
The Antibiotic Kitasamycin—A Potential Agent for Specific Fibrosis Preventing Therapy after Fistulating Glaucoma Surgery?
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Dose–Response Testing of Kitasamycin
2.3. Evaluation of the Antifibrotic Effect of Kitasamycin
2.3.1. Immunofluorescence
2.3.2. Western Blot Analysis
3. Results
3.1. Dose–Response Testing after In Vitro Incubation of hTFs with Kitasamycin
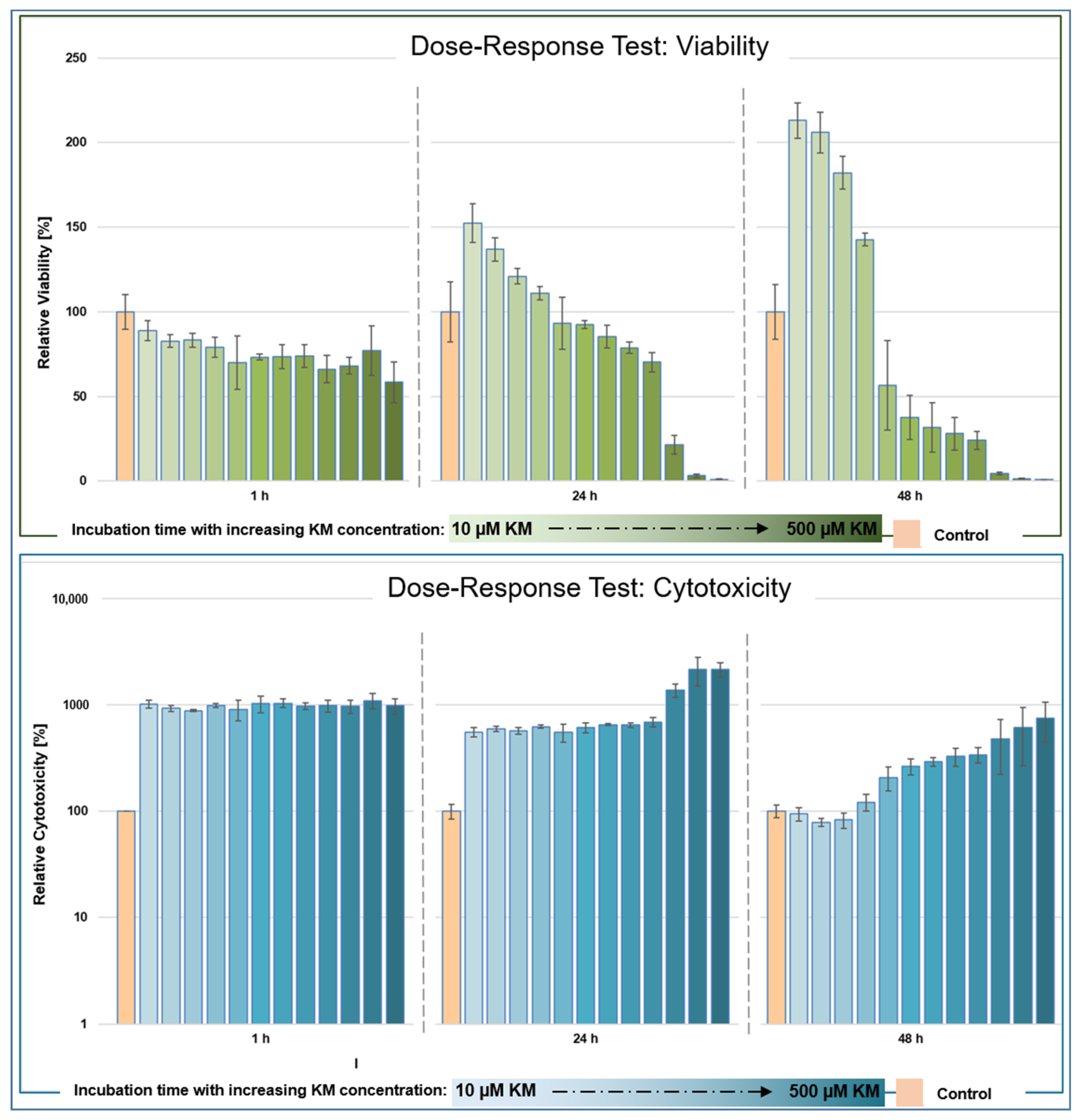
3.1.1. Viability
3.1.2. Cytotoxicity
3.2. In Vitro Evaluation of the Antifibrotic Effect of Kitasamycin in TGF-β1-Mediated Fibrotic Cell Culture Model
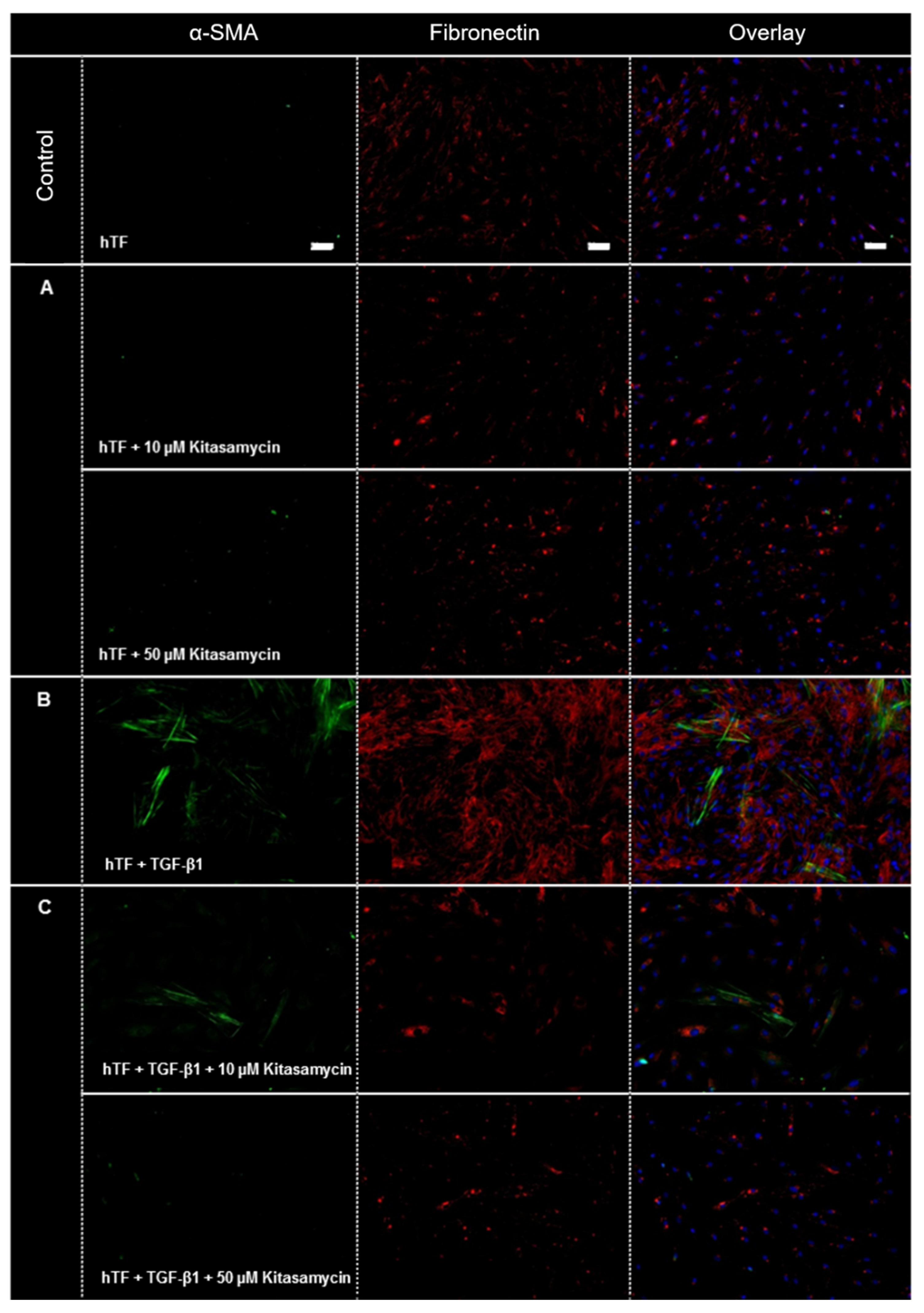
3.2.1. Immunofluorescence
