
Immunomodulatory Effect and Biological Significance of β-Glucans

 , and
, and
Abstract
1. Introduction
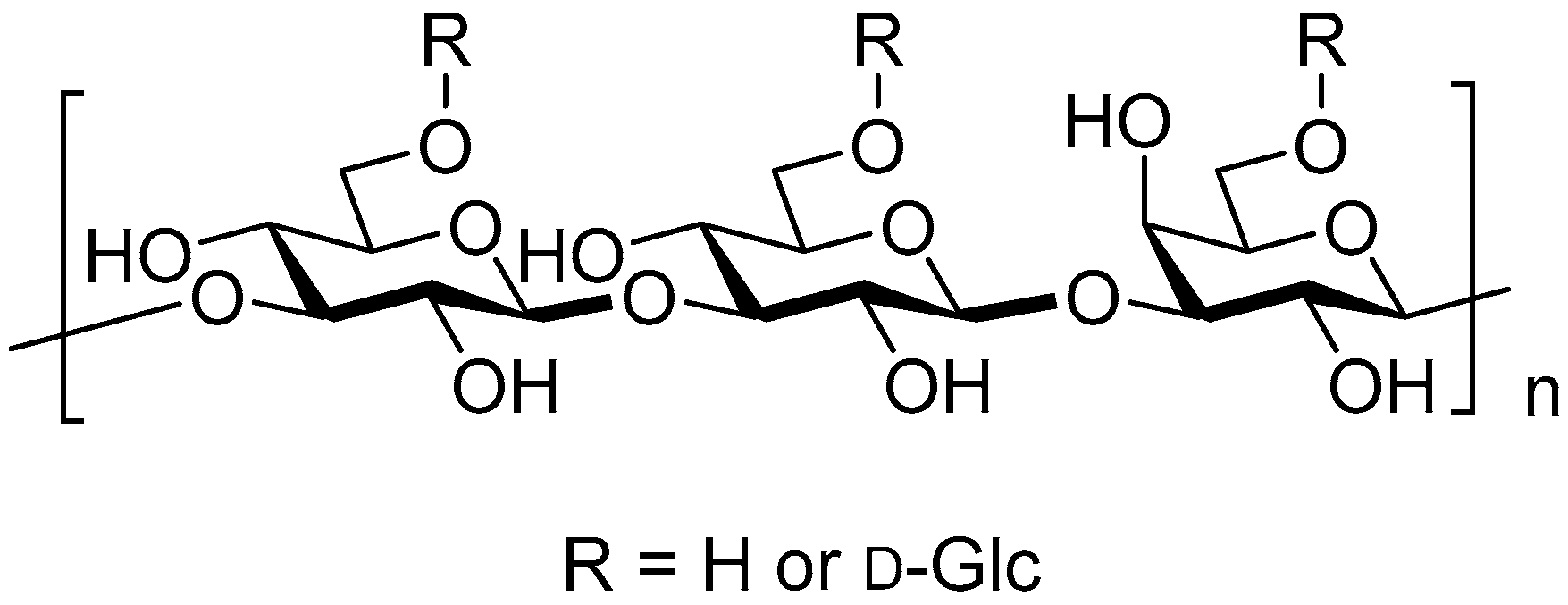
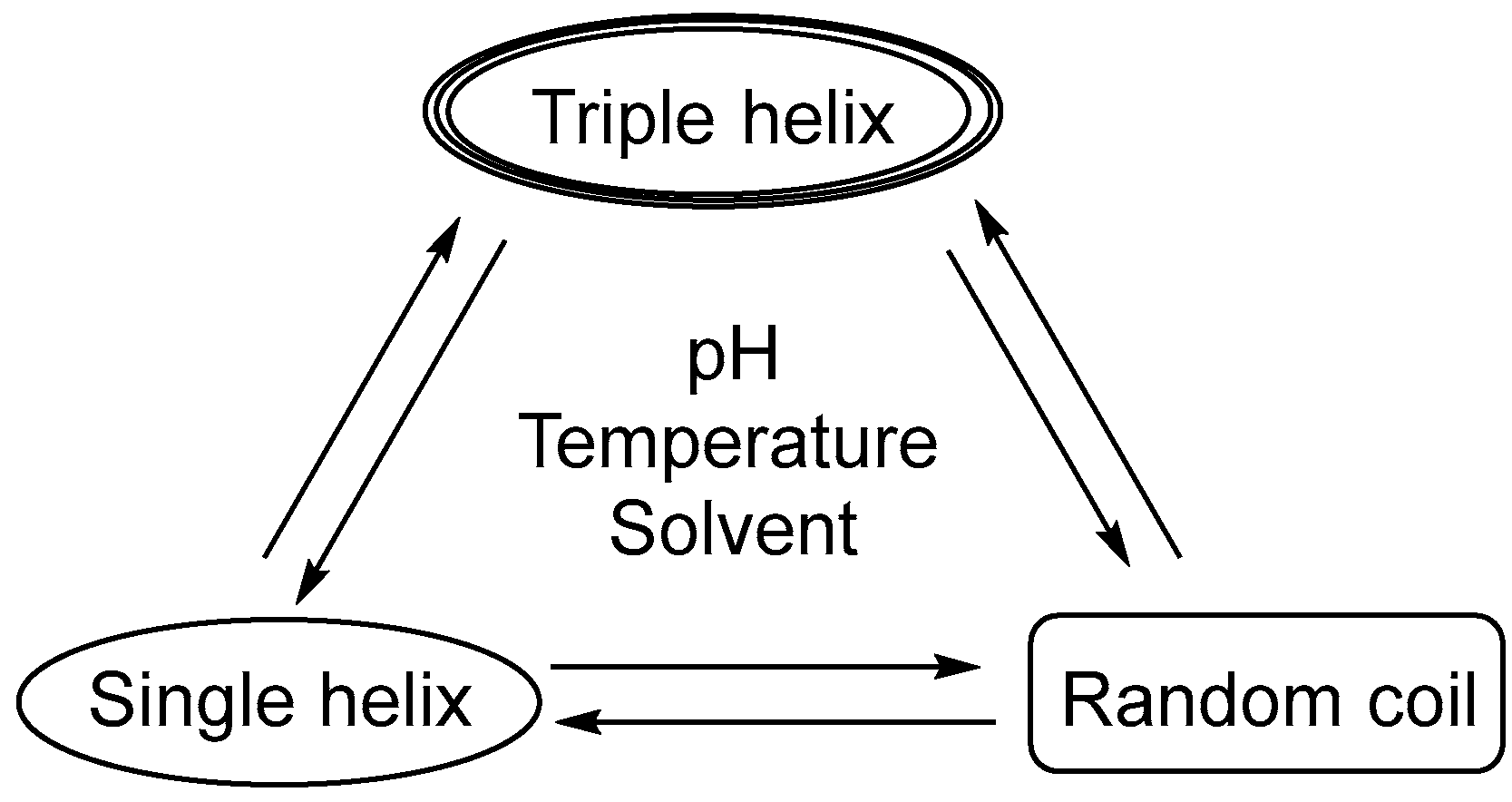
2. β-Glucan Sources and Properties
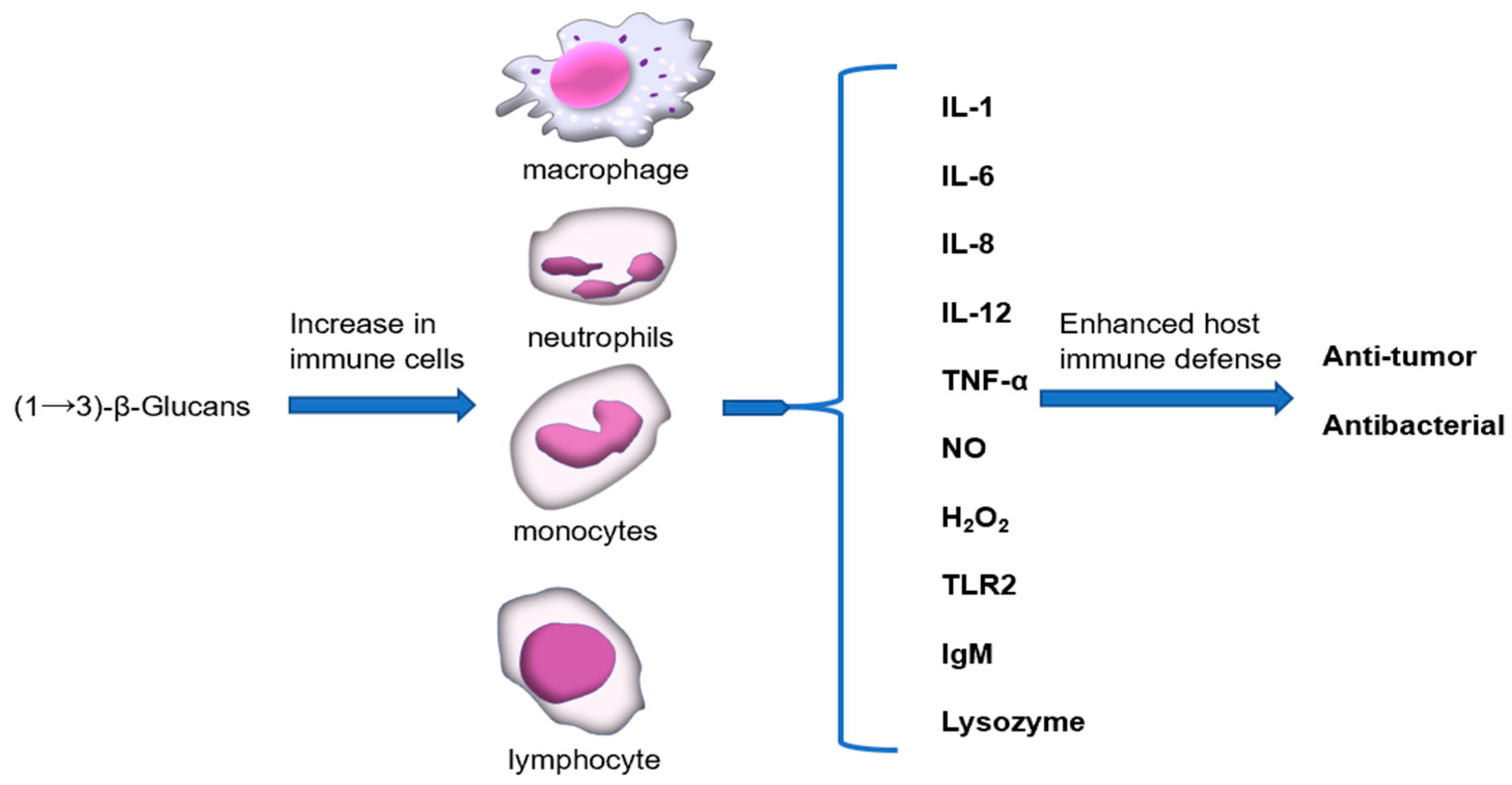
3. Immunostimulatory Properties of (1→3)-β-Glucan
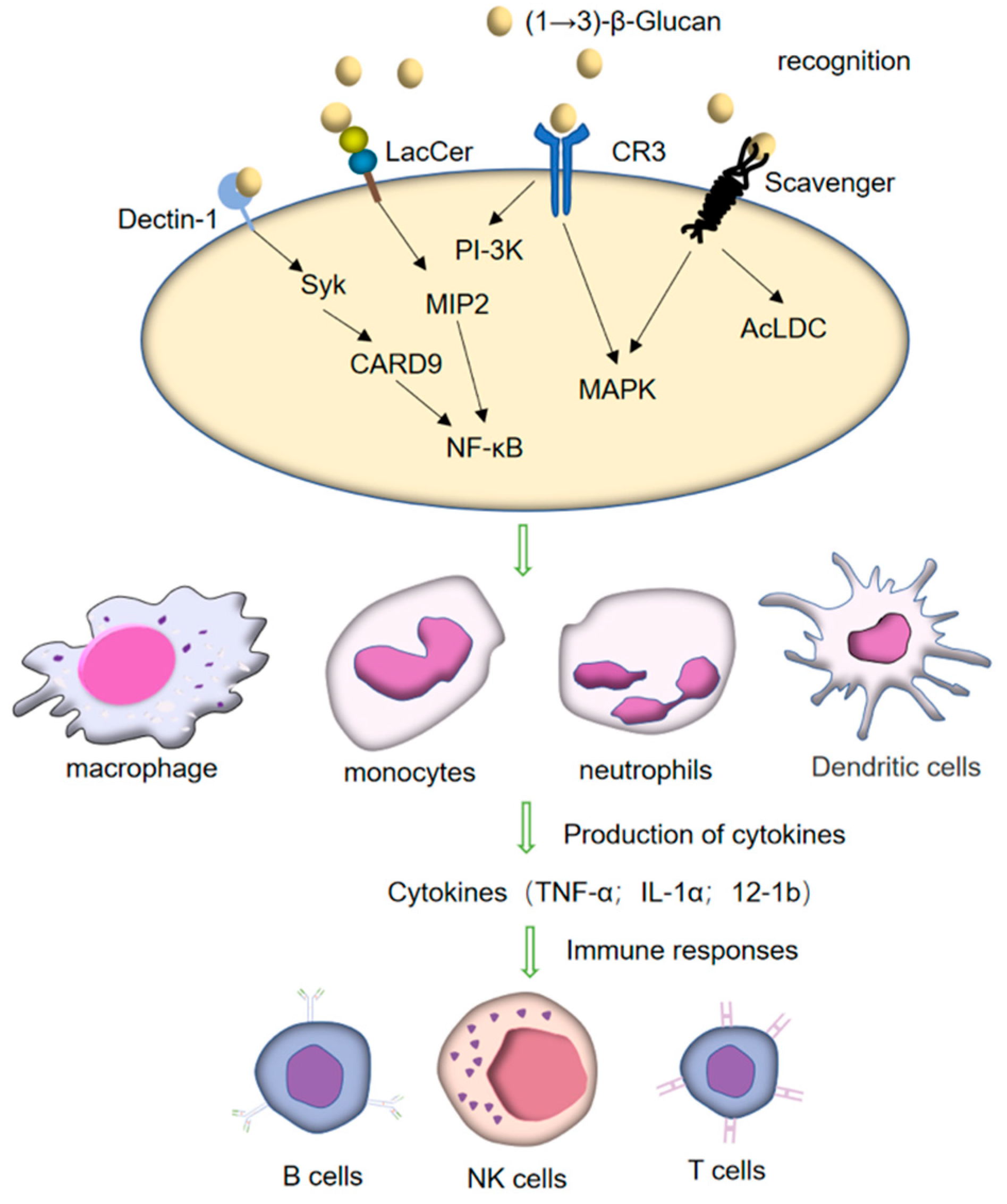
4. Immunoregulatory Receptor of (1→3)-β-Glucan
5. Clinical Applications of (1→3)-β-Glucans
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Demleitner, S.; Kraus, J.; Franz, G. Synthesis and antitumour activity of derivatives of curdlan and lichenan branched at C-6. Carbohydr. Res. 1992, 226, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Větvička, V.C.; Xia, Y.; Coxon, A.; Carroll, M.C.; Mayadas, T.N.; Ross, G.D. β-Glucan, a “specific” biologic response modifier that uses antibodies to target tumors for cytotoxic recognition by leukocyte complement receptor type 3 (CD11b/CD18). J. Immunol. 1999, 163, 3045–3052. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Novák, M. Structural diversity of fungal glucans. Carbohydr. Polym. 2013, 92, 792–809. [Google Scholar] [CrossRef] [PubMed]
- Batbayar, S.; Lee, D.-H.; Kim, H.-W. Immunomodulation of fungal β-glucan in host defense signaling by dectin-1. Biomol. Ther. 2012, 20, 433–445. [Google Scholar] [CrossRef]
- Legentil, L.; Paris, F.; Ballet, C.; Trouvelot, S.; Daire, X.; Vetvicka, V.; Ferrières, V. Molecular interactions of β-(1→3)-glucans with their receptors. Molecules 2015, 20, 9745–9766. [Google Scholar] [CrossRef]
- Akramiene, D.; Kondrotas, A.; Didziapetriene, J.; Kevelaitis, E. Effects of beta-glucans on the immune system. Medicina 2007, 43, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, K.; Masuda, H.; Kaga, N.; Nakayama, H.; Matsumoto, R.; Iwahara, C.; Yoshizaki, F.; Tamaki, Y.; Kobayashi, T.; Hayakawa, T. Properties and functions of lactosylceramide from mouse neutrophils. Glycobiology 2015, 25, 655–668. [Google Scholar] [CrossRef]
- PrabhuDas, M.; Bowdish, D.; Drickamer, K.; Febbraio, M.; Herz, J.; Kobzik, L.; Krieger, M.; Loike, J.; Means, T.K.; Moestrup, S.K. Standardizing scavenger receptor nomenclature. J. Immunol. 2014, 192, 1997–2006. [Google Scholar] [CrossRef]
- Mueller, A.; Raptis, J.; Rice, P.J.; Kalbfleisch, J.H.; Stout, R.D.; Ensley, H.E.; Browder, W.; Williams, D.L. The influence of glucan polymer structure and solution conformation on binding to (1→3)-β-D-glucan receptors in a human monocyte-like cell line. Glycobiology 2000, 10, 339–346. [Google Scholar] [CrossRef]
- Read, S.M.; Currie, G.; Bacic, A. Analysis of the structural heterogeneity of laminarin by electrospray-ionisation-mass spectrometry. Carbohydr. Res. 1996, 281, 187–201. [Google Scholar] [CrossRef]
- Ding, F.; Ishiwata, A.; Ito, Y. Recent advances of the stereoselective bimodal glycosylations for the synthesis of various glucans. In Studies in Natural Products Chemistry (Bioactive Natural Products); Elsevier: Amsterdam, The Netherlands, 2022; Volume 74, pp. 1–40. [Google Scholar] [CrossRef]
- Ding, F.; Ishiwata, A.; Zhou, S.; Zhong, X.; Ito, Y. Unified strategy toward sterecontrolled assembly of various glucans based on bimodal glycosyl donors. J. Org. Chem. 2020, 85, 5536–5558. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, A.; Tanaka, K.; Ao, J.; Ding, F.; Ito, Y. Recent advances in stereoselective 1.2-cis-O-glycosylations. Front. Chem. 2022, 10, 972429. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhong, X.; Guo, A.; Xiao, Q.; Ao, J.; Wanmeng Zhu, W.; Cai, H.; Ishiwata, A.; Ito, Y.; Liu, X.-W.; et al. ZnI2-directed stereocontrolled α-glucosylation. Org. Lett. 2021, 23, 6841–6845. [Google Scholar] [CrossRef]
- Błaszczyk, K.; Wilczak, J.; Harasym, J.; Gudej, S.; Suchecka, D.; Królikowski, T.; Lange, E.; Gromadzka-Ostrowska, J. Impact of low and high molecular weight oat beta-glucan on oxidative stress and antioxidant defense in spleen of rats with LPS induced enteritis. Food Hydrocoll. 2015, 51, 272–280. [Google Scholar] [CrossRef]
- Bacon, J.; Farmer, V.; Jones, D.; Taylor, I.F. The glucan components of the cell wall of baker’s yeast (Saccharomyces cerevisiae) considered in relation to its ultrastructure. Biochem. J. 1969, 114, 557–567. [Google Scholar] [CrossRef]
- Wakshull, E.; Brunke-Reese, D.; Lindermuth, J.; Fisette, L.; Nathans, R.S.; Crowley, J.J.; Tufts, J.C.; Zimmerman, J.; Mackin, W.; Adams, D.S. PGG-Glucan, a soluble β-(1, 3)-glucan, enhances the oxidative burst response, microbicidal activity, and activates an NF-κB-like factor in human PMN: Evidence for a glycosphingolipid β-(1, 3)-glucan receptor. Immunopharmacology 1999, 41, 89–107. [Google Scholar] [CrossRef]
