagy 2012, 8, 200–212. [Google Scholar] [CrossRef]
- Boone, B.A.; Bahary, N.; Zureikat, A.H.; Moser, A.J.; Normolle, D.P.; Wu, W.-C.; Singhi, A.D.; Bao, P.; Bartlett, D.L.; Liotta, L.A. Safety and biologic response of pre-operative autophagy inhibition in combination with gemcitabine in patients with pancreatic adenocarcinoma. Ann. Surg. Oncol. 2015, 22, 4402–4410. [Google Scholar] [CrossRef] [PubMed]
- Montanari, F.; Lu, M.; Marcus, S.; Saran, A.; Malankar, A.; Mazumder, A. A phase II trial of chloroquine in combination with bortezomib and cyclophosphamide in patients with relapsed and refractory multiple myeloma. Blood 2014, 124, 5775. [Google Scholar] [CrossRef]
- Goldberg, S.B.; Supko, J.G.; Neal, J.W.; Muzikansky, A.; Digumarthy, S.; Fidias, P.; Temel, J.S.; Heist, R.S.; Shaw, A.T.; McCarthy, P.O. A phase I study of erlotinib and hydroxychloroquine in advanced non–small-cell lung cancer. J. Thorac. Oncol. 2012, 7, 1602–1608. [Google Scholar] [CrossRef]
- Briceño, E.; Calderon, A.; Sotelo, J. Institutional experience with chloroquine as an adjuvant to the therapy for glioblastoma multiforme. Surg. Neurol. 2007, 67, 388–391. [Google Scholar] [CrossRef]
- Ducharme, J.; Farinotti, R. Clinical pharmacokinetics and metabolism of chloroquine. Focus on recent advancements. Clin Pharm. 1996, 31, 257–274. [Google Scholar] [CrossRef] [PubMed]
- FDA Approved Drug Products: Hydroxychloroquine Oral Tablets. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2019/009768Orig1s051lbl.pdf (accessed on 1 June 2018).
- Pari, L.; Murugan, P. Tetrahydrocurcumin: Effect on chloroquine-mediated oxidative damage in rat kidney. Basic Clin. Pharmacol. Toxicol. 2006, 99, 329–334. [Google Scholar] [CrossRef]
- Mũller-Hõcker, J.; Schmid, H.; Weiss, M.; Dendorfer, U.; Braun, G.S. Chloroquine-induced phospholipidosis of the kidney mimicking fabry's disease: Case report and review of the literature. Hum. Pathol. 2003, 34, 285–289. [Google Scholar] [CrossRef]
- Verbaanderd, C.; Maes, H.; Schaaf, M.B.; Sukhatme, V.P.; Pantziarka, P.; Sukhatme, V.; Agostinis, P.; Bouche, G. Repurposing Drugs in Oncology (ReDO)-chloroquine and hydroxychloroquine as anti-cancer agents. Ecancermedicalscience 2017, 11, 781. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wang, H.; Gardner, C.; Pan, Z.; Zhang, P.L.; Zhang, J.; You, G. The role of Nedd4-1 WW domains in binding and regulating human organic anion transporter 1. Am. J. Physiol.-Ren. Physiol. 2016, 311, F320–F329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hong, M.; Duan, P.; Pan, Z.; Ma, J.; You, G. Organic anion transporter OAT1 undergoes constitutive and protein kinase C-regulated trafficking through a dynamin-and clathrin-dependent pathway. J. Biol. Chem. 2008, 283, 32570–32579. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, J.; Zhang, Q.; Fan, Y.; Liu, C.; You, G. PKC/Nedd4-2 signaling pathway regulates the cell surface expression of drug transporter hOAT1. Drug Metab. Dispos. 2017, 45, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Hofstee, B. Non-inverted versus inverted plots in enzyme kinetics. Nature 1959, 184, 1296–1298. [Google Scholar] [CrossRef]
- You, G. Structure, function, and regulation of renal organic anion transporters. Med. Res. Rev. 2002, 22, 602–616. [Google Scholar] [CrossRef]
- Nigam, S.K.; Bush, K.T.; Martovetsky, G.; Ahn, S.-Y.; Liu, H.C.; Richard, E.; Bhatnagar, V.; Wu, W. The organic anion transporter (OAT) family: A systems biology perspective. Physiol. Rev. 2015, 95, 83–123. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, J.; You, G. Activation of protein kinase A stimulates SUMOylation, expression, and transport activity of organic anion transporter 3. AAPS J. 2019, 21, 1–11. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, H.; Fan, Y.; Yu, Z.; You, G. Regulation of organic anion transporters: Role in physiology, pathophysiology, and drug elimination. Pharmacol. Ther. 2021, 217, 107647. [Google Scholar] [CrossRef]