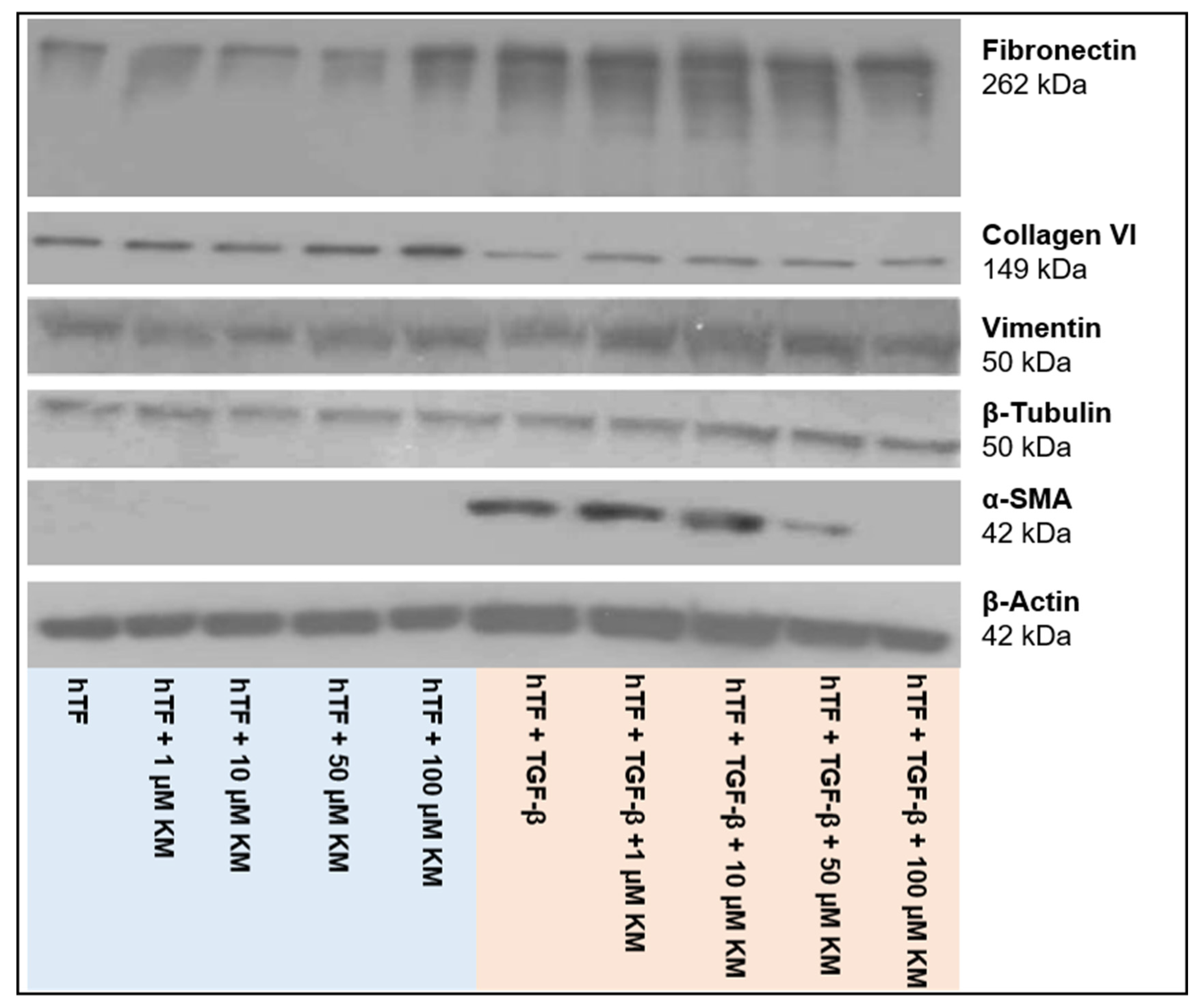
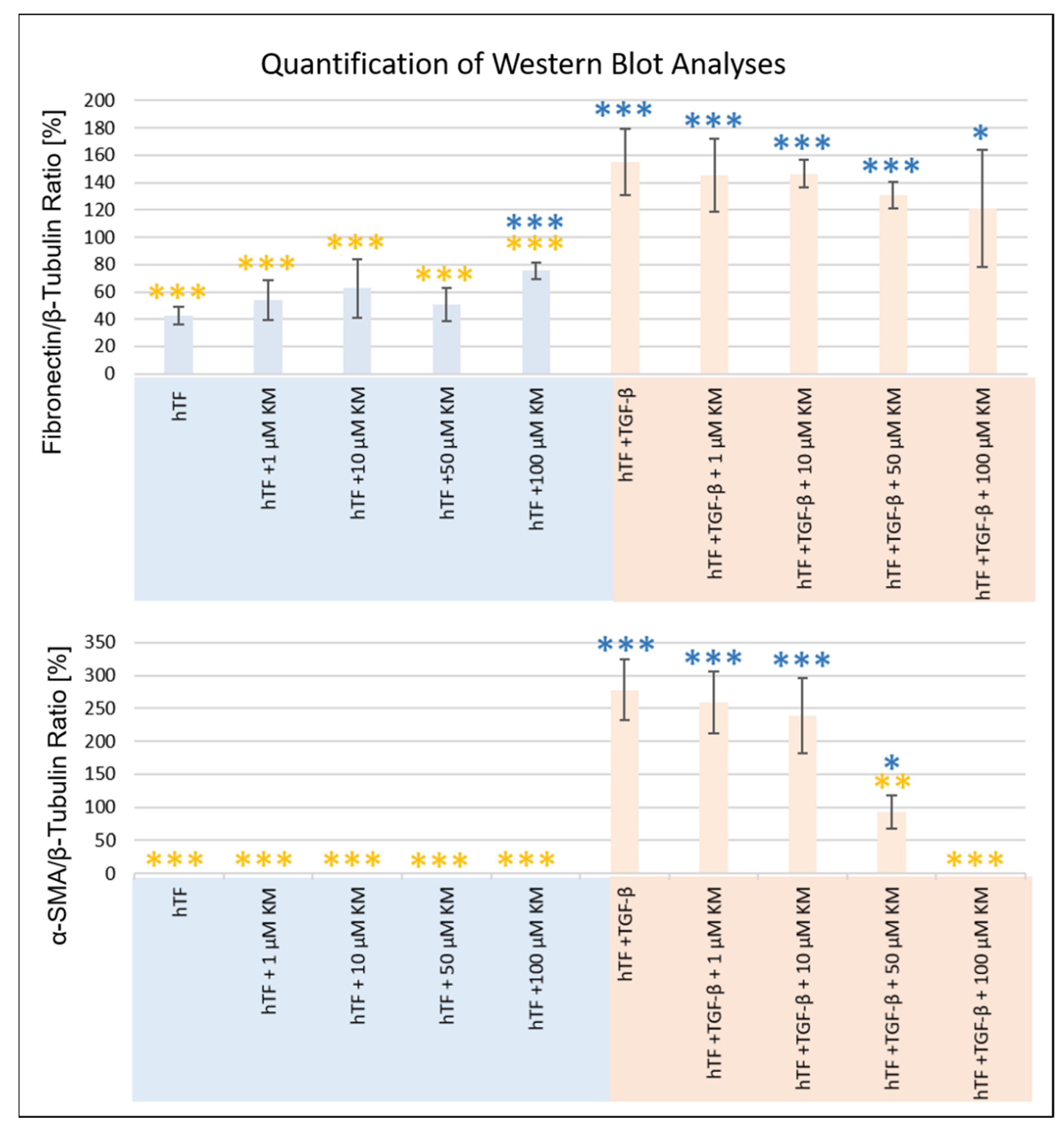
3.2.2. Western Blot Analysis
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Report on Vision. Available online: https://www.who.int/publications/i/item/world-report-on-vision (accessed on 19 January 2021).
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global Prevalence of Glaucoma and Projections of Glaucoma Burden through 2040: A Systematic Review and Meta-Analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Neuroprotection in Glaucoma—EyeWiki. Available online: https://eyewiki.aao.org/Neuroprotection_in_Glaucoma (accessed on 4 January 2023).
