
Neuroprotective–Neurorestorative Effects Induced by Progesterone on Global Cerebral Ischemia: A Narrative Review

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
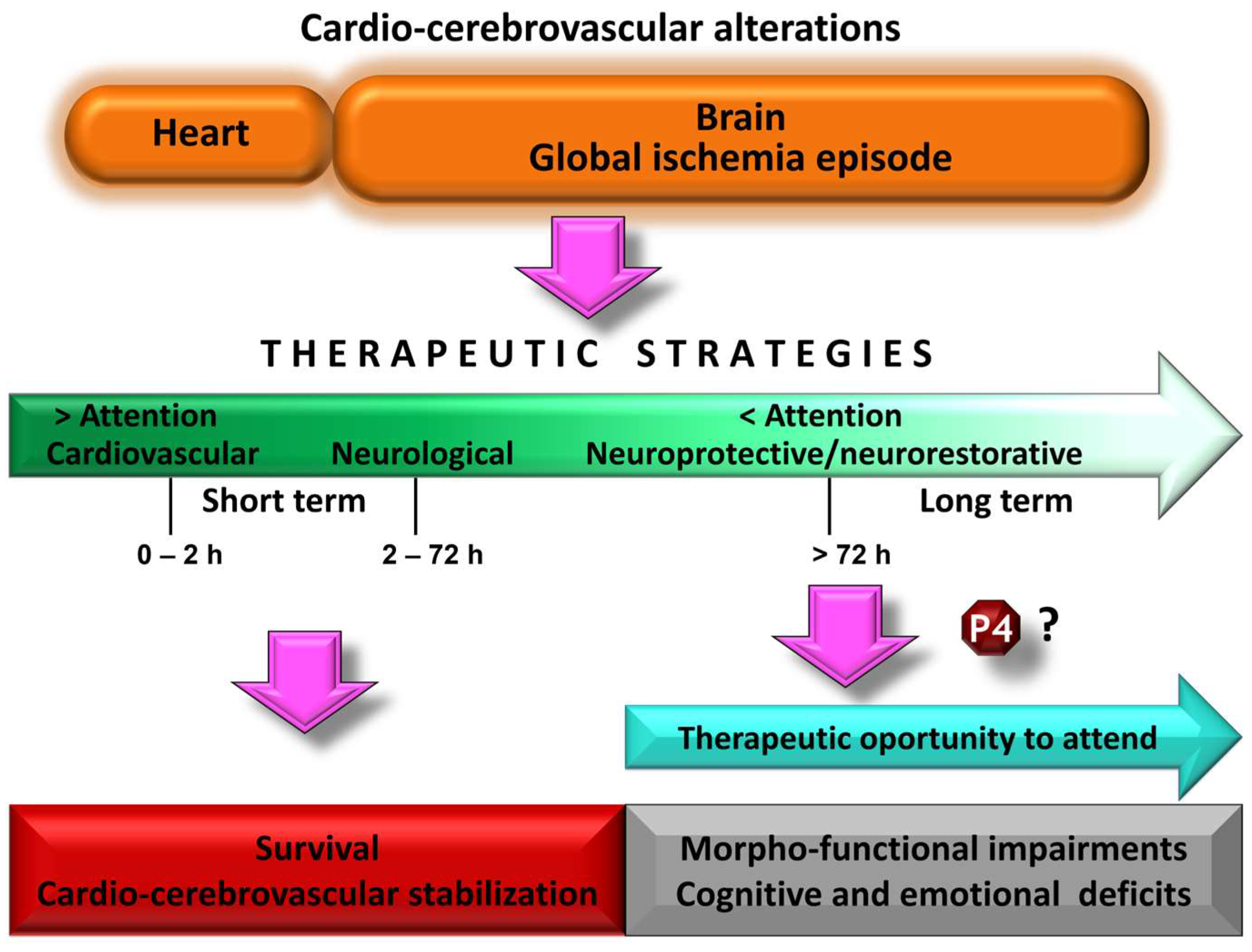
2. Pathophysiology of Global Cerebral Ischemia and Morphofunctional Changes
