
In Vitro Killing Activities of Anidulafungin and Micafungin with and without Nikkomycin Z against Four Candida auris Clades


 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. Whole Genome Sequencing
2.3. Antifungal Susceptibility Testing
2.4. Time–Kill Studies
2.5. Combination of Anidulafungin and Micafungin plus Nikkomycin Z in Time–Kill Studies
2.6. Statistical Analysis
3. Results
3.1. Anidulafungin, Micafungin and Nikkomycin Z MIC Values against four Candida auris Clades
3.1.1. Nikkomycin Z Killing Activity against four Candida auris Clades
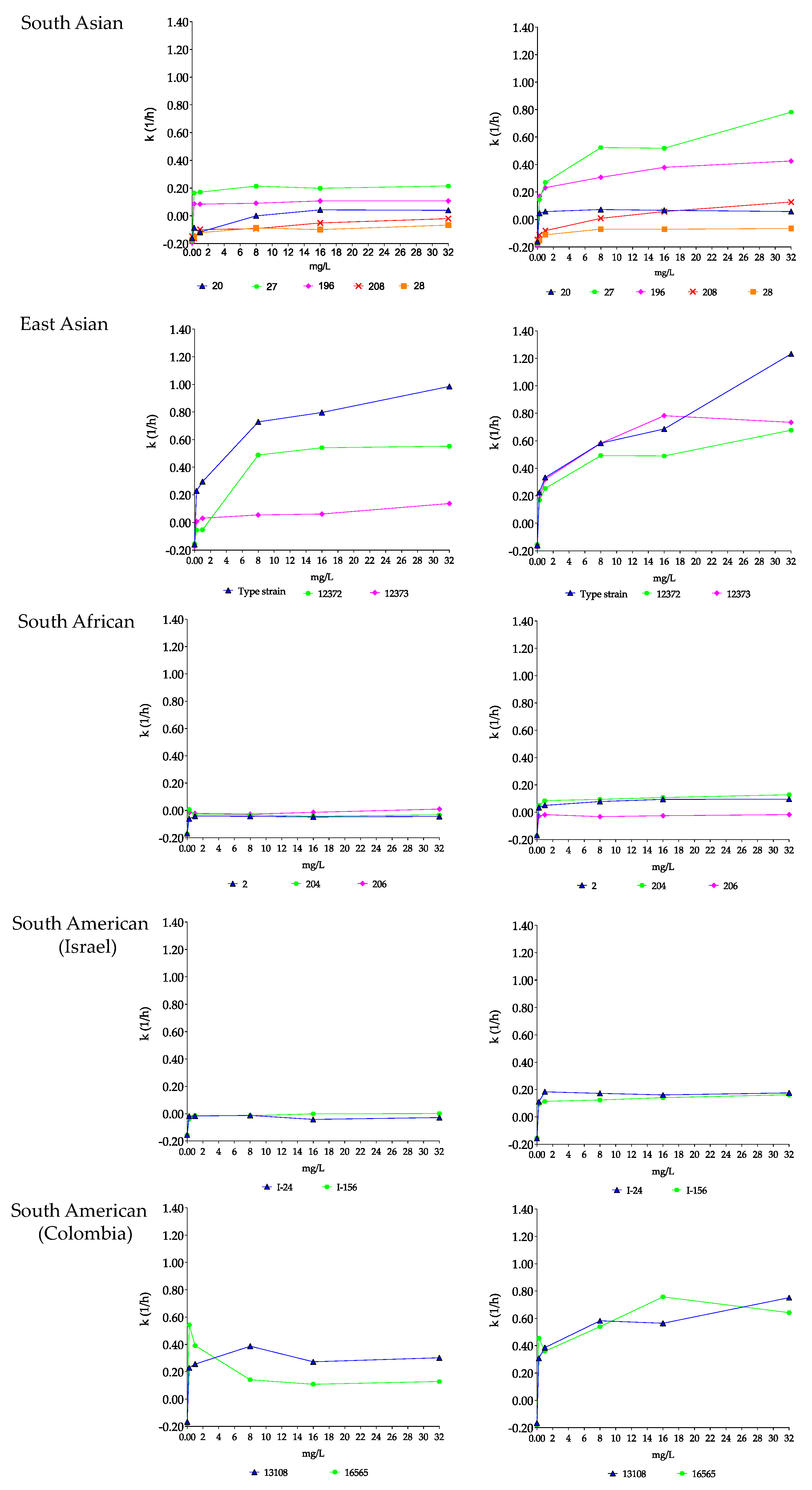
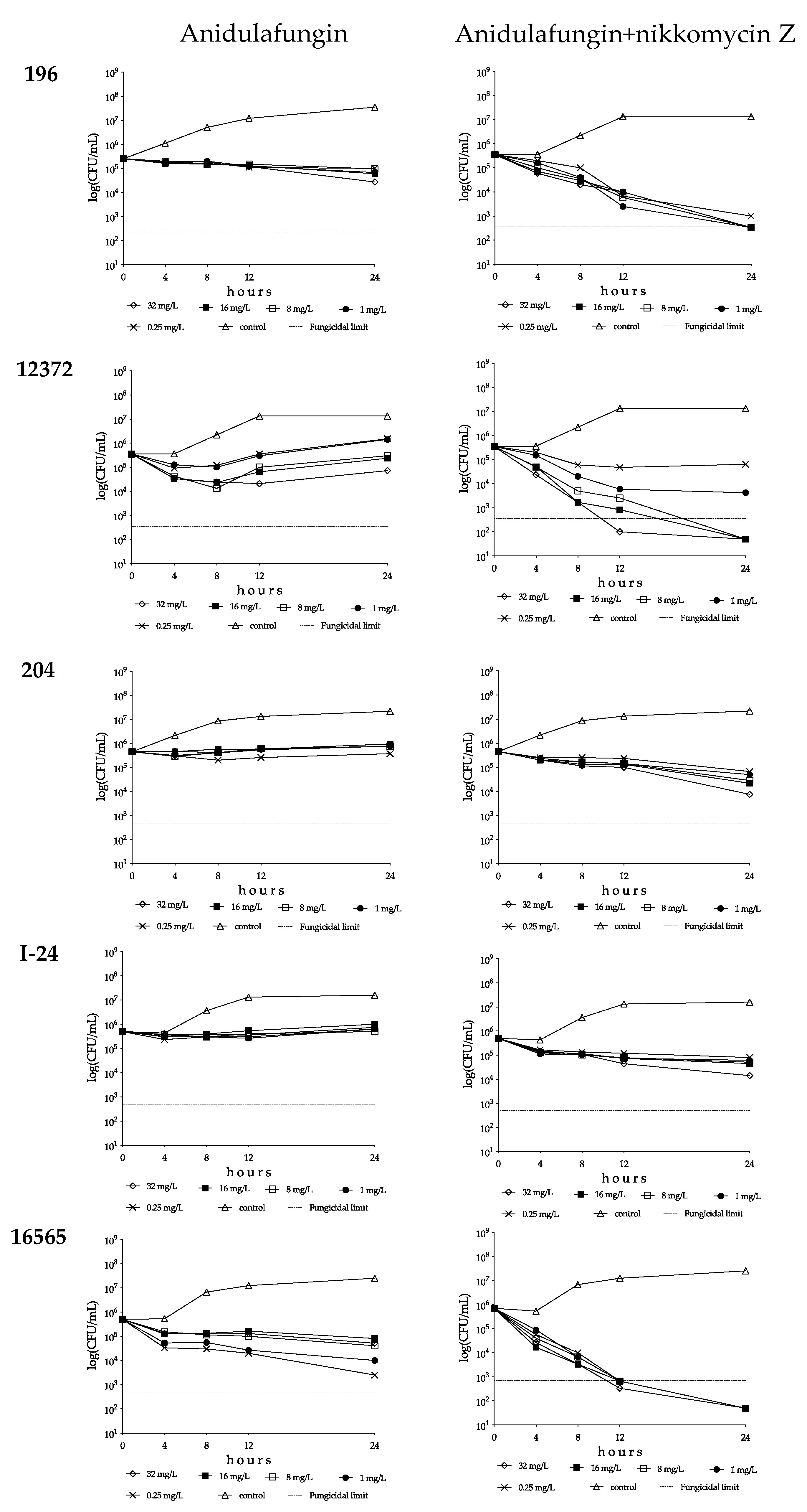