- De la Cruz, J.; Pintor-Toro, J.A.; Benitez, T.; LLobell, A. Purification and characterization of an endo-beta-1, 6-glucanase from Trichoderma harzianum that is related to its mycoparasitism. J. Bacteriol. 1995, 177, 1864–1871. [Google Scholar] [CrossRef][Green Version]
- Karácsonyi, Š.; Kuniak, Ľ. Polysaccharides of Pleurotus ostreatus: Isolation and structure of pleuran, an alkali-insoluble β-D-glucan. Carbohydr. Polym. 1994, 24, 107–111. [Google Scholar] [CrossRef]
- Chihara, G.; Hamuro, J.; Maeda, Y.Y.; Arai, Y.; Fukuoka, F. Fractionation and purification of the polysaccharides with marked antitumor activity, especially lentinan, from Lentinus edodes (Berk.) Sing. (an edible mushroom). Cancer Res. 1970, 30, 2776–2781. [Google Scholar]
- Perlin, A.; Suzuki, S. The structure of lichenin: Selective enzymolysis studies. Can. J. Chem. 1962, 40, 50–56. [Google Scholar] [CrossRef]
- Dong, Q.; Wang, Y.; Shi, L.; Yao, J.; Li, J.; Ma, F.; Ding, K. A novel water-soluble β-D-glucan isolated from the spores of Ganoderma lucidum. Carbohydr. Res. 2012, 353, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.-F.; Zhen, Y.; Ruan, L.; Fang, J.-N. Purification, characterization, and modification of T lymphocyte-stimulating polysaccharide from spores of Ganoderma lucidum. Chem. Pharm. Bull. 2002, 50, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Shi, L.; Ding, K. Structure elucidation and anti-tumor activity in vivo of a polysaccharide from spores of Ganoderma lucidum (Fr.) Karst. Int. J. Biol. Macromol. 2019, 141, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Allendorf, D.J.; Brandley, B. Yeast whole glucan particle (WGP) β-glucan in conjunction with antitumour monoclonal antibodies to treat cancer. Expert Opin. Biol. Ther. 2005, 5, 691–702. [Google Scholar] [CrossRef]
- Guidolin, L.S.; Arce-Gorvel, V.; Ciocchini, A.E.; Comerci, D.J.; Gorvel, J.P. Cyclic β-glucans at the bacteria–host cells interphase: One sugar ring to rule them all. Cell. Microbiol. 2018, 20, e12850. [Google Scholar] [CrossRef]
- Arellano-Reynoso, B.; Lapaque, N.; Salcedo, S.; Briones, G.; Ciocchini, A.E.; Ugalde, R.; Moreno, E.; Moriyón, I.; Gorvel, J.-P. Cyclic β-1,2-glucan is a brucella virulence factor required for intracellular survival. Nat. Immunol. 2005, 6, 618–625. [Google Scholar] [CrossRef]
- Bertin, C.; Pau-Roblot, C.; Courtois, J.; Manso-Silván, L.; Tardy, F.; Poumarat, F.; Citti, C.; Sirand-Pugnet, P.; Gaurivaud, P.; Thiaucourt, F. Highly Dynamic Genomic Loci Drive the Synthesis of Two Types of Capsular or Secreted Polysaccharides within the Mycoplasma mycoides Cluster. Appl. Environ. Microbiol. 2015, 81, 676–687. [Google Scholar] [CrossRef]
- Sedzicki, J.; Ni, D.; Lehmann, F.; Wu, N.; Zenobi, R.; Jung, S.; Stahlberg, H.; Dehio, C. Mechanism of cyclic β-glucan export by ABC transporter Cgt of Brucella. Nat. Struct. Mol. Biol. 2022, 29, 1170–1177. [Google Scholar] [CrossRef]
- Fisher, M.; Yang, L.-X. Anticancer effects and mechanisms of polysaccharide-K (PSK): Implications of cancer immunotherapy. Anticancer Res. 2002, 22, 1737–1754. [Google Scholar]
- Albersheim, P.; Darvill, A.; Augur, C.; Cheong, J.J.; Eberhard, S.; Hahn, M.G.; Marfa, V.; Mohnen, D.; O’Neill, M.A. Oligosaccharins: Oligosaccharide regulatory molecules. Acc. Chem. Res. 1992, 25, 77–83. [Google Scholar] [CrossRef]
- Saito, H.; Misaki, A.; Harada, T. A comparison of the structure of curdlan and pachyman. Agric. Biol. Chem. 1968, 32, 1261–1269. [Google Scholar] [CrossRef]
- Noss, I.; Doekes, G.; Thorne, P.S.; Heederik, D.J.; Wouters, I.M. Comparison of the potency of a variety of β-glucans to induce cytokine production in human whole blood. Innate Immun. 2013, 19, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Ohno, N.; Yadomae, T. Intravenously administered (1→3)-β-D-glucan, SSG, obtained from Sclerotinia sclerotiorum IFO 9395 augments murine peritoneal macrophage functions in vivo. Chem. Pharm. Bull. 1992, 40, 2120–2124. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bae, A.-H.; Lee, S.-W.; Ikeda, M.; Sano, M.; Shinkai, S.; Sakurai, K. Rod-like architecture and helicity of the poly (C)/schizophyllan complex observed by AFM and SEM. Carbohydr. Res. 2004, 339, 251–258. [Google Scholar] [CrossRef]
- Iorio, E.; Torosantucci, A.; Bromuro, C.; Chiani, P.; Ferretti, A.; Giannini, M.; Cassone, A.; Podo, F. Candida albicans cell wall comprises a branched β-d-(1→6)-glucan with β-d-(1→3)-side chains. Carbohydr. Res. 2008, 343, 1050–1061. [Google Scholar] [CrossRef]