3. Guide Recommendations for the Study of Potential Treatments against Global Cerebral Ischemia
4. Progesterone as a Neuroactive Steroid and Its Mechanisms of Action
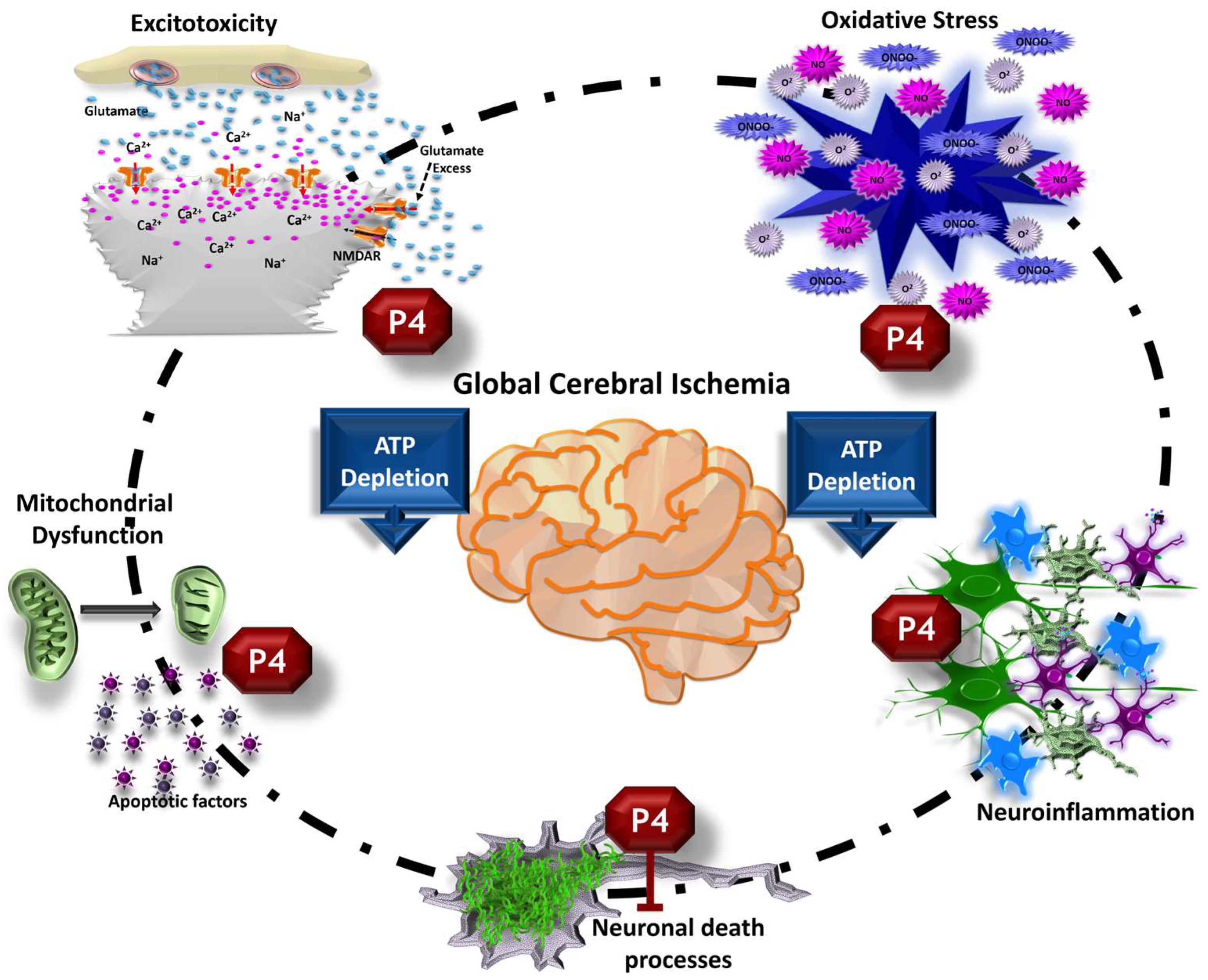
5. Biochemical and Cellular Effects Supporting Progesterone as a Treatment against Global Cerebral Ischemia