3.1.2. Anidulafungin and Anidulafungin plus Nikkomycin Z Killing Activities against Four Candida auris Clades
South Asian Clade
East Asian Clade
South African Clade
South American Clade
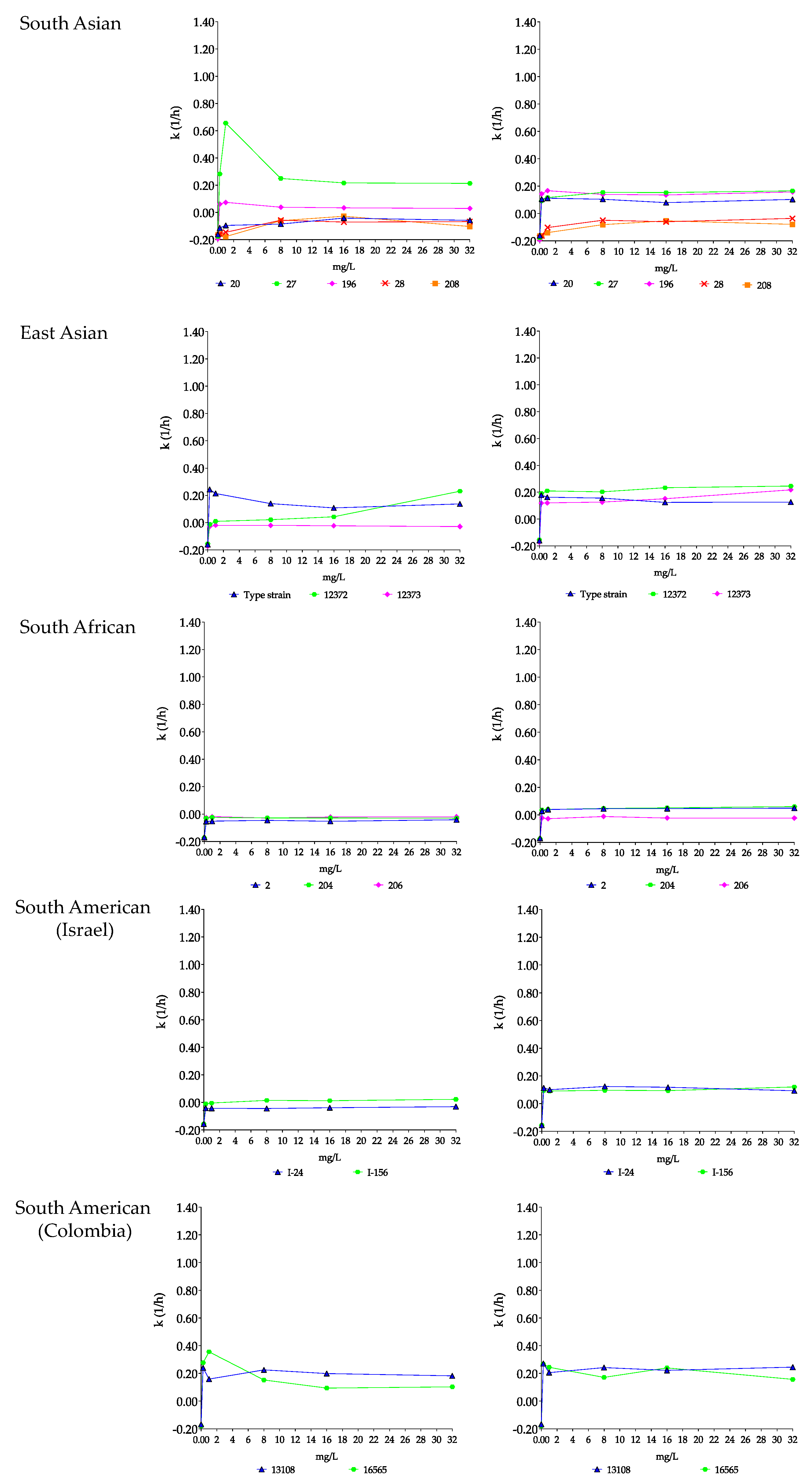
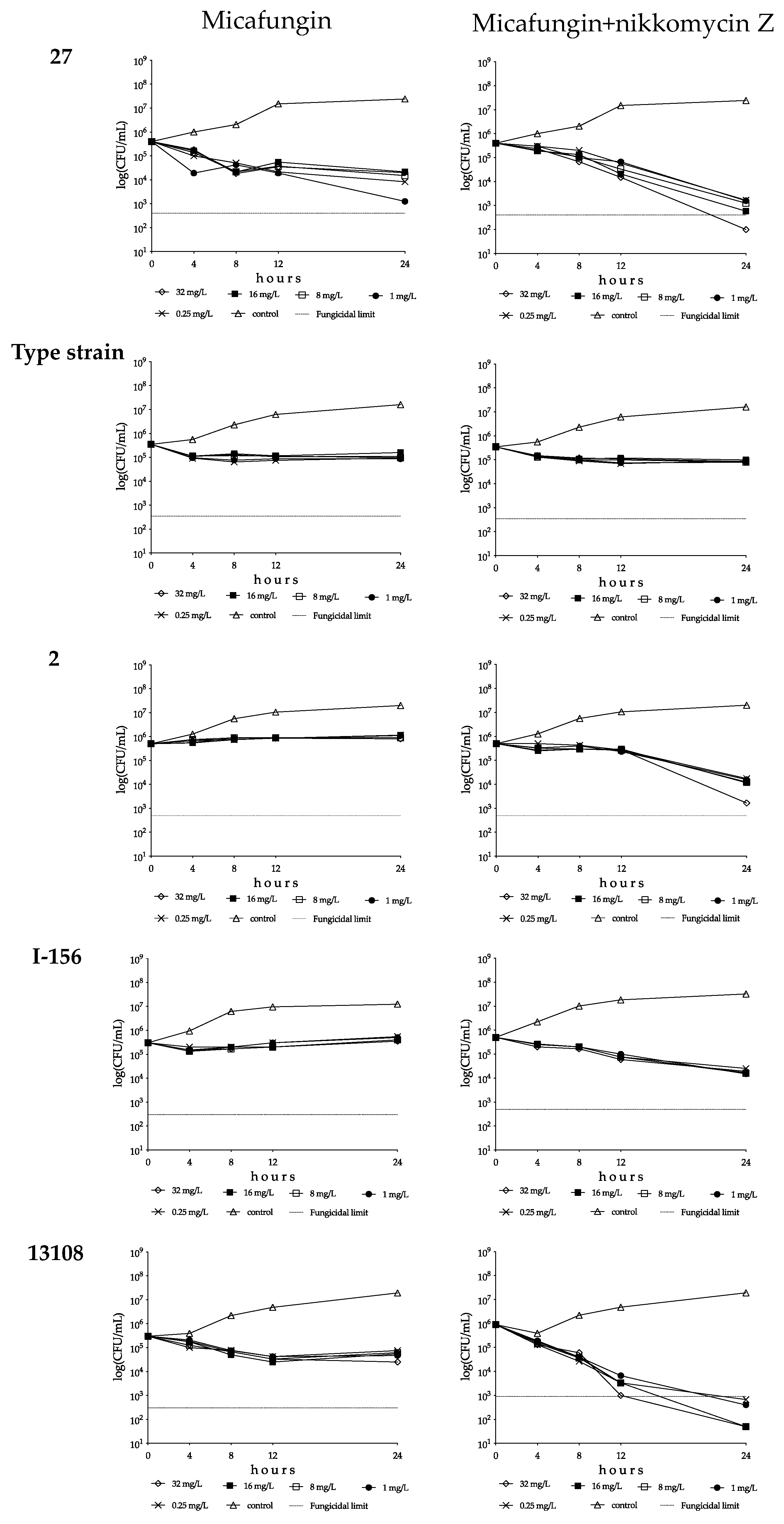
3.1.3. Micafungin and Micafungin plus Nikkomycin Z Killing Activities against Four Candida auris Clades
South Asian Clade
East Asian Clade
South African Clade
South American Clade
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Fungal Priority Pathogens List to Guide Research, Development and Public Health