- Sasaki, T.; Takasuka, N. Further study of the structure of lentinan, an anti-tumor polysaccharide from Lentinus edodes. Carbohydr. Res. 1976, 47, 99–104. [Google Scholar] [CrossRef]
- Beattie, A.; Hirst, E.; Percival, E. Studies on the metabolism of the Chrysophyceae. Comparative structural investigations on leucosin (chrysolaminarin) separated from diatoms and laminarin from the brown algae. Biochem. J. 1961, 79, 531. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vetvickova, J. Immune-enhancing effects of Maitake (Grifola frondosa) and Shiitake (Lentinula edodes) extracts. Ann. Transl. Med. 2014, 2, 14. [Google Scholar] [CrossRef]
- Di Carlo, F.J.; Fiore, J.V. On the composition of zymosan. Science 1958, 127, 756–757. [Google Scholar] [CrossRef]
- Young, S.-H.; Jacobs, R.R. Sodium hydroxide-induced conformational change in schizophyllan detected by the fluorescence dye, aniline blue. Carbohydr. Res. 1998, 310, 91–99. [Google Scholar] [CrossRef]
- Yadomae, T. Structure and biological activities of fungal beta-1, 3-glucans. Yakugaku Zasshi 2000, 120, 413–431. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, X.; Zhang, L. Thermally induced conformation transition of triple-helical lentinan in NaCl aqueous solution. J. Phys. Chem. B 2008, 112, 10343–10351. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ostroff, G.R.; Lee, C.K.; Agarwal, S.; Ram, S.; Rice, P.A.; Specht, C.A.; Levitz, S.M. Relative contributions of dectin-1 and complement to immune responses to particulate β-glucans. J. Immunol. 2012, 189, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.L.; Rice, P.J.; Graves, B.; Ensley, H.E.; Yu, H.; Brown, G.D.; Gordon, S.; Monteiro, M.A.; Papp-Szabo, E.; Lowman, D.W. Differential high-affinity interaction of dectin-1 with natural or synthetic glucans is dependent upon primary structure and is influenced by polymer chain length and side-chain branching. J. Pharmacol. Exp. Ther. 2008, 325, 115–123. [Google Scholar] [CrossRef]
- Di Luzio, N.R. Update on the immunomodulating activities of glucans. In Springer Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 1985; pp. 387–400. [Google Scholar]
- Wang, W.-J.; Wu, Y.-S.; Chen, S.; Liu, C.-F.; Chen, S.-N. Mushroom β-glucan may immunomodulate the tumor-associated macrophages in the Lewis lung carcinoma. BioMed Res. Int. 2015, 2015, 604385. [Google Scholar] [CrossRef]
- Tian, X.; Tian, J.; Tang, X.; Rui, K.; Zhang, Y.; Ma, J.; Wang, Y.; Xu, H.; Lu, L.; Wang, S. Particulate β-glucan regulates the immunosuppression of granulocytic myeloid-derived suppressor cells by inhibiting NFIA expression. Oncoimmunology 2015, 4, e1038687. [Google Scholar] [CrossRef]
- Miura, T.; Ohno, N.; Miura, N.; Adachi, Y.; Shimada, S.; Yadomae, T. Antigen-specific response of murine immune system toward a yeast β-glucan preparation, zymosan. FEMS Immunol. Med. Microbiol. 1999, 24, 131–139. [Google Scholar] [CrossRef]
- Du, B.; Bian, Z.; Xu, B. Skin health promotion effects of natural beta-glucan derived from cereals and microorganisms: A review. Phytother. Res. 2014, 28, 159–166. [Google Scholar] [CrossRef]
- El Khoury, D.; Cuda, C.; Luhovyy, B.; Anderson, G. Beta glucan: Health benefits in obesity and metabolic syndrome. J. Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef]
- Murphy, E.J.; Rezoagli, E.; Major, I.; Rowan, N.J.; Laffey, J.G. β-glucan metabolic and immunomodulatory properties and potential for clinical application. J. Fungi 2020, 6, 356. [Google Scholar] [CrossRef]
- Du, B.; Lin, C.; Bian, Z.; Xu, B. An insight into anti-inflammatory effects of fungal beta-glucans. Trends Food Sci. Technol. 2015, 41, 49–59. [Google Scholar] [CrossRef]
- Lei, N.; Wang, M.; Zhang, L.; Xiao, S.; Fei, C.; Wang, X.; Zhang, K.; Zheng, W.; Wang, C.; Yang, R. Effects of low molecular weight yeast β-glucan on antioxidant and immunological activities in mice. Int. J. Mol. 2015, 16, 21575–21590. [Google Scholar] [CrossRef] [PubMed]
- Karumuthil-Melethil, S.; Gudi, R.; Johnson, B.M.; Perez, N.; Vasu, C. Fungal β-glucan, a Dectin-1 ligand, promotes protection from type 1 diabetes by inducing regulatory innate immune response. J. Immunol. 2014, 193, 3308–3321. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Sun, X.; Wang, M.; Zhang, C.; Cao, Y.; Mo, G.; Liang, J.; Zhu, S. Quantitative assessment of the effects of beta-glucan consumption on serum lipid profile and glucose level in hypercholesterolemic subjects. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vannucci, L.; Sima, P.; Richter, J. Beta glucan: Supplement or drug? From laboratory to clinical trials. Molecules 2019, 24, 1251. [Google Scholar] [CrossRef]