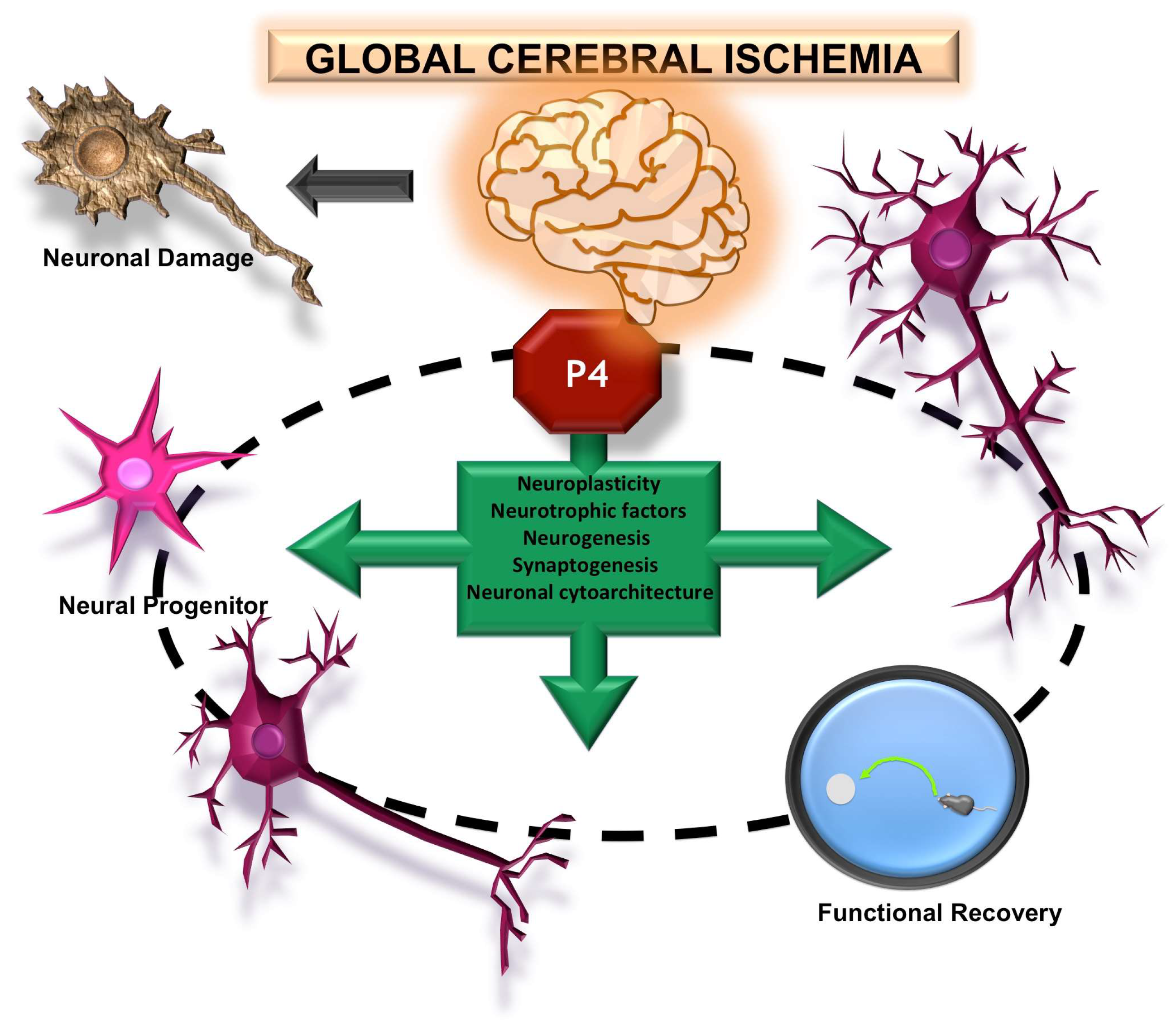
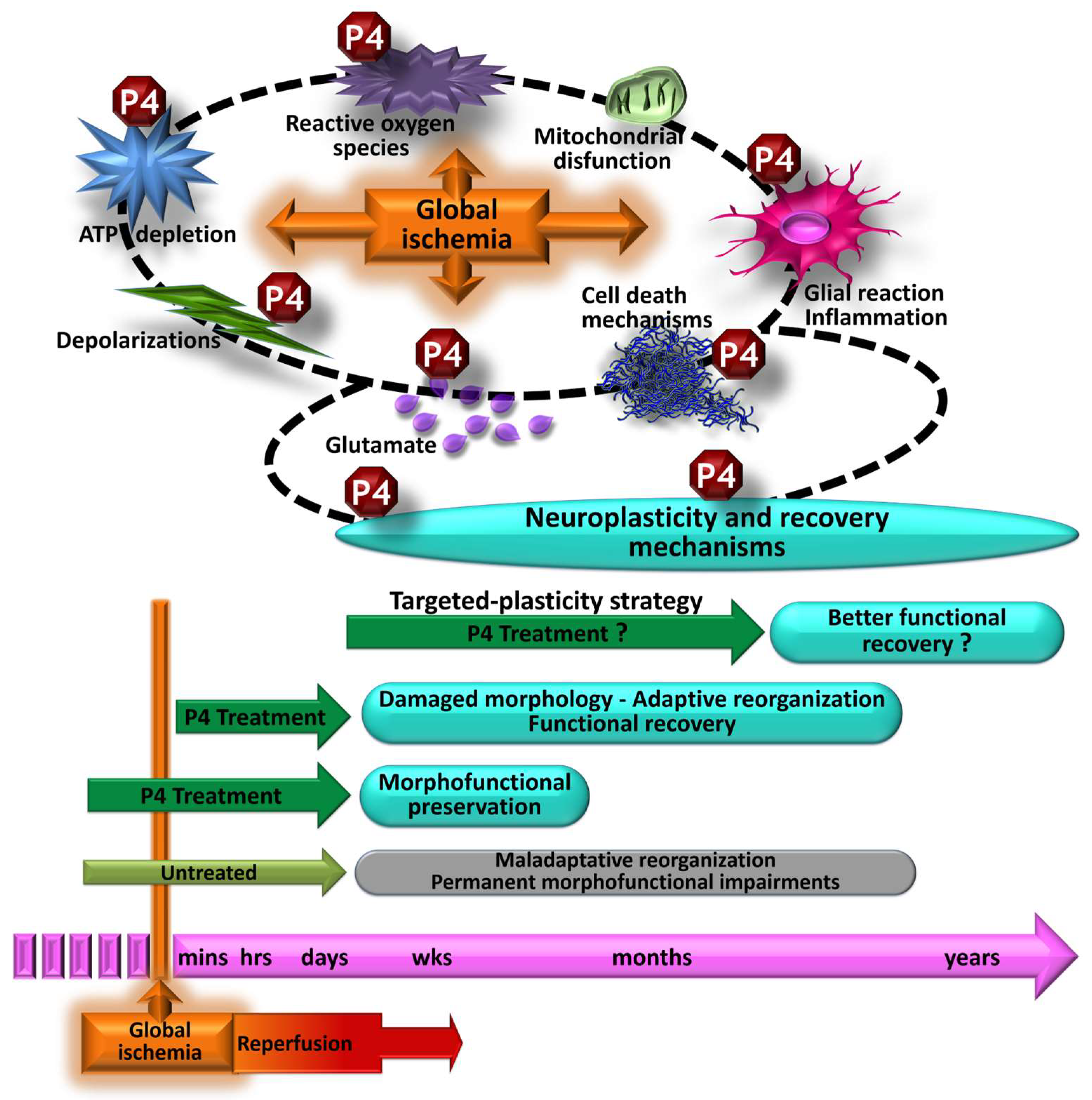
6. Morphological and Functional Effects Induced by Progesterone on Global Cerebral Ischemia
7. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 21 November 2023).