Action; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Khojasteh, S.; Jafarzdeh, J.; Hosseini, S.A.; Haghani, I.; Turki, H.; Gharehbolagh, S.A.; Abastabar, M.; Mahmoudi, S. Candida auris and COVID-19: A health threatening combination. Curr. Med Mycol. 2022, 8, 44–50. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous emergence of multidrug-resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef]
- Osei, S.J. Candida auris: A systematic review and meta-analysis of current updates on an emerging multidrug-resistant pathogen. MicrobiologyOpen 2018, 7, e578. [Google Scholar] [CrossRef]
- Armstrong, P.A.; Rivera, S.M.; Escandon, P.; Caceres, D.H.; Chow, N.; Stuckey, M.J.; Diaz, J.; Gomez, A.; Velez, N.; Espinosa-Bode, A.; et al. Hospital-associated multicenter outbreak of emerging fungus Candida auris, Colombia, 2016. Emerg. Infect. Dis. 2019, 25, 1339–1346. [Google Scholar] [CrossRef]
- Ruiz-Gaitán, A.; Moret, A.M.; Tasias-Pitarch, M.; Aleixandre, A.I.; Martinez-Morel, H.; Calabuig, E.; Salavert-Lleti, M.; Ramirez, P.; Lopez-Hontangas, J.L.; Hagen, F.; et al. An outbreak due to Candida auris with prolonged colonisation and candidaemia in a tertiary care European hospital. Mycoses 2018, 61, 498–505. [Google Scholar] [CrossRef]
- CDC. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html (accessed on 31 January 2023).