- Pillemer, L.; Blum, L.; Lepow, I.H.; Ross, O.A.; Todd, E.W.; Wardlaw, A.C. The properdin system and immunity: I. Demonstration and isolation of a new serum protein, properdin, and its role in immune phenomena. Science 1954, 120, 279–285. [Google Scholar] [CrossRef]
- Mahla, R.S.; Reddy, M.C.; Prasad, D.V.R.; Kumar, H. Sweeten PAMPs: Role of sugar complexed PAMPs in innate immunity and vaccine biology. Front. Immunol. 2013, 4, 248. [Google Scholar] [CrossRef]
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef]
- Vetvicka, V. Glucan-immunostimulant, adjuvant, potential drug. World J. Clin. Oncol. 2011, 2, 115. [Google Scholar] [CrossRef]
- Di Luzio, N.; McNamee, R.; Williams, D.; Gilbert, K.; Spanjers, M. Glucan induced inhibition of tumor growth and enhancement of survival in a variety of transplantable and spontaneous murine tumor models. In Macrophages and Lymphocytes; Springer: Berlin/Heidelberg, Germany, 1980; pp. 269–290. [Google Scholar]
- Di Luzio, N.R. Immunopharmacology of glucan: A broad spectrum enhancer of host defense mechanisms. Trends Pharmacol. Sci. 1983, 4, 344–347. [Google Scholar] [CrossRef]
- Demir, G.; Klein, H.; Mandel-Molinas, N.; Tuzuner, N. Beta glucan induces proliferation and activation of monocytes in peripheral blood of patients with advanced breast cancer. Int. Immunopharmacol. 2007, 7, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.L.; Mueller, A.; Browder, W. Glucan-based macrophage stimulators. Clin. Immunother. 1996, 5, 392–399. [Google Scholar] [CrossRef]
- Salama, S.F. Beta-glucan ameliorates gamma-rays induced oxidative injury in male swiss albino rats. Pak. J. Zool. 2011, 43, 933–939. [Google Scholar]
- Iraz, M.; Iraz, M.; Eşrefoğlu, M.; Aydin, M.Ş. Protective effect of\beta-glucan on acute lung injury induced bylipopolysaccharide in rats. Turk. J. Med. Sci. 2015, 45, 261–267. [Google Scholar] [CrossRef]
- Barton, C.; Vigor, K.; Scott, R.; Jones, P.; Lentfer, H.; Bax, H.J.; Josephs, D.H.; Karagiannis, S.N.; Spicer, J.F. Beta-glucan contamination of pharmaceutical products: How much should we accept? Cancer Immunol. Immun. 2016, 65, 1289–1301. [Google Scholar] [CrossRef]
- Sato, M.; Sano, H.; Iwaki, D.; Kudo, K.; Konishi, M.; Takahashi, H.; Takahashi, T.; Imaizumi, H.; Asai, Y.; Kuroki, Y. Direct binding of Toll-like receptor 2 to zymosan, and zymosan-induced NF-κB activation and TNF-α secretion are down-regulated by lung collectin surfactant protein A. J. Immunol. 2003, 171, 417–425. [Google Scholar] [CrossRef]
- Dillon, S.; Agrawal, S.; Banerjee, K.; Letterio, J.; Denning, T.L.; Oswald-Richter, K.; Kasprowicz, D.J.; Kellar, K.; Pare, J.; van Dyke, T. Yeast zymosan, a stimulus for TLR2 and dectin-1, induces regulatory antigen-presenting cells and immunological tolerance. J. Clin. Investig. 2006, 116, 916–928. [Google Scholar] [CrossRef]
- Yoshitomi, H.; Sakaguchi, N.; Kobayashi, K.; Brown, G.D.; Tagami, T.; Sakihama, T.; Hirota, K.; Tanaka, S.; Nomura, T.; Miki, I. A role for fungal β-glucans and their receptor Dectin-1 in the induction of autoimmune arthritis in genetically susceptible mice. J. Exp. Med. 2005, 201, 949–960. [Google Scholar] [CrossRef]
- Ohno, N.; Egawa, Y.; Hashimoto, T.; Adachi, Y.; Yadomae, T. Effect of β-glucans on the nitric oxide synthesis by peritoneal macrophage in mice. Biol. Pharm. Bull. 1996, 19, 608–612. [Google Scholar] [CrossRef]
- Hashimoto, T.; Ohno, N.; Adachi, Y.; Yadomae, T. Enhanced production of inducible nitric oxide synthase by β-glucans in mice. FEMS Immunol. Med. Microbiol. 1997, 19, 131–135. [Google Scholar] [CrossRef]
- Medina-Gali, R.M.; del Mar Ortega-Villaizan, M.; Mercado, L.; Novoa, B.; Coll, J.; Perez, L. Beta-glucan enhances the response to SVCV infection in zebrafish. Dev. Comp. Immunol. 2018, 84, 307–314. [Google Scholar] [CrossRef]
- Petit, J.; Wiegertjes, G.F. Long-lived effects of administering β-glucans: Indications for trained immunity in fish. Dev. Comp. Immunol. 2016, 64, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Xu, Q.; Wang, R.; Qin, L.; Peng, X.; Hu, L.; Liu, Y.; Fang, Z.; Lin, Y.; Xu, S.; et al. Effects of dietary β-glucan supplementation on growth performance and immunological and metabolic parameters of weaned pigs administered with Escherichia coli lipopolysaccharide. Food Funct. 2018, 9, 3338–3343. [Google Scholar] [CrossRef] [PubMed]
- Effects of Bread Yeast Cell Wall Beta-Glucans on Mice with Loperamide-Induced Constipation. J. Med. Food 2019, 22, 1009–1021. [CrossRef] [PubMed]