- Perez, C.A.; Samudra, N.; Aiyagari, V. Cognitive and Functional Consequence of Cardiac Arrest. Curr. Neurol. Neurosci. Rep. 2016, 16, 70. [Google Scholar] [CrossRef] [PubMed]
- Elmer, J.; Callaway, C.W. The Brain after Cardiac Arrest. Semin. Neurol. 2017, 37, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, G.; Ihle-Hansen, H.; Sandset, E.C.; Jacobsen, D.; Wimmer, H.; Ihle-Hansen, H. Long Term Cognitive Function after Cardiac Arrest: A Mini-Review. Front. Aging Neurosci. 2022, 14, 885226. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Lana, S.; Marquie, M.; Ruiz, A.; Boada, M. Cognitive and Neuropsychiatric Manifestations of COVID-19 and Effects on Elderly Individuals With Dementia. Front. Aging Neurosci. 2020, 12, 588872. [Google Scholar] [CrossRef] [PubMed]
- Dou, Q.; Wei, X.; Zhou, K.; Yang, S.; Jia, P. Cardiovascular Manifestations and Mechanisms in Patients with COVID-19. Trends Endocrinol. Metab. 2020, 31, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, L.; Yue, L.; Xiao, S. Alzheimer’s disease and COVID-19: Interactions, intrinsic linkages, and the role of immunoinflammatory responses in this process. Front. Immunol. 2023, 14, 1120495. [Google Scholar] [CrossRef] [PubMed]
- Berg, K.M.; Soar, J.; Andersen, L.W.; Bottiger, B.W.; Cacciola, S.; Callaway, C.W.; Couper, K.; Cronberg, T.; D’Arrigo, S.; Deakin, C.D.; et al. Adult Advanced Life Support: 2020 International Consensus on Cardiopulmonary Resuscitation and Emergency Cardiovascular Care Science with Treatment Recommendations. Circulation 2020, 142, S92–S139. [Google Scholar] [CrossRef]
- Nolan, J.P.; Sandroni, C.; Bottiger, B.W.; Cariou, A.; Cronberg, T.; Friberg, H.; Genbrugge, C.; Haywood, K.; Lilja, G.; Moulaert, V.R.M.; et al. European Resuscitation Council and European Society of Intensive Care Medicine guidelines 2021: Post-resuscitation care. Intensive Care Med. 2021, 47, 369–421. [Google Scholar] [CrossRef]
- Mangus, D.B.; Huang, L.; Applegate, P.M.; Gatling, J.W.; Zhang, J.; Applegate, R.L., 2nd. A systematic review of neuroprotective strategies after cardiac arrest: From bench to bedside (Part I—Protection via specific pathways). Med. Gas. Res. 2014, 4, 9. [Google Scholar] [CrossRef]
- Choudhary, R.C.; Shoaib, M.; Sohnen, S.; Rolston, D.M.; Jafari, D.; Miyara, S.J.; Hayashida, K.; Molmenti, E.P.; Kim, J.; Becker, L.B. Pharmacological Approach for Neuroprotection after Cardiac Arrest—A Narrative Review of Current Therapies and Future Neuroprotective Cocktail. Front. Med. 2021, 8, 636651. [Google Scholar] [CrossRef]
- Giatti, S.; Garcia-Segura, L.M.; Barreto, G.E.; Melcangi, R.C. Neuroactive steroids, neurosteroidogenesis and sex. Prog. Neurobiol. 2019, 176, 1–17. [Google Scholar] [CrossRef]
- Guennoun, R.; Labombarda, F.; Gonzalez Deniselle, M.C.; Liere, P.; De Nicola, A.F.; Schumacher, M. Progesterone and allopregnanolone in the central nervous system: Response to injury and implication for neuroprotection. J. Steroid Biochem. Mol. Biol. 2015, 146, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.; Mattern, C.; Ghoumari, A.; Oudinet, J.P.; Liere, P.; Labombarda, F.; Sitruk-Ware, R.; De Nicola, A.F.; Guennoun, R. Revisiting the roles of progesterone and allopregnanolone in the nervous system: Resurgence of the progesterone receptors. Prog. Neurobiol. 2014, 113, 6–39. [Google Scholar] [CrossRef] [PubMed]
- Cespedes Rubio, A.E.; Perez-Alvarez, M.J.; Lapuente Chala, C.; Wandosell, F. Sex steroid hormones as neuroprotective elements in ischemia models. J. Endocrinol. 2018, 237, R65–R81. [Google Scholar] [CrossRef]
- Guennoun, R. Progesterone in the Brain: Hormone, Neurosteroid and Neuroprotectant. Int. J. Mol. Sci. 2020, 21, 5271. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.T.; Cohan, C.H.; Dave, K.R.; Wright, C.B.; Perez-Pinzon, M.A. Global cerebral ischemia: Synaptic and cognitive dysfunction. Curr. Drug Targets 2013, 14, 20–35. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, M.S.; Ainslie, P.N.; Griesdale, D.E. Clinical pathophysiology of hypoxic ischemic brain injury after cardiac arrest: A “two-hit” model. Crit. Care 2017, 21, 90. [Google Scholar] [CrossRef]
- van den Brule, J.M.D.; van der Hoeven, J.G.; Hoedemaekers, C.W.E. Cerebral Perfusion and Cerebral Autoregulation after Cardiac Arrest. Biomed. Res. Int. 2018, 2018, 4143636. [Google Scholar] [CrossRef]
- Iordanova, B.; Li, L.; Clark, R.S.B.; Manole, M.D. Alterations in Cerebral Blood Flow after Resuscitation from Cardiac Arrest. Front. Pediatr. 2017, 5, 174. [Google Scholar] [CrossRef]
- Sanganalmath, S.K.; Gopal, P.; Parker, J.R.; Downs, R.K.; Parker, J.C., Jr.; Dawn, B. Global cerebral ischemia due to circulatory arrest: Insights into cellular pathophysiology and diagnostic modalities. Mol. Cell Biochem. 2017, 426, 111–127. [Google Scholar] [CrossRef]