- Shastri, P.S.; Shankarnarayan, S.A.; Oberoi, J.; Rudramurthy, S.M.; Wattal, C.; Chakrabarti, A. Candida auris candidaemia in an intensive care unit—Prospective observational study to evaluate epidemiology, risk factors, and outcome. J. Crit. Care 2020, 57, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Lozano, H.; Treviño-Rangel, R.D.J.; González, G.M.; Elizondo, M.T.; Medrano, R.; Bocanegra, M.C.; Lara, C.E.; Gaona, N.; Castilleja, F.; Torre-Amione, G.; et al. Outbreak of Candida auris infection in a COVID-19 hospital in Mexico. Clin. Microbiol. Infect. 2021, 27, 813–816. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Tarai, B.; Singh, A.; Sharma, A. Multidrug-resistant Candida auris infections in critically ill Coronavirus disease patients, India, April–July 2020. Emerg. Infect. Dis. 2020, 26, 2694–2696. [Google Scholar] [CrossRef] [PubMed]
- Hanson, B.M.; Dinh, A.Q.; Tran, T.T.; Arenas, S.; Pronty, D.; Gershengorn, H.B.; Ferreira, T.; Arias, C.A.; Shukla, B.S. Candida auris Invasive Infections during a COVID-19 Case Surge. Antimicrob. Agents Chemother. 2021, 65, e0114621. [Google Scholar] [CrossRef] [PubMed]
- Bandara, N.; Samaranayake, L. Emerging and future strategies in the management of recalcitrant Candida auris. Med. Mycol. 2022, 60, myac008. [Google Scholar] [CrossRef] [PubMed]
- Rauseo, A.M.; Coler-Reilly, A.; Larson, L.; Spec, A. Hope on the Horizon: Novel Fungal Treatments in Development. Open Forum Infect. Dis. 2020, 7, ofaa016. [Google Scholar] [CrossRef]
- Kim, J.H.; Cheng, L.W.; Chan, K.L.; Tam, C.C.; Mahoney, N.; Friedman, M.; Shilman, M.M.; Land, K.M. Antifungal Drug Repurposing. Antibiotics 2020, 9, 812. [Google Scholar] [CrossRef]
- Fioriti, S.; Brescini, L.; Pallotta, F.; Canovari, B.; Morroni, G.; Barchiesi, F. Antifungal Combinations against Candida Species: From Bench to Bedside. J. Fungi 2022, 8, 1077. [Google Scholar] [CrossRef]
- Bentz, M.L.; Nunnally, N.; Lockhart, S.R.; Sexton, D.J.; Berkow, E.L. Antifungal activity of nikkomycin Z against Candida auris. J. Antimicrob. Chemother. 2021, 76, 1495–1497. [Google Scholar] [CrossRef] [PubMed]
- Nix, D.E.; Swezey, R.R.; Hector, R.; Galgiani, J.N. Pharmacokinetics of Nikkomycin Z after Single Rising Oral Doses. Antimicrob. Agents Chemother. 2009, 53, 2517–2521. [Google Scholar] [CrossRef]
- Larwood, D.J. Nikkomycin Z-Ready to meet the promise? J. Fungi 2020, 6, 261. [Google Scholar] [CrossRef] [PubMed]
- Szilágyi, J.; Földi, R.; Bayegan, S.; Kardos, G.; Majoros, L. Effect of nikkomycin Z and 50% human serum on the killing activity of high-concentration caspofungin against Candida species using time-kill methodology. J. Chemother. 2012, 24, 18–25. [Google Scholar] [CrossRef]
- Borman, A.M.; Szekely, A.; Johnson, E.M. Comparative Pathogenicity of United Kingdom Isolates of the Emerging Pathogen Candida auris and Other Key Pathogenic Candida Species. Msphere 2016, 18, e00189-16. [Google Scholar] [CrossRef]
- Kovács, R.; Tóth, Z.; Locke, J.B.; Forgacs, L.; Kardos, G.; Nagy, F.; Borman, A.M.; Majoros, L. Comparison of in vitro killing activity of rezafungin, anidulafungin, caspofungin, and micafungin against four Candida auris clades in RPMI-1640 in the absence and presence of human serum. Microorganisms 2021, 9, 863. [Google Scholar] [CrossRef] [PubMed]
- Forgács, L.; Borman, A.M.; Prépost, E.; Tóth, Z.; Kardos, G.; Kovács, R.; Szekely, A.; Nagy, F.; Kovacs, I.; Majoros, L. Comparison of in vivo pathogenicity of four Candida auris clades in a neutropenic bloodstream infection murine model. Emerg. Microbes Infect. 2020, 9, 1160–1169. [Google Scholar] [CrossRef]