- Cao, Y.; Sun, Y.; Zou, S.; Duan, B.; Sun, M.; Xu, X. Yeast β-Glucan Suppresses the Chronic Inflammation and Improves the Microenvironment in Adipose Tissues of ob/ob Mice. J. Agric. Food Chem. 2018, 66, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Fan, H.; Yao, M.; Yang, S.; Han, J. Oral administration of yeast β-glucan ameliorates inflammation and intestinal barrier in dextran sodium sulfate-induced acute colitis. J. Funct. Foods 2017, 35, 115–126. [Google Scholar] [CrossRef]
- Pelizon, A.C.; Kaneno, R.; Soares, A.; Meira, D.A.; Sartori, A. Immunomodulatory activities associated with β-glucan derived from Saccharomyces cerevisiae. Physiol. Res. 2005, 54, 557–564. [Google Scholar] [CrossRef]
- Hellinga, A.H.; Tsallis, T.; Eshuis, T.; Triantis, V.; Ulfman, L.H.; van Neerven, R.J.J. In Vitro Induction of Trained Innate Immunity by bIgG and Whey Protein Extracts. Int. J. Mol. Sci. 2020, 21, 9077. [Google Scholar] [CrossRef]
- Kalafati, L.; Kourtzelis, I.; Schulte-Schrepping, J.; Li, X.; Hatzioannou, A.; Grinenko, T.; Hagag, E.; Sinha, A.; Has, C.; Dietz, S.; et al. Innate Immune Training of Granulopoiesis Promotes Anti-tumor Activity. Cell 2020, 183, 771–785.e12. [Google Scholar] [CrossRef]
- Cook, M.T.; Hayball, P.J.; Hutchinson, W.; Nowak, B.; Hayball, J.D. The efficacy of a commercial β-glucan preparation, EcoActiva™, on stimulating respiratory burst activity of head-kidney macrophages from pink snapper (Pagrus auratus), Sparidae. Fish Shellfish Immunol. 2001, 11, 661–672. [Google Scholar] [CrossRef]
- Muthusamy, G.; Joardar, S.N.; Samanta, I.; Isore, D.P.; Roy, B.; Maiti, K. β-glucan from edible mushroom (Pleurotus florida) enhances mucosal immunity in poultry. Adv. Anim. Vet. Sci. 2013, 1, 116–119. [Google Scholar]
- Kwon, K.H.; Kim, K.I.; Jun, W.J.; Shin, D.H.; Cho, H.Y.; Hong, B.S. In Vitro and in Vivo Effects of Macrophage-Stimulatory Polysaccharide from Leaves of Perilla frutescens var crispa. Biol. Pharm. Bull. 2002, 25, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, I.; Tanaka, H.; Kinoshita, A.; Oikawa, S.; Osawa, M.; Yadomae, T. Effect of orally administered β-glucan on macrophage function in mice. Int. J. Immunopharmacol. 1990, 12, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Wolf, A.J.; Underhill, D.M. β-glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef]
- Ross, P.; Farrell, M.P. The Road to Structurally Defined β-Glucans. Chem. Rec. 2021, 21, 3178–3193. [Google Scholar]
- De Marco Castro, E.; Calder, P.C.; Roche, H.M. β-1,3/1,6-Glucans and Immunity: State of the Art and Future Directions. Mol. Nutr. Food Res. 2021, 65, e1901071. [Google Scholar] [CrossRef]
- Chauhan, N.; Tiwari, S.; Iype, T.; Jain, U. An overview of adjuvants utilized in prophylactic vaccine formulation as immunomodulators. Expert Rev. Vaccines 2017, 16, 491–502. [Google Scholar] [CrossRef]
- Petrovsky, N.; Cooper, P.D. Carbohydrate-based immune adjuvants. Expert Rev. Vaccines 2011, 10, 523–537. [Google Scholar] [CrossRef]
- Hu, X.; Liu, R.; Zhu, N. Enhancement of humoral and cellular immune responses by monophosphoryl lipid A (MPLA) as an adjuvant to the rabies vaccine in BALB/c mice. Immunobiology 2013, 218, 1524–1528. [Google Scholar] [CrossRef]
- Córdova-Martínez, A.; Caballero-García, A.; Roche, E.; Noriega, D.C. β-Glucans Could Be Adjuvants for SARS-CoV-2 Virus Vaccines (COVID-19). Int. J. Environ. Res. Public Health 2021, 18, 12636. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, G.A.; Van Kooyk, Y.; Cobb, B.A. Glycobiology of immune responses. Ann. N. Y. Acad. Sci. 2012, 1253, 1–15. [Google Scholar] [CrossRef]
- Hamilos, D.L. Antigen presenting cells. Immunol. Res. 1989, 8, 98–117. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.M.; Gani, A.; Mir, S.A.; Masoodi, F.A.; Khanday, F.A. β-Glucan: A dual regulator of apoptosis and cell proliferation. Int. J. Biol. Macromol. 2021, 182, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Gordon, S. Fungal β-glucans and mammalian immunity. Immunity 2003, 19, 311–315. [Google Scholar] [CrossRef]
- Barreto-Bergter, E.; Figueiredo, R.T. Fungal glycans and the innate immune recognition. Front. Cell. Infect. Microbiol. 2014, 4, 145. [Google Scholar] [CrossRef]
- Ariizumi, K.; Shen, G.-L.; Shikano, S.; Xu, S.; Ritter, R.; Kumamoto, T.; Edelbaum, D.; Morita, A.; Bergstresser, P.R.; Takashima, A. Identification of a novel, dendritic cell-associated molecule, dectin-1, by subtractive cDNA cloning. J. Biol. Chem. 2000, 275, 20157–20167. [Google Scholar] [CrossRef]