- Daniele, S.G.; Trummer, G.; Hossmann, K.A.; Vrselja, Z.; Benk, C.; Gobeske, K.T.; Damjanovic, D.; Andrijevic, D.; Pooth, J.S.; Dellal, D.; et al. Brain vulnerability and viability after ischaemia. Nat. Rev. Neurosci. 2021, 22, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Kostandy, B.B. The role of glutamate in neuronal ischemic injury: The role of spark in fire. Neurol. Sci. 2012, 33, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Neves, D.; Salazar, I.L.; Almeida, R.D.; Silva, R.M. Molecular mechanisms of ischemia and glutamate excitotoxicity. Life Sci. 2023, 328, 121814. [Google Scholar] [CrossRef] [PubMed]
- Chalkias, A.; Xanthos, T. Post-cardiac arrest brain injury: Pathophysiology and treatment. J. Neurol. Sci. 2012, 315, 1–8. [Google Scholar] [CrossRef]
- Mayor, D.; Tymianski, M. Neurotransmitters in the mediation of cerebral ischemic injury. Neuropharmacology 2018, 134, 178–188. [Google Scholar] [CrossRef]
- Reis, C.; Akyol, O.; Araujo, C.; Huang, L.; Enkhjargal, B.; Malaguit, J.; Gospodarev, V.; Zhang, J.H. Pathophysiology and the Monitoring Methods for Cardiac Arrest Associated Brain Injury. Int. J. Mol. Sci. 2017, 18, 129. [Google Scholar] [CrossRef]
- Belov Kirdajova, D.; Kriska, J.; Tureckova, J.; Anderova, M. Ischemia-Triggered Glutamate Excitotoxicity from the Perspective of Glial Cells. Front. Cell Neurosci. 2020, 14, 51. [Google Scholar] [CrossRef]
- Bakthavachalam, P.; Shanmugam, P.S.T. Mitochondrial dysfunction—Silent killer in cerebral ischemia. J. Neurol. Sci. 2017, 375, 417–423. [Google Scholar] [CrossRef]
- Pulsinelli, W.A.; Brierley, J.B.; Plum, F. Temporal profile of neuronal damage in a model of transient forebrain ischemia. Ann. Neurol. 1982, 11, 491–498. [Google Scholar] [CrossRef]
- Saito, A.; Maier, C.M.; Narasimhan, P.; Nishi, T.; Song, Y.S.; Yu, F.; Liu, J.; Lee, Y.S.; Nito, C.; Kamada, H.; et al. Oxidative stress and neuronal death/survival signaling in cerebral ischemia. Mol. Neurobiol. 2005, 31, 105–116. [Google Scholar] [CrossRef]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting Oxidative Stress and Inflammation to Prevent Ischemia-Reperfusion Injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.W.; Zou, B.; Fan, Y.; Li, Y.; Lin, N.; Zeng, Y.S.; Gao, T.M.; Yao, Z.; Xu, Z.C. Dendritic plasticity of CA1 pyramidal neurons after transient global ischemia. Neuroscience 2006, 140, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.W.; Han, X.J.; Shi, Z.S.; Lei, Z.G.; Xu, Z.C. Remodeling of synapses in the CA1 area of the hippocampus after transient global ischemia. Neuroscience 2012, 218, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Burgos, I.; Letechipia-Vallejo, G.; Lopez-Loeza, E.; Morali, G.; Cervantes, M. Long-term study of dendritic spines from hippocampal CA1 pyramidal cells, after neuroprotective melatonin treatment following global cerebral ischemia in rats. Neurosci. Lett. 2007, 423, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Block, F. Global ischemia and behavioural deficits. Prog. Neurobiol. 1999, 58, 279–295. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, S.M.; Grubb, N.R.; O’Carroll, R.E. In-hospital cardiac arrest leads to chronic memory impairment. Resuscitation 2003, 58, 73–79. [Google Scholar] [CrossRef]
- Stradecki-Cohan, H.M.; Cohan, C.H.; Raval, A.P.; Dave, K.R.; Reginensi, D.; Gittens, R.A.; Youbi, M.; Perez-Pinzon, M.A. Cognitive Deficits after Cerebral Ischemia and Underlying Dysfunctional Plasticity: Potential Targets for Recovery of Cognition. J. Alzheimers Dis. 2017, 60, S87–S105. [Google Scholar] [CrossRef]
- Orbo, M.C.; Vangberg, T.R.; Tande, P.M.; Anke, A.; Aslaksen, P.M. Memory performance, global cerebral volumes and hippocampal subfield volumes in long-term survivors of Out-of-Hospital Cardiac Arrest. Resuscitation 2018, 126, 21–28. [Google Scholar] [CrossRef]
- Alia, C.; Spalletti, C.; Lai, S.; Panarese, A.; Lamola, G.; Bertolucci, F.; Vallone, F.; Di Garbo, A.; Chisari, C.; Micera, S.; et al. Neuroplastic Changes Following Brain Ischemia and their Contribution to Stroke Recovery: Novel Approaches in Neurorehabilitation. Front. Cell Neurosci. 2017, 11, 76. [Google Scholar] [CrossRef]
- Grefkes, C.; Fink, G.R. Recovery from stroke: Current concepts and future perspectives. Neurol. Res. Pract. 2020, 2, 17. [Google Scholar] [CrossRef]
- Montes, P.; Vigueras-Villasenor, R.M.; Rojas-Castaneda, J.C.; Monfil, T.; Cervantes, M.; Morali, G. Progesterone treatment in rats after severe global cerebral ischemia promotes hippocampal dentate gyrus neurogenesis and functional recovery. Neurol. Res. 2019, 41, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Weihs, W.; Warenits, A.M.; Ettl, F.; Magnet, I.A.M.; Herkner, H.; Kramer, A.M.; Teubenbacher, U.; Lobmeyr, E.; Schriefl, C.; Clodi, C.; et al. CA1 Hippocampal Pyramidal Cells in Rats, Resuscitated from 8 Minutes of Ventricular Fibrillation Cardiac Arrest, Recover after 20 Weeks of Survival: A Retrospective Pilot Study. Shock 2020, 54, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Stroke Therapy Academic Industry Roundtable (STAIR). Recommendations for standards regarding preclinical neuroprotective and restorative drug development. Stroke 1999, 30, 2752–2758. [Google Scholar] [CrossRef]