- Balla, N.; Kovács, F.; Balázs, B.; Borman, A.M.; Bozó, A.; Tóth, Z.; Kobaissi, O.; Majoros, L.; Kovács, R. Synergistic Interaction of Caspofungin Combined with Posaconazole against FKS Wild-Type and Mutant Candida auris Planktonic Cells and Biofilms. Antibiotics 2022, 11, 1601. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed.; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antifungal Susceptibility Testing of Yeasts, 1st ed.; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Dudiuk, C.; Berrio, I.; Leonardelli, F.; Morales-Lopez, S.; Theill, L.; Macedo, D.; Rodriguez, J.Y.; Salcedo, S.; Marin, A.; Gamarra, S.; et al. Antifungal activity and killing kinetics of anidulafungin, caspofungin and amphotericin B against Candida auris. J. Antimicrob. Chemother. 2019, 74, 2295–2302. [Google Scholar] [CrossRef] [PubMed]
- Poester, V.R.; Munhoz, L.S.; Benelli, J.L.; Melo, A.M.; Al-Hatmi, A.M.S.; Larwood, D.J.; Martinez, M.; Stevens, D.A.; Xavier, M.O. Initial Results of the International Efforts in Screening New Agents against Candida auris. J. Fungi 2022, 8, 771. [Google Scholar] [CrossRef]
- Sandovsky-Losica, H.; Shwartzman, R.; Lahat, Y.; Segal, E. Antifungal activity against Candida albicans of nikkomycin Z in combination with caspofungin, voriconazole or amphotericin B. J. Antimicrob. Chemother. 2008, 62, 635–637. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Arendrup, M.C.; Pfaller, M.A.; Bonfietti, B.; Bustamente, E.; Canton, E.; Cuenca, M.; Dannaoui, E.; Fothergill, A.; Fuller, J. Interlaboratory variability of caspofungin MICs for Candida spp. using CLSI and EUCAST methods: Should the clinical laboratory be testing this agent? Antimicrob. Agents Chemother. 2013, 57, 5836–5842. [Google Scholar] [CrossRef]
- Briano, F.; Magnasco, L.; Sepulcri, C.; Dettori, S.; Dentone, C.; Mikulska, M.; Ball, L.; Vena, A.; Robba, C.; Patroniti, N.; et al. Candida auris Candidemia in Critically Ill, Colonized Patients: Cumulative Incidence and Risk Factors. Infect. Dis. Ther. 2022, 11, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Alatoom, A.; Sartawi, M.; Lawlor, K.; AbdelWareth, L.; Thomsen, J.; Nusair, A.; Mirza, I. Persistent candidemia despite appropriate fungal therapy: First case of Candida auris from the United Arab Emirates. Int. J. Infect. Dis. 2018, 70, 36–37. [Google Scholar] [CrossRef]
- Betts, R.F.; Nucci, M.; Talwar, D.; Gareca, M.; Telles, F.; Bedimo, R.J.; Herbrecht, R. Caspofungin high-dose study group. A multicenter, double-blind trial of a high-dose caspofungin treatment regimen versus a standard caspofungin treatment regimen for adult patients with invasive candidiasis. Clin. Infect. Dis. 2009, 48, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Rotstein, C.M.F.; Betts, R.F.; Nucci, M.; Talwar, D.; Waele, J.J.; Vazquez, J.S.; Dupont, B.F.; Horn, D.L.; Reboli, A.C. Micafungin versus caspofungin for treatment of candidemia and other forms of invasive candidiasis. Clin. Infect. Dis. 2007, 45, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Caballero, U.; Eraso, E.; Quindós, G.; Jauregizar, N. In Vitro Interaction and Killing-Kinetics of Amphotericin B Combined with Anidulafungin or Caspofungin against Candida auris. Pharmaceutics 2021, 13, 1333. [Google Scholar] [CrossRef]
- Navarro-Arias, M.J.; Hernández-Chávez, M.J.; Garcia-Carnero, L.C.; Amezcua-Hernández, D.G.; E Lozoya-Pérez, N.; Estrada-Mata, E.; Martínez-Duncker, I.; Franco, B.; Mora-Montes, H.M. Differential recognition of Candida tropicalis, Candida guilliermondii, Candida krusei, and Candida auris by human innate immune cells. Infect. Drug Resist. 2019, 12, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Shahi, G.; Kumar, M.; Skwarecki, A.S.; Edmondson, M.; Banerjee, A.; Usher, J.; Gow, N.A.; Milewski, S.; Prasad, R. Fluconazole resistant Candida auris clinical isolates have increased levels of cell wall chitin and increased susceptibility to a glucosamine-6-phosphate synthase inhibitor. Cell Surf. 2022, 8, 100076. [Google Scholar] [CrossRef]