- Ikeda, Y.; Adachi, Y.; Ishii, T.; Tamura, H.; Aketagawa, J.; Tanaka, S.; Ohno, N. Blocking Effect of Anti-Dectin-1 Antibodies on the Anti-tumor Activity of 1, 3-β-Glucan and the Binding of Dectin-1 to 1, 3-β-Glucan. Biol. Pharm. Bull. 2007, 30, 1384–1389. [Google Scholar] [CrossRef]
- Willment, J.A.; Gordon, S.; Brown, G.D. Characterization of the human β-glucan receptor and its alternatively spliced isoforms. J. Biol. Chem. 2001, 276, 43818–43823. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. A new receptor for β-glucans. Nature 2001, 413, 36–37. [Google Scholar] [CrossRef]
- Taylor, P.R.; Brown, G.D.; Reid, D.M.; Willment, J.A.; Martinez-Pomares, L.; Gordon, S.; Wong, S.Y. The β-glucan receptor, dectin-1, is predominantly expressed on the surface of cells of the monocyte/macrophage and neutrophil lineages. J. Immunol. 2002, 169, 3876–3882. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Plato, A.; Willment, J.A.; Brown, G.D. C-type lectin-like receptors of the dectin-1 cluster: Ligands and signaling pathways. Int. Rev. Immunol. 2013, 32, 134–156. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhao, Y. The biological role of dectin-1 in immune response. Int. Rev. Immunol. 2007, 26, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Hoving, J.C.; Wilson, G.J.; Brown, G.D. Signalling C-type lectin receptors, microbial recognition and immunity. Cell. Microbiol. 2014, 16, 185–194. [Google Scholar] [CrossRef]
- Suram, S.; Brown, G.D.; Ghosh, M.; Gordon, S.; Loper, R.; Taylor, P.R.; Akira, S.; Uematsu, S.; Williams, D.L.; Leslie, C.C. Regulation of cytosolic phospholipase A2 activation and cyclooxygenase 2 expression in macrophages by the β-glucan receptor. J. Biol. Chem. 2006, 281, 5506–5514. [Google Scholar] [CrossRef]
- Yadav, M.; Schorey, J.S. The β-glucan receptor dectin-1 functions together with TLR2 to mediate macrophage activation by mycobacteria. Blood 2006, 108, 3168–3175. [Google Scholar] [CrossRef]
- Dedloff, M.R.; Effler, C.S.; Holban, A.M.; Gestal, M.C. Use of biopolymers in mucosally-administered vaccinations for respiratory disease. Materials 2019, 12, 2445. [Google Scholar] [CrossRef]
- De Smet, R.; Demoor, T.; Verschuere, S.; Dullaers, M.; Ostroff, G.R.; Leclercq, G.; Allais, L.; Pilette, C.; Dierendonck, M.; De Geest, B.G. β-Glucan microparticles are good candidates for mucosal antigen delivery in oral vaccination. J. Control. Release 2013, 172, 671–678. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Reyes, C.N.; Becker, C.A.; Katsumoto, T.R.; Ma, J.; Wolf, A.J.; Bose, N.; Chan, A.S.; Magee, A.S.; Danielson, M.E. Activation of the innate immune receptor Dectin-1 upon formation of a ‘phagocytic synapse’. Nature 2011, 472, 471–475. [Google Scholar] [CrossRef]
- Hoffman, O.; Standing, J.; Limper, A. Pneumocystis carinii stimulates tumor necrosis factor-alpha release from alveolar macrophages through a beta-glucan-mediated mechanism. J. Immunol. 1993, 150, 3932–3940. [Google Scholar] [CrossRef] [PubMed]
- Kashem, S.W.; Igyártó, B.Z.; Gerami-Nejad, M.; Kumamoto, Y.; Mohammed, J.; Jarrett, E.; Drummond, R.A.; Zurawski, S.M.; Zurawski, G.; Berman, J. Candida albicans morphology and dendritic cell subsets determine T helper cell differentiation. Immunity 2015, 42, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.; Giovannini, G.; De Luca, A.; D’angelo, C.; Casagrande, A.; Iannitti, R.G.; Ricci, G.; Cunha, C.; Romani, L. Dectin-1 isoforms contribute to distinct Th1/Th17 cell activation in mucosal candidiasis. Cell. Mol. Immunol. 2012, 9, 276–286. [Google Scholar] [CrossRef]
- Sarma, J.V.; Ward, P.A. The complement system. Cell Tissue Res. 2011, 343, 227–235. [Google Scholar] [CrossRef]
- Thornton, B.P.; Vĕtvicka, V.; Pitman, M.; Goldman, R.C.; Ross, G.D. Analysis of the sugar specificity and molecular location of the beta-glucan-binding lectin site of complement receptor type 3 (CD11b/CD18). J. Immunol. 1996, 156, 1235–1246. [Google Scholar] [CrossRef]
- Ina, K.; Kataoka, T.; Ando, T. The use of lentinan for treating gastric cancer. Anti-Cancer Agents Med. (Former. Curr. Med. Chem.—Anti-Cancer Agents) 2013, 13, 681–688. [Google Scholar] [CrossRef]
- Le Cabec, V.; Carréno, S.; Moisand, A.; Bordier, C.; Maridonneau-Parini, I. Complement receptor 3 (CD11b/CD18) mediates type I and type II phagocytosis during nonopsonic and opsonic phagocytosis, respectively. J. Immunol. 2002, 169, 2003–2009. [Google Scholar] [CrossRef] [PubMed]