- Cheng, Y.D.; Al-Khoury, L.; Zivin, J.A. Neuroprotection for ischemic stroke: Two decades of success and failure. NeuroRx 2004, 1, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Wahlgren, N.G.; Ahmed, N. Neuroprotection in cerebral ischaemia: Facts and fancies—The need for new approaches. Cerebrovasc. Dis. 2004, 17 (Suppl. S1), 153–166. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, M.D. Neuroprotection for ischemic stroke: Past, present and future. Neuropharmacology 2008, 55, 363–389. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.Y.; Liu, L.; Yang, Q.W. Refocusing Neuroprotection in Cerebral Reperfusion Era: New Challenges and Strategies. Front. Neurol. 2018, 9, 249. [Google Scholar] [CrossRef]
- Leker, R.R.; Shohami, E. Cerebral ischemia and trauma-different etiologies yet similar mechanisms: Neuroprotective opportunities. Brain Res. Brain Res. Rev. 2002, 39, 55–73. [Google Scholar] [CrossRef]
- Corbett, D.; Nurse, S. The problem of assessing effective neuroprotection in experimental cerebral ischemia. Prog. Neurobiol. 1998, 54, 531–548. [Google Scholar] [CrossRef]
- Moretti, A.; Ferrari, F.; Villa, R.F. Neuroprotection for ischaemic stroke: Current status and challenges. Pharmacol. Ther. 2015, 146, 23–34. [Google Scholar] [CrossRef]
- Schmidt, A.; Minnerup, J. Promoting recovery from ischemic stroke. Expert. Rev. Neurother. 2016, 16, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Cramer, S.C. Recovery after Stroke. Contin. (Minneap Minn.) 2020, 26, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Szelenberger, R.; Kostka, J.; Saluk-Bijak, J.; Miller, E. Pharmacological Interventions and Rehabilitation Approach for Enhancing Brain Self-repair and Stroke Recovery. Curr. Neuropharmacol. 2020, 18, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Diotel, N.; Charlier, T.D.; Lefebvre d’Hellencourt, C.; Couret, D.; Trudeau, V.L.; Nicolau, J.C.; Meilhac, O.; Kah, O.; Pellegrini, E. Steroid Transport, Local Synthesis, and Signaling within the Brain: Roles in Neurogenesis, Neuroprotection, and Sexual Behaviors. Front. Neurosci. 2018, 12, 84. [Google Scholar] [CrossRef] [PubMed]
- Morali, G.; Montes, P.; Hernandez-Morales, L.; Monfil, T.; Espinosa-Garcia, C.; Cervantes, M. Neuroprotective effects of progesterone and allopregnanolone on long-term cognitive outcome after global cerebral ischemia. Restor. Neurol. Neurosci. 2011, 29, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, L.M.; Cornil, C.A.; Mittelman-Smith, M.A.; Rainville, J.R.; Remage-Healey, L.; Sinchak, K.; Micevych, P.E. Actions of Steroids: New Neurotransmitters. J. Neurosci. 2016, 36, 11449–11458. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, R. Neuroactive steroids: Mechanisms of action and neuropsychopharmacological properties. Psychoneuroendocrinology 2003, 28, 139–168. [Google Scholar] [CrossRef]
- Melcangi, R.C.; Giatti, S.; Calabrese, D.; Pesaresi, M.; Cermenati, G.; Mitro, N.; Viviani, B.; Garcia-Segura, L.M.; Caruso, D. Levels and actions of progesterone and its metabolites in the nervous system during physiological and pathological conditions. Prog. Neurobiol. 2014, 113, 56–69. [Google Scholar] [CrossRef]
- Majewska, M.D.; Harrison, N.L.; Schwartz, R.D.; Barker, J.L.; Paul, S.M. Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor. Science 1986, 232, 1004–1007. [Google Scholar] [CrossRef]
- Smith, S.S. Progesterone enhances inhibitory responses of cerebellar Purkinje cells mediated by the GABAA receptor subtype. Brain Res. Bull. 1989, 23, 317–322. [Google Scholar] [CrossRef]
- Cai, W.; Zhu, Y.; Furuya, K.; Li, Z.; Sokabe, M.; Chen, L. Two different molecular mechanisms underlying progesterone neuroprotection against ischemic brain damage. Neuropharmacology 2008, 55, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Garcia, C.; Vigueras-Villasenor, R.M.; Rojas-Castaneda, J.C.; Aguilar-Hernandez, A.; Monfil, T.; Cervantes, M.; Morali, G. Post-ischemic administration of progesterone reduces caspase-3 activation and DNA fragmentation in the hippocampus following global cerebral ischemia. Neurosci. Lett. 2013, 550, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.L.; Liu, J.; Lee, E.; Ling, G.S.; McCabe, J.T. Progesterone differentially regulates pro- and anti-apoptotic gene expression in cerebral cortex following traumatic brain injury in rats. J. Neurotrauma 2005, 22, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Ozacmak, V.H.; Sayan, H. The effects of 17beta estradiol, 17alpha estradiol and progesterone on oxidative stress biomarkers in ovariectomized female rat brain subjected to global cerebral ischemia. Physiol. Res. 2009, 58, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Gibson, C.L.; Constantin, D.; Prior, M.J.; Bath, P.M.; Murphy, S.P. Progesterone suppresses the inflammatory response and nitric oxide synthase-2 expression following cerebral ischemia. Exp. Neurol. 2005, 193, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Garcia, C.; Sayeed, I.; Yousuf, S.; Atif, F.; Sergeeva, E.G.; Neigh, G.N.; Stein, D.G. Stress primes microglial polarization after global ischemia: Therapeutic potential of progesterone. Brain Behav. Immun. 2017, 66, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Betz, A.L.; Coester, H.C. Effect of steroid therapy on ischaemic brain oedema and blood to brain sodium transport. Acta Neurochir. Suppl. (Wien) 1990, 51, 256–258. [Google Scholar] [CrossRef]