- Pezzotti, G.; Kobara, M.; Asai, T.; Nakaya, T.; Miyamoto, N.; Adachi, T.; Yamamoto, T.; Kanamura, N.; Ohgitani, E.; Marin, E.; et al. Raman Imaging of Pathogenic Candida auris: Visualization of Structural Characteristics and Machine-Learning Identification. Front. Microbiol. 2021, 12, 769597. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Paul, R.A.; Rudramurthy, S.M.; Kashyap, N.; Bhattacharya, S.; Soman, R.; Shankarnarayan, S.A.; Chavan, D.; Singh, S.; Das, P.; et al. Impact of FKS1 Genotype on Echinocandin In Vitro Susceptibility in Candida auris and In Vivo Response in a Murine Model of Infection. Antimicrob. Agents Chemother. 2022, 66, e0165221. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates Number | Clade | Body Site | FKS Mutation | MIC (mg/L) | ||
|---|---|---|---|---|---|---|
| ANI | MICA | NIK | ||||
| 20 | South Asian | Wound swab | HS1 WT HS2 R1354H | 0.25 | 0.25 | 2 |
| 27 (NCPF 8991) | South Asian | Pleural fluid | HS1 WT HS2 WT | 0.12 | 0.25 | >16 |
| 28 (NCPF 8992) | South Asian | Central line | HS1 S639Y HS2 WT | 4 | 4 | 16 |
| 196 | South Asian | Blood | HS1 WT HS2 WT | 0.12 | 0.25 | >16 |
| 208 | South Asian | Screening swab | HS1 S639P HS2 WT | 0.5 | 2 | 16 |
| Type strain (NCPF 13029 = CBS 10913) | East Asian | External ear | HS1 WT HS2 WT | 0.03 | 0.12 | >16 |
| 12372 (CBS 12372) | East Asian | Blood | HS1 WT HS2 WT | 0.03 | 0.03 | >16 |
| 12373 (CBS 12373) | East Asian | Blood | HS1 WT HS2 WT | 0.06 | 0.06 | 16 |
| 2 (NCPF 8977) | South African | CSF | HS1 WT HS2 WT | 0.03 | 0.25 | 8 |
| 204 | South African | Tracheostomy | HS1 WT HS2 WT | 0.03 | 0.12 | >16 |
| 206 | South African | Blood | HS1 WT HS2 WT | 0.06 | 0.25 | 4 |
| I-24 | South American (Israel) | Blood | HS1 WT HS2 WT | 0.06 | 0.12 | 4 |
| I-156 | South American (Israel) | Blood | HS1 WT HS2 WT | 0.06 | 0.12 | 4 |
| 13108 (CDC B-13108) | South American (Colombia) | Hospital environment | HS1 WT HS2 WT | 0.06 | 0.12 | 4 |
| 16565 (CDC B-16565) | South American (Colombia) | Hospital environment | HS1 WT HS2 WT | 0.015 | 0.06 | 8 |
| Clade | Isolate Number | Maximum Log Decreases/Increases in CFU in Time–Killing Experiments at the Indicated Anidulafungin and Anidulafungin + Nikkomycin Z Concentrations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Anidulafungin (mg/L) | Anidulafungin (mg/L) + Nikkomycin Z (mg/L) | ||||||||||
| 0.25 | 1 | 8 | 16 | 32 | 0.25 + 8 | 1 + 8 | 8 + 8 | 16 + 8 | 32 + 8 | ||
| South Asian | 20 | −0.27 * | −0.22 * | −0.29 * | −0.88 | −0.74 | −0.69 | −1.0 | −1.57 | −1.48 | −1.57 |
| 27 | −0.61 | −1.04 | −1.4 | −1.32 | −1.4 | −2.3 | −2.78 | −3.16 | −3.22 | −3.05 | |
| 28 | +2.39 | −0.22 * | −0.19 * | +0.88 | +0.54 | +1.42 | −0.05 * | −0.10 * | −0.10 * | −0.08 * | |
| 196 | −0.39 | −0.57 | −0.42 | −0.62 | −0.96 | −2.54 | −3.02 | −3.02 | −3.02 | −3.02 | |
| 208 | −0.08 * | +1.24 | +1.22 | +0.82 | +0.52 | +1.40 | +1.10 | −0.15 * | −0.10 * | −0.43 | |
| East Asian | Type strain | −1.15 * | −1.54 * | −1.98 | −1.98 | −3.0 | −1.0 | −1.13 | −2.13 | −3.9 | −3.9 |
| 12372 | −0.58 * | −0.54 * | −1.42 * | −1.18 * | −1.23 * | −0.87 * | −1.92 | −3.85 | −3.85 | −3.85 | |
| 12373 | −0.52 * | −0.62 * | −0.62 * | −0.70 * | −0.80 * | −1.0 | −2.22 | −4.0 | −4.0 | −4.0 | |
| South African | 2 | +0.45 | +0.24 | +0.26 | +0.32 | +0.26 | −1.28 | −1.30 | −1.41 | −1.48 | −1.60 |
| 204 | −0.35 * | −0.03 * | −0.19 * | +0.33 | −0.15 * | −0.83 | −0.95 | −1.20 | −1.32 | −1.82 | |
| 206 | +0.48 | +0.51 | +0.54 | −0.19 * | −0.92 * | −0.34 * | −0.21 * | −0.22 * | −0.29 * | −0.44 * | |