- Michalek, M.; Melican, D.; Brunke-Reese, D.; Langevin, M.; Lemerise, K.; Galbraith, W.; Patchen, M.; Mackin, W. Activation of rat macrophages by Betafectin PGG-glucan requires cross-linking of membrane receptors distinct from complement receptor three (CR3). J. Leukoc. Biol. 1998, 64, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Baert, K.; Sonck, E.; Goddeeris, B.M.; Devriendt, B.; Cox, E. Cell type-specific differences in β-glucan recognition and signalling in porcine innate immune cells. Dev. Comp. Immunol. 2015, 48, 192–203. [Google Scholar] [CrossRef]
- Peiser, L.; Mukhopadhyay, S.; Gordon, S. Scavenger receptors in innate immunity. Curr. Opin. Immunol. 2002, 14, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Means, T.K. Fungal pathogen recognition by scavenger receptors in nematodes and mammals. Virulence 2010, 1, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, J.W.; Lindermuth, J.; Fish, P.A.; Palace, G.P.; Stevenson, T.T.; DeMong, D.E. A novel carbohydrate-glycosphingolipid interaction between a β-(1–3)-glucan immunomodulator, PGG-glucan, and lactosylceramide of human leukocytes. J. Biol. Chem. 1998, 273, 22014–22020. [Google Scholar] [CrossRef] [PubMed]
- Gudej, S.; Filip, R.; Harasym, J.; Wilczak, J.; Dziendzikowska, K.; Oczkowski, M.; Jałosińska, M.; Juszczak, M.; Lange, E.; Gromadzka-Ostrowska, J. Clinical outcomes after oat beta-glucans dietary treatment in gastritis patients. Nutrients 2021, 13, 2791. [Google Scholar] [CrossRef] [PubMed]
- Cheung, I.Y.; Cheung, N.V.; Modak, S.; Mauguen, A.; Feng, Y.; Basu, E.; Roberts, S.S.; Ragupathi, G.; Kushner, B.H. Survival Impact of Anti-GD2 Antibody Response in a Phase II Ganglioside Vaccine Trial Among Patients with High-Risk Neuroblastoma with Prior Disease Progression. J. Clin. Oncol. 2021, 39, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, S.D.V.; Cordeiro, S.L.; Cavalcanti, J.E.C.; Melchuna, K.M.; Lima, A.; Filho, I.A.; Medeiros, A.C.; Rocha, K.B.F.; Oliveira, E.M.; Faria, E.D.B.; et al. Effects of purified Saccharomyces cerevisiae (1→3)-β-glucan on venous ulcer healing. Int. J. Mol. Sci. 2012, 13, 8142–8158. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Veronesi, M.; Strocchi, E.; Grandi, E.; Rizzoli, E.; Poli, A.; Marangoni, F.; Borghi, C. A randomized Placebo-Controlled Clinical Trial to Evaluate the Medium-Term Effects of Oat Fibers on Human Health: The Beta-Glucan Effects on Lipid Profile, Glycemia and inTestinal Health (BELT) Study. Nutrients 2020, 12, 686. [Google Scholar] [CrossRef]
- Bobovčák, M.; Kuniaková, R.; Gabriž, J.; Majtán, J. Effect of Pleuran (β-glucan from Pleurotus ostreatus) supplementation on cellular immune response after intensive exercise in elite athletes. Appl. Physiol. Nutr. Metab. 2010, 35, 755–762. [Google Scholar] [CrossRef]
- Fuller, R.; Moore, M.V.; Lewith, G.; Stuart, B.L.; Ormiston, R.V.; Fisk, H.L.; Noakes, P.S.; Calder, P.C. Yeast-derived β-1,3/1,6 glucan, upper respiratory tract infection and innate immunity in older adults. Nutrition 2017, 39–40, 30–35. [Google Scholar] [CrossRef]
- Fazilaty, Z.; Chenari, H.; Shariatpanahi, Z.V. Effect of ß-glucan on serum levels of IL-12, hs-CRP, and clinical outcomes in multiple-trauma patients: A prospective randomized study. Ulus. Travma Acil Cerrahi Derg. 2018, 24, 287–293. [Google Scholar]
- Lehne, G.; Haneberg, B.; Gaustad, P.; Johansen, P.W.; Preus, H.; Abrahamsen, T.G. Oral administration of a new soluble branched beta-1,3-D-glucan is well tolerated and can lead to increased salivary concentrations of immunoglobulin A in healthy volunteers. Clin. Exp. Immunol. 2006, 143, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Auinger, A.; Riede, L.; Bothe, G.; Busch, R.; Gruenwald, J. Yeast (1,3)-(1,6)-beta-glucan helps to maintain the body’s defence against pathogens: A double-blind, randomized, placebo-controlled, multicentric study in healthy subjects. Eur. J. Nutr. 2013, 52, 1913–1918. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.G.; Kim, Y.S.; Lee, Y.J.; Ahn, H.Y.; Kim, M.; Kim, M.; Cho, M.J.; Cho, Y.; Lee, J.H. Effect of Immune-Enhancing Enteral Nutrition Enriched with or without Beta-Glucan on Immunomodulation in Critically Ill Patients. Nutrients 2016, 8, 336. [Google Scholar] [CrossRef]
- Carpenter, K.C.; Breslin, W.L.; Davidson, T.; Adams, A.; McFarlin, B.K. Baker’s yeast β-glucan supplementation increases monocytes and cytokines post-exercise: Implications for infection risk? Br. J. Nutr. 2013, 109, 478–486. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linkage | (Glucan) | Source | References |
|---|---|---|---|
| β-(1→2) | Mycoplasma capricolum and Mycoplasma leachii | [26,27,28,29] | |