- Ishrat, T.; Sayeed, I.; Atif, F.; Hua, F.; Stein, D.G. Progesterone and allopregnanolone attenuate blood-brain barrier dysfunction following permanent focal ischemia by regulating the expression of matrix metalloproteinases. Exp. Neurol. 2010, 226, 183–190. [Google Scholar] [CrossRef]
- Schumacher, M.; Hussain, R.; Gago, N.; Oudinet, J.P.; Mattern, C.; Ghoumari, A.M. Progesterone synthesis in the nervous system: Implications for myelination and myelin repair. Front. Neurosci. 2012, 6, 10. [Google Scholar] [CrossRef]
- Espinosa-Garcia, C.; Aguilar-Hernandez, A.; Cervantes, M.; Morali, G. Effects of progesterone on neurite growth inhibitors in the hippocampus following global cerebral ischemia. Brain Res. 2014, 1545, 23–34. [Google Scholar] [CrossRef]
- Hansberg-Pastor, V.; Gonzalez-Arenas, A.; Pina-Medina, A.G.; Camacho-Arroyo, I. Sex Hormones Regulate Cytoskeletal Proteins Involved in Brain Plasticity. Front. Psychiatry 2015, 6, 165. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, J.; Liu, C.; Jiang, C.; Zhao, C.; Zhu, Z. Progesterone influences postischemic synaptogenesis in the CA1 region of the hippocampus in rats. Synapse 2011, 65, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Zuo, F.; Wang, Y.; Lu, H.; Yang, Q.; Wang, J. Progesterone Changes VEGF and BDNF Expression and Promotes Neurogenesis after Ischemic Stroke. Mol. Neurobiol. 2016, 54, 571–581. [Google Scholar] [CrossRef]
- Novais, A.; Silva, A.; Ferreira, A.C.; Falcao, A.M.; Sousa, N.; Palha, J.A.; Marques, F.; Sousa, J.C. Adult Hippocampal Neurogenesis Modulation by the Membrane-Associated Progesterone Receptor Family Member Neudesin. Front. Cell Neurosci. 2018, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Morali, G.; Montes, P.; Gonzalez-Burgos, I.; Velazquez-Zamora, D.A.; Cervantes, M. Cytoarchitectural characteristics of hippocampal CA1 pyramidal neurons of rats, four months after global cerebral ischemia and progesterone treatment. Restor. Neurol. Neurosci. 2012, 30, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, R.; Cai, W.; Bai, Y.; Sokabe, M.; Chen, L. Treatment with progesterone after focal cerebral ischemia suppresses proliferation of progenitor cells but enhances survival of newborn neurons in adult male mice. Neuropharmacology 2010, 58, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Chopp, M.; Stein, D.; Feit, H. Progesterone is neuroprotective after transient middle cerebral artery occlusion in male rats. Brain Res. 1996, 735, 101–107. [Google Scholar] [CrossRef]
- Chen, J.; Chopp, M.; Li, Y. Neuroprotective effects of progesterone after transient middle cerebral artery occlusion in rat. J. Neurol. Sci. 1999, 171, 24–30. [Google Scholar] [CrossRef]
- Alkayed, N.J.; Murphy, S.J.; Traystman, R.J.; Hurn, P.D.; Miller, V.M. Neuroprotective effects of female gonadal steroids in reproductively senescent female rats. Stroke 2000, 31, 161–168. [Google Scholar] [CrossRef]
- Kumon, Y.; Kim, S.C.; Tompkins, P.; Stevens, A.; Sakaki, S.; Loftus, C.M. Neuroprotective effect of postischemic administration of progesterone in spontaneously hypertensive rats with focal cerebral ischemia. J. Neurosurg. 2000, 92, 848–852. [Google Scholar] [CrossRef]
- Murphy, S.J.; Littleton-Kearney, M.T.; Hurn, P.D. Progesterone administration during reperfusion, but not preischemia alone, reduces injury in ovariectomized rats. J. Cereb. Blood Flow. Metab. 2002, 22, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Gibson, C.L.; Murphy, S.P. Progesterone enhances functional recovery after middle cerebral artery occlusion in male mice. J. Cereb. Blood Flow. Metab. 2004, 24, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Ogaeri, T.; Samsonov, M.; Sokabe, M. Progesterone improves functional outcomes after transient focal cerebral ischemia in both aged male and female rats. Exp. Gerontol. 2018, 113, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Guennoun, R.; Zhu, X.; Frechou, M.; Gaignard, P.; Slama, A.; Liere, P.; Schumacher, M. Steroids in Stroke with Special Reference to Progesterone. Cell Mol. Neurobiol. 2019, 39, 551–568. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Ogaeri, T.; Samsonov, M.; Sokabe, M. The 5alpha-Reductase Inhibitor Finasteride Exerts Neuroprotection against Ischemic Brain Injury in Aged Male Rats. Transl. Stroke Res. 2019, 10, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, I.; Guo, Q.; Hoffman, S.W.; Stein, D.G. Allopregnanolone, a progesterone metabolite, is more effective than progesterone in reducing cortical infarct volume after transient middle cerebral artery occlusion. Ann. Emerg. Med. 2006, 47, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Traystman, R.J. Animal models of focal and global cerebral ischemia. ILAR J. 2003, 44, 85–95. [Google Scholar] [CrossRef]