- Kuo, C.-Y.; Liu, C.J.-L. Neuroprotection in Glaucoma: Basic Aspects and Clinical Relevance. J. Pers. Med. 2022, 12, 1884. [Google Scholar] [CrossRef] [PubMed]
- Sena, D.F.; Ramchand, K.; Lindsley, K. Neuroprotection for Treatment of Glaucoma in Adults. Cochrane Database Syst. Rev. 2017, 1, CD006539. [Google Scholar] [CrossRef] [PubMed]
- Komáromy, A.M.; Koehl, K.L.; Park, S.A. Looking into the Future: Gene and Cell Therapies for Glaucoma. Veter Ophthalmol. 2021, 24, 16–33. [Google Scholar] [CrossRef]
- Baudouin, C.; Labbé, A.; Liang, H.; Pauly, A.; Brignole-Baudouin, F. Preservatives in Eyedrops: The Good, the Bad and the Ugly. Prog. Retin. Eye Res. 2010, 29, 312–334. [Google Scholar] [CrossRef]
- Frech, S.; Kreft, D.; Guthoff, R.F.; Doblhammer, G. Pharmacoepidemiological Assessment of Adherence and Influencing Co-Factors among Primary Open-Angle Glaucoma Patients—An Observational Cohort Study. PLoS ONE 2018, 13, e0191185. [Google Scholar] [CrossRef]
- Miller, P.E.; Eaton, J.S. Medical Anti-Glaucoma Therapy: Beyond the Drop. Veter Ophthalmol. 2020, 24, 2–15. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, C.M.; Ferreira, J.d.L.M. Overview of Cicatricial Modulators in Glaucoma Fistulizing Surgery. Int. Ophthalmol. 2020, 40, 2789–2796. [Google Scholar] [CrossRef]
- Hoffmann, E.M.; Hengerer, F.; Klabe, K.; Schargus, M.; Thieme, H.; Voykov, B. Glaucoma Surgery Today. Ophthalmologe 2021, 118, 239–247. [Google Scholar] [CrossRef]
- Molteno, A.C. New Implant for Drainage in Glaucoma. Animal Trial. Br. J. Ophthalmol. 1969, 53, 161–168. [Google Scholar] [CrossRef]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef] [PubMed]
- Pillunat, L.E.; Erb, C.; Jünemann, A.G.M.; Kimmich, F. Micro-Invasive Glaucoma Surgery (MIGS): A Review of Surgical Procedures Using Stents. Clin. Ophthalmol. 2017, 11, 1583–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlunck, G.; Meyer-ter-Vehn, T.; Klink, T.; Grehn, F. Conjunctival Fibrosis Following Filtering Glaucoma Surgery. Exp. Eye. Res. 2016, 142, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, O.; Kitano-Izutani, A.; Tomoyose, K.; Reinach, P.S. Pathobiology of Wound Healing after Glaucoma Filtration Surgery. BMC Ophthalmol. 2015, 15, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holló, G. Wound Healing and Glaucoma Surgery: Modulating the Scarring Process with Conventional Antimetabolites and New Molecules. Dev. Ophthalmol. 2017, 59, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Wolters, J.E.J.; van Mechelen, R.J.S.; Al Majidi, R.; Pinchuk, L.; Webers, C.A.B.; Beckers, H.J.M.; Gorgels, T.G.M.F. History, Presence, and Future of Mitomycin C in Glaucoma Filtration Surgery. Curr. Opin. Ophthalmol. 2021, 32, 148–159. [Google Scholar] [CrossRef]
- Stahnke, T.; Gajda-Deryło, B.; Jünemann, A.G.; Stachs, O.; Sterenczak, K.A.; Rejdak, R.; Beck, J.; Schütz, E.; Möller, S.; Barrantes, I.; et al. Suppression of the TGF-β Pathway by a Macrolide Antibiotic Decreases Fibrotic Responses by Ocular Fibroblasts in Vitro. R. Soc. Open Sci. 2020, 7, 200441. [Google Scholar] [CrossRef]
- Stahnke, T.; Gajda-Derylo, B.; Stachs, O.; Guthoff, R.F.; Jünemann, A.; Füllen, G. Systems Biology in Ophthalmology—Innovative Drug Identification for the Specific Prevention of Postoperative Fibrosis. Klin. Monbl. Augenheilkd. 2019, 236, 1428–1434. [Google Scholar] [CrossRef]
- Vezina, C.; Bolduc, C.; Kudelski, A.; Audet, P. Biosynthesis of Kitasamycin (Leucomycin) by Leucine Analog-Resistant Mutants of Streptomyces Kitasatoensis. Antimicrob. Agents Chemother. 1979, 15, 738–746. [Google Scholar] [CrossRef] [Green Version]
- KITASAMYCIN. Available online: https://drugs.ncats.io/drug/G986N7X02W#general (accessed on 4 January 2023).