| β-(1→4) | (cellulose) | Plant (common) | |
| β-(1→6) | (pustulan) | Gyrophera esculenta | [18] |
| β-(1→3)-β-(1→4)-β-(1→6) | (PSK) | Trametes versicolor | [30] |
| β-(1→6)-β-(1→4) | (LC11) | Lentinus edodes | [20] |
| β-(1→6)-β-(1→4) | (GLSA50-1B) | Ganoderma lucidum | [22] |
| β-(1→6)-β-(1→3) | (phytoalexins) | P. megasperma mycelial | [31] |
| β-(1→3)-glucans | |||
| β-(1→3) | (pachymaran) | Poria cocos | [32] |
| β-(1→3) | (curdlan) | Alcaligenes faecali | [33] |
| β-(1→3) | (SSG) | Sclerotinia sclerotiorum | [34] |
| β-(1→3)-β-(1→4) | (hemicellulose) | Plant (common) | |
| β-(1→3)-β-(1→4) | (lichenan) | Cetraria islandica | [21] |
| β-(1→3)-β-(1→4) | (Barley glucan) | Hordeum vulgare | [33] |
| β-(1→3)-β-(1→4) | (Oat glucan) | Avena sativa | [15,16] |
| β-(1→3)-β-(1→6) | (schizophillan) | Schizophyllum commune | [35] |
| Candida albicans | [36] | ||
| β-(1→3)-β-(1→6) | (lentinan) | Lentinula edodes | [37] |
| β-(1→3)-β-(1→6) | (Yeast glucan) | Saccharomyces cerevisiae | [11,12,13,14] |
| β-(1→3)-β-(1→6) | (scleroglucan) | Sclerotium glucanicum | [9,10] |
| β-(1→3)-β-(1→6) | (laminaran) | Laminaria sigitota | [9,10,38] |
| β-(1→3)-β-(1→6) | (grifolan) | Grifola frondosa | [39] |
| β-(1→3)-β-(1→6) | (pachyman) | Poria cocos | [32] |
| β-(1→3)-β-(1→6) | (PSGL-I-1A) | Ganoderma lucidum | [23] |
| β-(1→3)-β-(1→6) | (WGLP) | Ganoderma lucidum | [24] |
| β-(1→3)-β-(1→6) | (PGG) | Saccharomyces cerevisiae | [17] |
| β-(1→3)-β-(1→4)-β-(1→6) | (pleuran) | Pleuritus ostreatus | [19] |
| β-(1→3)-β-(1→6) | (WGP) | Saccharomyces cerevisiae | [25] |
| β-(1→3)-β-(1→6) | (zymosan) | Saccharomyces cerevisiae | [40] |
| Study | Participants | Intervention | Main Findings | Ref. |
|---|---|---|---|---|
| Gudej et al. (2021) | 129 participants with dyspepsia | Oat β-D-glucan Supplements | Reduced mucosal damage | [127] |
| Cheung et al. (2022) | One hundred two patients with HR-NB | Oral β-glucan Supplements | Elicited robust antibody responses in patients | [128] |
| Medeiros et al. (2012) | 12 patients who had venous ulcers | Saccharomyces cerevisiae (1→3)-β-glucan Supplements | Enhanced venous ulcer healing and increased epithelial hyperplasia | [129] |
| Cicero et al. (2020) | 83 Italian with a moderate hypercholesterolemia and a low cardiovascular risk | Oat β-glucan Supplements | Reducing LDL-C, TC and non-HDL-C | [130] |
| Bobovčák et al. (2010) | 20 elite athletes | Pleurotus ostreatus β-glucan Supplements | May play a role in modulating exercise-induced changes in natural killer cell activity | [131] |
| Fuller et al. (2017) | 49 participants | Yeast β-1,3/1,6 glucan Supplements | Prevent the occurrence or reduce the severity of upper respiratory tract infection | [132] |
| Fazilaty et al. (2018) | 40 multiple trauma patients | β-glucan Supplements | Increase serum levels of IL-12 | [133] |
| Lehne et al. (2006) | Eighteen healthy volunteers | Yeast β-1,3-D-glucan Supplements | Increased the immunoglobulin A | [134] |
| Auinger et al. (2013) | 162 healthy participants | Yeast (1,3)-(1,6)- β-D-glucan Supplements | Increased the body’s potential to defend against invading pathogens | [135] |
| Lee et al. (2016) | 30 patients in critically ill | β-glucan Supplements | Increases in natural killer (NK) cell activities | [136] |
| Carpenter et al. (2012) | 60 recreationally active men and women | Yeast β-glucan supplementation | Stimulated cytokine production, increased total (CD14+) | [137] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, X.; Wang, G.; Li, F.; Fang, S.; Zhou, S.; Ishiwata, A.; Tonevitsky, A.G.; Shkurnikov, M.; Cai, H.; Ding, F. Immunomodulatory Effect and Biological Significance of β-Glucans. Pharmaceutics 2023, 15, 1615. https://doi.org/10.3390/pharmaceutics15061615
Zhong X, Wang G, Li F, Fang S, Zhou S, Ishiwata A, Tonevitsky AG, Shkurnikov M, Cai H, Ding F. Immunomodulatory Effect and Biological Significance of β-Glucans. Pharmaceutics. 2023; 15(6):1615. https://doi.org/10.3390/pharmaceutics15061615
Chicago/Turabian StyleZhong, Xuemei, Guoqing Wang, Fu Li, Sixian Fang, Siai Zhou, Akihiro Ishiwata, Alexander G. Tonevitsky, Maxim Shkurnikov, Hui Cai, and Feiqing Ding. 2023. "Immunomodulatory Effect and Biological Significance of β-Glucans" Pharmaceutics 15, no. 6: 1615. https://doi.org/10.3390/pharmaceutics15061615
APA StyleZhong, X., Wang, G., Li, F., Fang, S., Zhou, S., Ishiwata, A., Tonevitsky, A. G., Shkurnikov, M., Cai, H., & Ding, F. (2023). Immunomodulatory Effect and Biological Significance of β-Glucans. Pharmaceutics, 15(6), 1615. https://doi.org/10.3390/pharmaceutics15061615