- Hsueh, C.-H.; Yoshida, K.; Zhao, P.; Meyer, T.W.; Zhang, L.; Huang, S.-M.; Giacomini, K.M. Identification and quantitative assessment of uremic solutes as inhibitors of renal organic anion transporters, OAT1 and OAT3. Mol. Pharm. 2016, 13, 3130–3140. [Google Scholar] [CrossRef]
- Nigam, S.K. What do drug transporters really do? Nat. Rev. Drug Discov. 2015, 14, 29–44. [Google Scholar] [CrossRef] [PubMed]
- The International Transporter Consortium. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.; Fromm, M.F. Transporter-mediated drug–drug interactions. Pharmacogenomics 2011, 12, 1017–1037. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, C.; Meng, Q.; Huo, X.; Sun, H.; Peng, J.; Ma, X.; Sun, P.; Liu, K. MDR1 and OAT1/OAT3 mediate the drug-drug interaction between puerarin and methotrexate. Pharm. Res. 2014, 31, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, X.; Bi, Y.; Yu, H.; Wei, J.; Zhang, Y.; Han, L.; Zhang, Y. Potent inhibitors of organic anion transporters 1 and 3 from natural compounds and their protective effect on aristolochic acid nephropathy. Toxicol. Sci. 2020, 175, 279–291. [Google Scholar] [CrossRef]
- Zhu, Y.; Huo, X.; Wang, C.; Meng, Q.; Liu, Z.; Sun, H.; Tan, A.; Ma, X.; Peng, J.; Liu, K. Organic anion transporters also mediate the drug–drug interaction between imipenem and cilastatin. Asian J. Pharm. Sci. 2020, 15, 252–263. [Google Scholar] [CrossRef]
- Titus, E.O. Recent developments in the understanding of the pharmacokinetics and mechanism of action of chloroquine. Ther. Drug Monit. 1989, 11, 369–379. [Google Scholar] [CrossRef]
- Lim, H.S.; Im, J.S.; Cho, J.Y.; Bae, K.S.; Klein, T.A.; Yeom, J.S.; Kim, T.S.; Choi, J.S.; Jang, I.J.; Park, J.W. Pharmacokinetics of hydroxychloroquine and its clinical implications in chemoprophylaxis against malaria caused by Plasmodium vivax. Antimicrob Agents Chemother 2009, 53, 1468–1475. [Google Scholar] [CrossRef]
- Carmichael, S.J.; Charles, B.; Tett, S.E. Population pharmacokinetics of hydroxychloroquine in patients with rheumatoid arthritis. Ther. Drug Monit. 2003, 25, 671–681. [Google Scholar] [CrossRef]
- Furst, D.E. Pharmacokinetics of hydroxychloroquine and chloroquine during treatment of rheumatic diseases. Lupus 1996, 5, 11–15. [Google Scholar] [CrossRef]
- Bhardwaj, R.K.; Herrera-Ruiz, D.; Eltoukhy, N.; Saad, M.; Knipp, G.T. The functional evaluation of human peptide/histidine transporter 1 (hPHT1) in transiently transfected COS-7 cells. Eur. J. Pharm. Sci. 2006, 27, 533–542. [Google Scholar] [CrossRef]
- Xue, X.; Gong, L.-K.; Maeda, K.; Luan, Y.; Qi, X.-M.; Sugiyama, Y.; Ren, J. Critical role of organic anion transporters 1 and 3 in kidney accumulation and toxicity of aristolochic acid I. Mol. Pharm. 2011, 8, 2183–2192. [Google Scholar] [CrossRef] [PubMed]
- Toh, M.F.; Suh, W.; Wang, H.; Zhou, P.; Hu, L.; You, G. Inhibitory effects of chemotherapeutics on human organic anion transporter hOAT4. Int. J. Biochem. Mol. Biol. 2016, 7, 11. [Google Scholar] [PubMed]
- Yu, Z.; Zhang, J.; Liang, Z.; Wu, J.; Liu, K.; You, G. Pancreatic Hormone Insulin Modulates Organic Anion Transporter 1 in the Kidney: Regulation via Remote Sensing and Signaling Network. AAPS J. 2023, 25, 13. [Google Scholar] [CrossRef] [PubMed]