- Sterenczak, K.A.; Barrantes, I.; Stahnke, T.; Stachs, O.; Fuellen, G.; Undre, N. Co-Infections: Testing Macrolides for Added Benefit in Patients with COVID-19. Lancet Microbe 2020, 1, e313. [Google Scholar] [CrossRef]
- Wada, M.; Nakayama, T.; Ishizaki, T.; Satoh, T.; Ikeda, S. Practice Patterns for Lower Respiratory Tract Infections in Hospital Patients with Particular Focus on Bacteriological Examinations and Injection Antibiotics Use. Int. J. Gen. Med. 2013, 6, 597–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Castellanos, E.P.; Gallardo, S.G.; Sanchez-Betancourt, J.I.; Ortega, M.E.T.; Guerrero, D.Q.; Carrasco, A.C.; Elvira, S.E.M. Effect Found in the Combination of Kitasamycin with Other Antibiotics Used against Actinobacillus Pleuropneumoniae. J. Biosci. Med. 2020, 8, 63–75. [Google Scholar] [CrossRef]
- Stahnke, T.; Löbler, M.; Kastner, C.; Stachs, O.; Wree, A.; Sternberg, K.; Schmitz, K.P.; Guthoff, R. Different Fibroblast Subpopulations of the Eye: A Therapeutic Target to Prevent Postoperative Fibrosis in Glaucoma Therapy. Exp. Eye. Res. 2012, 100, 88–97. [Google Scholar] [CrossRef]
- Promega Corporation. RealTime-GloTM MT Cell Viability Assay Instructions for Use of Products G9711, G9712 and G9713; Promega: Madison, WI, USA, 2021. [Google Scholar]
- Promega Corporation. CellToxTM Green Cytotoxicity Assay Instructions for Use of Products G8741, G8742, G8743 and G8731; Promega: Madison, WI, USA, 2020. [Google Scholar]
- Michie, C. Anti-Inflammatory Activity of Macrolide Antibiotics. Trends Mol. Med. 2002, 8, 547. [Google Scholar] [CrossRef]
- Arsic, B.; Barber, J.; Čikoš, A.; Mladenovic, M.; Stankovic, N.; Novak, P. 16-Membered Macrolide Antibiotics: A Review. Int. J. Antimicrob. Agents 2018, 51, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Ökesli-Armlovich, A.; Morgens, D.; Bassik, M.C.; Khosla, C. A Genome-Wide Analysis of Targets of Macrolide Antibiotics in Mammalian Cells. J. Biol. Chem. 2020, 295, 2057–2067. [Google Scholar] [CrossRef]
- Zada, M.; Pattamatta, U.; White, A. Modulation of Fibroblasts in Conjunctival Wound Healing. Ophthalmology 2018, 125, 179–192. [Google Scholar] [CrossRef]
- Van de Velde, S.; Van Bergen, T.; Vandewalle, E.; Moons, L.; Stalmans, I. Modulation of Wound Healing in Glaucoma Surgery. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2015; Volume 221, pp. 319–340. [Google Scholar]
- Wimmer, I.; Grehn, F. Control of Wound Healing after Glaucoma Surgery. Effect and Inhibition of the Growth Factor TGF-Beta. Ophthalmologe 2002, 99, 678–682. [Google Scholar] [CrossRef]
- Walker, J.L.; Bleaken, B.M.; Romisher, A.R.; Alnwibit, A.A.; Menko, A.S. In Wound Repair Vimentin Mediates the Transition of Mesenchymal Leader Cells to a Myofibroblast Phenotype. Mol. Biol. Cell 2018, 29, 1555–1570. [Google Scholar] [CrossRef]
- Torr, E.E.; Ngam, C.R.; Bernau, K.; Tomasini-Johansson, B.; Acton, B.; Sandbo, N. Myofibroblasts Exhibit Enhanced Fibronectin Assembly That Is Intrinsic to Their Contractile Phenotype. J. Biol. Chem. 2015, 290, 6951–6961. [Google Scholar] [CrossRef]