| South American (from Israel) | I-24 | −0.33 * | −0.27 * | −0.22 * | −0.19 * | −0.11 * | −0.57 | −0.69 | −0.77 | −0.82 | −1.32 |
| I-156 | −0.35 * | −0.18 * | −0–14 * | −0.22 * | −0–15 * | −0.85 | −1.22 | −1.41 | −1.71 | −2.26 | |
| South American (from Colombia) | 13108 | −0.96 | −0.77 | −0.78 | −0.78 | −0.48 | −4.08 | −4.08 | −4.08 | −4.08 | −4.08 |
| 16565 | −2.3 | −1.7 | −1.09 | −0.79 | −0.99 | −4.15 | −4.15 | −4.15 | −4.15 | −4.15 | |
| Clade | Isolate Number | Maximum Log Decreases/Increases in CFU in Time–Killing Experiments at the Indicated Micafungin and Micafungin + Nikkomycin Z Concentrations | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Micafungin (mg/L) | Micafungin (mg/L) +Nikkomycin Z (mg/L) | ||||||||||
| 0.25 | 1 | 8 | 16 | 32 | 0.25 + 8 | 1 + 8 | 8 + 8 | 16 + 8 | 32 + 8 | ||
| South Asian | 20 | +1.02 | −0.47 * | −0.35 * | −0.52 * | −0.44 * | −1.78 | −1.68 | −1.13 | −0.95 | −1.38 * |
| 27 | −1.68 | −2.51 | −1.43 | −1.27 | −1.30 | −2.38 | −2.40 | −2.51 | −2.84 | −3.6 | |
| 28 | +1.45 | +1.34 | −0.83 * | −0.35 * | −0.44 * | +1.52 | −0.16 * | −0.70 * | −0.57 * | −0.52 * | |
| 196 | −0.51 | −0.69 | −0.32 | −0.24 | −0.24 | −2.32 | −2.15 | −2.62 | −2.62 | −2.72 | |
| 208 | +1.67 | +1.89 | −0.08 * | −0.14 * | +1.13 | +1.72 | +1.52 | −0–04 * | −0.04 * | −0.13 * | |
| East Asian | Type strain | −0.67 * | −0.60 | −0.56 | −0.49 * | −0.51 * | −0.69 | −0.64 | −0.64 | −0.54 | −0.62 |
| 12372 | −0.54 * | −0.67 * | −0.54 * | −0.64 * | −0.72 * | −1.51 | −1.90 | −2.12 | −2.54 | −3.15 | |
| 12373 | −0.30 * | −0.39 * | −0.52 * | −0.33 * | −0.39 * | −1.47 | −1.49 | −1.70 | −1.74 | −2.64 | |
| South African | 2 | +0.37 | +0.34 | +0.26 | +0.36 | +0.21 | −1.46 | −1.50 | −1.60 | −1.67 | −2.48 |
| 204 | −0.08 * | −0.11 * | −0.05 * | −0.05 * | −0.11 * | −0.56 | −0.65 | −0.65 | −0.78 | −0.88 | |
| 206 | +0.54 | +0.51 | +0.60 | +0.48 | +0.48 | −0.23 * | −0.23 * | −0.23 * | −0.16 * | −0.22 * | |
| South American (from Israel) | I−24 | −0.30 * | −0.24 * | −0.19 * | −0.15 * | −0.24 * | −0.65 | −0.83 | −0.69 * | −0.83 | −0.79 |
| I−156 | −0.18 * | −0.30 * | −0.35 * | −0.35 * | −0.30 * | −1.30 | −1.52 | −1.49 | −1.48 | −1.42 | |
| South American (from Colombia) | 13108 | −0.86 * | −0.86 * | −0.95 * | −1.08 * | −1.08 | −3.13 | −3.35 | −4.26 | −4.26 | −4.26 |
| 16565 | −1.15 | −1.15 | −0.79 * | −0.59 * | −0.77 * | −4.26 | −4.26 | −4.26 | −4.26 | −4.26 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adnan, A.; Borman, A.M.; Tóth, Z.; Forgács, L.; Kovács, R.; Balázsi, D.; Balázs, B.; Udvarhelyi, G.; Kardos, G.; Majoros, L. In Vitro Killing Activities of Anidulafungin and Micafungin with and without Nikkomycin Z against Four Candida auris Clades. Pharmaceutics 2023, 15, 1365. https://doi.org/10.3390/pharmaceutics15051365
Adnan A, Borman AM, Tóth Z, Forgács L, Kovács R, Balázsi D, Balázs B, Udvarhelyi G, Kardos G, Majoros L. In Vitro Killing Activities of Anidulafungin and Micafungin with and without Nikkomycin Z against Four Candida auris Clades. Pharmaceutics. 2023; 15(5):1365. https://doi.org/10.3390/pharmaceutics15051365
Chicago/Turabian StyleAdnan, Awid, Andrew M. Borman, Zoltán Tóth, Lajos Forgács, Renátó Kovács, Dávid Balázsi, Bence Balázs, Gergely Udvarhelyi, Gábor Kardos, and László Majoros. 2023. "In Vitro Killing Activities of Anidulafungin and Micafungin with and without Nikkomycin Z against Four Candida auris Clades" Pharmaceutics 15, no. 5: 1365. https://doi.org/10.3390/pharmaceutics15051365
APA StyleAdnan, A., Borman, A. M., Tóth, Z., Forgács, L., Kovács, R., Balázsi, D., Balázs, B., Udvarhelyi, G., Kardos, G., & Majoros, L. (2023). In Vitro Killing Activities of Anidulafungin and Micafungin with and without Nikkomycin Z against Four Candida auris Clades. Pharmaceutics, 15(5), 1365. https://doi.org/10.3390/pharmaceutics15051365