- Rödiger, M.; Zhang, X.; Ugele, B.; Gersdorff, N.; Wright, S.H.; Burckhardt, G.; Bahn, A. Organic anion transporter 3 (OAT3) and renal transport of the metal chelator 2, 3-dimercapto-1-propanesulfonic acid (DMPS). Can. J. Physiol. Pharmacol. 2010, 88, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.P.; Jackson, K.M.; Gustafson, D.L. Hydroxychloroquine: A Physiologically-Based Pharmacokinetic Model in the Context of Cancer-Related Autophagy Modulation. J. Pharm. Exp. 2018, 365, 447–459. [Google Scholar] [CrossRef]
- Mauthe, M.; Orhon, I.; Rocchi, C.; Zhou, X.; Luhr, M.; Hijlkema, K.-J.; Coppes, R.P.; Engedal, N.; Mari, M.; Reggiori, F. Chloroquine inhibits autophagic flux by decreasing autophagosome-lysosome fusion. Autophagy 2018, 14, 1435–1455. [Google Scholar] [CrossRef]
- Gonzalez-Noriega, A.; Grubb, J.H.; Talkad, V.; Sly, W.S. Chloroquine inhibits lysosomal enzyme pinocytosis and enhances lysosomal enzyme secretion by impairing receptor recycling. J. Cell Biol. 1980, 85, 839–852. [Google Scholar] [CrossRef]
- Abraham, R.; Hendy, R.; Grasso, P. Formation of myeloid bodies in rat liver lysosomes after chloroquine administration. Exp. Mol. Pathol. 1968, 9, 212–229. [Google Scholar] [CrossRef]
- Lie, S.O.; Schofield, B. Inactivation of lysosomal function in normal cultured human fibroblasts by chloroquine. Biochem. Pharmacol. 1973, 22, 3109–3114. [Google Scholar] [CrossRef]
- Boya, P.; Gonzalez-Polo, R.-A.; Poncet, D.; Andreau, K.; Vieira, H.L.; Roumier, T.; Perfettini, J.-L.; Kroemer, G. Mitochondrial membrane permeabilization is a critical step of lysosome-initiated apoptosis induced by hydroxychloroquine. Oncogene 2003, 22, 3927–3936. [Google Scholar] [CrossRef] [PubMed]
- Sundelin, S.P.; Terman, A. Different effects of chloroquine and hydroxychloroquine on lysosomal function in cultured retinal pigment epithelial cells. Apmis 2002, 110, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.; Gonzalez, E.; Visekruna, A.; Kühl, A.A.; Loddenkemper, C.; Mollenkopf, H.; Kaufmann, S.H.E.; Steinhoff, U.; Joeris, T. Targeting the proteasome: Partial inhibition of the proteasome by bortezomib or deletion of the immunosubunit LMP7 attenuates experimental colitis. Gut 2010, 59, 896–906. [Google Scholar] [CrossRef]
- Coëffier, M.; Gloro, R.; Boukhettala, N.; Aziz, M.; Lecleire, S.; Vandaele, N.; Antonietti, M.; Savoye, G.; Bôle-Feysot, C.; Déchelotte, P. Increased proteasome-mediated degradation of occludin in irritable bowel syndrome. Off. J. Am. Coll. Gastroenterol.|ACG 2010, 105, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Gandolfi, S.; Laubach, J.P.; Hideshima, T.; Chauhan, D.; Anderson, K.C.; Richardson, P.G. The proteasome and proteasome inhibitors in multiple myeloma. Cancer Metastasis Rev. 2017, 36, 561–584. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Z.; You, G. Chloroquine and Hydroxychloroquine, as Proteasome Inhibitors, Upregulate the Expression and Activity of Organic Anion Transporter 3. Pharmaceutics 2023, 15, 1725. https://doi.org/10.3390/pharmaceutics15061725
Liang Z, You G. Chloroquine and Hydroxychloroquine, as Proteasome Inhibitors, Upregulate the Expression and Activity of Organic Anion Transporter 3. Pharmaceutics. 2023; 15(6):1725. https://doi.org/10.3390/pharmaceutics15061725
Chicago/Turabian StyleLiang, Zhengxuan, and Guofeng You. 2023. "Chloroquine and Hydroxychloroquine, as Proteasome Inhibitors, Upregulate the Expression and Activity of Organic Anion Transporter 3" Pharmaceutics 15, no. 6: 1725. https://doi.org/10.3390/pharmaceutics15061725
APA StyleLiang, Z., & You, G. (2023). Chloroquine and Hydroxychloroquine, as Proteasome Inhibitors, Upregulate the Expression and Activity of Organic Anion Transporter 3. Pharmaceutics, 15(6), 1725. https://doi.org/10.3390/pharmaceutics15061725