- Baum, J.; Duffy, H.S. Fibroblasts and Myofibroblasts: What Are We Talking About? J. Cardiovasc. Pharm. 2011, 57, 376–379. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| t-Test Analysis (Two-Sample t-Test) | Fibronectin | Collagen VI | Vimentin | α-SMA | β-Actin | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| p-Value | Level | p-Value | Level | p-Value | Level | p-Value | Level | p-Value | Level | ||
| hTF | hTF + 1 µM KM | 0.183 | n.s | 0.178 | n.s | 0.085 | n.s | n.d | - | 0.835 | n.s |
| hTF + 10 µM KM | 0.111 | n.s | 0.117 | n.s | 0.046 | * | n.d | - | 0.153 | n.s | |
| hTF + 50 µM KM | 0.431 | n.s | 0.004 | ** | 0.141 | n.s | n.d | - | 0.200 | n.s | |
| hTF + 100 µM KM | 0.000 | *** | 0.234 | n.s | 0.637 | n.s | n.d | - | 0.523 | n.s | |
| hTF + TGF-β1 | 0.000 | *** | 0.090 | n.s | 0.915 | n.s | 0.000 | *** | 0.027 | * | |
| hTF + TGF-β1 + 1 µM KM | 0.000 | *** | 0.121 | n.s | 0.466 | n.s | 0.000 | *** | 0.252 | n.s | |
| hTF + TGF-β1 + 10 µM KM | 0.000 | *** | 0.122 | n.s | 0.006 | ** | 0.000 | *** | 0.127 | n.s | |
| hTF + TGF-β1 + 50 µM KM | 0.001 | *** | 0.056 | * | 0.031 | * | 0.035 | * | 0.321 | n.s | |
| hTF + TGF-β1 + 100 µM KM | 0.050 | * | 0.037 | * | 0.145 | n.s | n.d | - | 0.154 | n.s | |
| hTF + TGF-β1 | hTF | 0.000 | *** | 0.900 | n.s | 0.915 | n.s | 0.000 | *** | 0.027 | n.s |
| hTF + 1 µM KM | 0.000 | *** | 0.013 | ** | 0.069 | n.s | 0.000 | *** | 0.017 | * | |
| hTF + 10 µM KM | 0.000 | *** | 0.015 | ** | 0.054 | * | 0.000 | *** | 0.514 | n.s | |
| hTF + 50 µM KM | 0.000 | *** | 0.001 | *** | 0.149 | n.s | 0.000 | *** | 0.231 | n.s | |
| hTF + 100 µM KM | 0.001 | *** | 0.077 | n.s | 0.571 | n.s | 0.000 | *** | 0.471 | n.s | |
| hTF + TGF-β1 + 1 µM KM | 0.616 | n.s | 0.659 | n.s | 0.414 | n.s | 0.584 | n.s | 0.213 | n.s | |
| hTF + TGF-β1 + 10 µM KM | 0.533 | n.s | 0.614 | n.s | 0.005 | ** | 0.312 | n.s | 0.107 | n.s | |
| hTF + TGF-β1 + 50 µM KM | 0.131 | n.s | 0.695 | n.s | 0.028 | * | 0.001 | ** | 0.518 | n.s | |
| hTF + TGF-β1 + 100 µM KM | 0.290 | n.s | 1.000 | n.s | 0.131 | n.s | 0.000 | *** | 0.852 | n.s | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sterenczak, K.A.; Fuellen, G.; Jünemann, A.; Guthoff, R.F.; Stachs, O.; Stahnke, T. The Antibiotic Kitasamycin—A Potential Agent for Specific Fibrosis Preventing Therapy after Fistulating Glaucoma Surgery? Pharmaceutics 2023, 15, 329. https://doi.org/10.3390/pharmaceutics15020329
Sterenczak KA, Fuellen G, Jünemann A, Guthoff RF, Stachs O, Stahnke T. The Antibiotic Kitasamycin—A Potential Agent for Specific Fibrosis Preventing Therapy after Fistulating Glaucoma Surgery? Pharmaceutics. 2023; 15(2):329. https://doi.org/10.3390/pharmaceutics15020329
Chicago/Turabian StyleSterenczak, Katharina A., Georg Fuellen, Anselm Jünemann, Rudolf F. Guthoff, Oliver Stachs, and Thomas Stahnke. 2023. "The Antibiotic Kitasamycin—A Potential Agent for Specific Fibrosis Preventing Therapy after Fistulating Glaucoma Surgery?" Pharmaceutics 15, no. 2: 329. https://doi.org/10.3390/pharmaceutics15020329
APA StyleSterenczak, K. A., Fuellen, G., Jünemann, A., Guthoff, R. F., Stachs, O., & Stahnke, T. (2023). The Antibiotic Kitasamycin—A Potential Agent for Specific Fibrosis Preventing Therapy after Fistulating Glaucoma Surgery? Pharmaceutics, 15(2), 329. https://doi.org/10.3390/pharmaceutics15020329