- Kaya, A.H.; Erdogan, H.; Tasdemiroglu, E. Searching Evidences of Stroke in Animal Models: A Review of Discrepancies A Review of Discrepancies. Turk. Neurosurg. 2017, 27, 167–173. [Google Scholar] [CrossRef]
- Gonzalez-Vidal, M.D.; Cervera-Gaviria, M.; Ruelas, R.; Escobar, A.; Morali, G.; Cervantes, M. Progesterone: Protective effects on the cat hippocampal neuronal damage due to acute global cerebral ischemia. Arch. Med. Res. 1998, 29, 117–124. [Google Scholar]
- Cervantes, M.; Gonzalez-Vidal, M.D.; Ruelas, R.; Escobar, A.; Morali, G. Neuroprotective effects of progesterone on damage elicited by acute global cerebral ischemia in neurons of the caudate nucleus. Arch. Med. Res. 2002, 33, 6–14. [Google Scholar] [CrossRef]
- Aggarwal, R.; Medhi, B.; Pathak, A.; Dhawan, V.; Chakrabarti, A. Neuroprotective effect of progesterone on acute phase changes induced by partial global cerebral ischaemia in mice. J. Pharm. Pharmacol. 2008, 60, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Morali, G.; Letechipia-Vallejo, G.; Lopez-Loeza, E.; Montes, P.; Hernandez-Morales, L.; Cervantes, M. Post-ischemic administration of progesterone in rats exerts neuroprotective effects on the hippocampus. Neurosci. Lett. 2005, 382, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Lieberwirth, C.; Pan, Y.; Liu, Y.; Zhang, Z.; Wang, Z. Hippocampal adult neurogenesis: Its regulation and potential role in spatial learning and memory. Brain Res. 2016, 1644, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Abbott, L.C.; Nigussie, F. Adult neurogenesis in the mammalian dentate gyrus. Anat. Histol. Embryol. 2020, 49, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Garcia, C.; Atif, F.; Yousuf, S.; Sayeed, I.; Neigh, G.N.; Stein, D.G. Progesterone Attenuates Stress-Induced NLRP3 Inflammasome Activation and Enhances Autophagy following Ischemic Brain Injury. Int. J. Mol. Sci. 2020, 21, 3740. [Google Scholar] [CrossRef]
- Hedayatpour, A.; Shiasi, M.; Famitafreshi, H.; Abolhassani, F.; Ebrahimnia, P.; Mokhtari, T.; Hassanzaeh, G.; Karimian, M.; Nazparvar, B.; Marefati, N.; et al. Co-Administration of Progesterone and Melatonin Attenuates Ischemia-Induced Hippocampal Damage in Rats. J. Mol. Neurosci. 2018, 66, 251–260. [Google Scholar] [CrossRef]
- Wolpaw, J.R.; Thompson, A.K. Enhancing neurorehabilitation by targeting beneficial plasticity. Front. Rehabil. Sci. 2023, 4, 1198679. [Google Scholar] [CrossRef]
- Boyce, L.W.; Goossens, P.H. Rehabilitation after Cardiac Arrest: Integration of Neurologic and Cardiac Rehabilitation. Semin. Neurol. 2017, 37, 94–102. [Google Scholar] [CrossRef]
- Geocadin, R.G.; Agarwal, S.; Goss, A.L.; Callaway, C.W.; Richie, M. Cardiac Arrest and Neurologic Recovery: Insights from the Case of Mr. Damar Hamlin. Ann. Neurol. 2023, 93, 871–876. [Google Scholar] [CrossRef]
- Skolnick, B.E.; Maas, A.I.; Narayan, R.K.; van der Hoop, R.G.; MacAllister, T.; Ward, J.D.; Nelson, N.R.; Stocchetti, N.; Investigators, S.T. A clinical trial of progesterone for severe traumatic brain injury. N. Engl. J. Med. 2014, 371, 2467–2476. [Google Scholar] [CrossRef]
- Wright, D.W.; Yeatts, S.D.; Silbergleit, R.; Palesch, Y.Y.; Hertzberg, V.S.; Frankel, M.; Goldstein, F.C.; Caveney, A.F.; Howlett-Smith, H.; Bengelink, E.M.; et al. Very early administration of progesterone for acute traumatic brain injury. N. Engl. J. Med. 2014, 371, 2457–2466. [Google Scholar] [CrossRef] [PubMed]
- Merck, L.H.; Yeatts, S.D.; Silbergleit, R.; Manley, G.T.; Pauls, Q.; Palesch, Y.; Conwit, R.; Le Roux, P.; Miller, J.; Frankel, M.; et al. The Effect of Goal-Directed Therapy on Patient Morbidity and Mortality After Traumatic Brain Injury: Results from the Progesterone for the Treatment of Traumatic Brain Injury III Clinical Trial. Crit. Care Med. 2019, 47, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, M.; Denier, C.; Oudinet, J.P.; Adams, D.; Guennoun, R. Progesterone neuroprotection: The background of clinical trial failure. J. Steroid Biochem. Mol. Biol. 2016, 160, 53–66. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montes, P.; Ortíz-Islas, E.; Rodríguez-Pérez, C.E.; Ruiz-Sánchez, E.; Silva-Adaya, D.; Pichardo-Rojas, P.; Campos-Peña, V. Neuroprotective–Neurorestorative Effects Induced by Progesterone on Global Cerebral Ischemia: A Narrative Review. Pharmaceutics 2023, 15, 2697. https://doi.org/10.3390/pharmaceutics15122697
Montes P, Ortíz-Islas E, Rodríguez-Pérez CE, Ruiz-Sánchez E, Silva-Adaya D, Pichardo-Rojas P, Campos-Peña V. Neuroprotective–Neurorestorative Effects Induced by Progesterone on Global Cerebral Ischemia: A Narrative Review. Pharmaceutics. 2023; 15(12):2697. https://doi.org/10.3390/pharmaceutics15122697
Chicago/Turabian StyleMontes, Pedro, Emma Ortíz-Islas, Citlali Ekaterina Rodríguez-Pérez, Elizabeth Ruiz-Sánchez, Daniela Silva-Adaya, Pavel Pichardo-Rojas, and Victoria Campos-Peña. 2023. "Neuroprotective–Neurorestorative Effects Induced by Progesterone on Global Cerebral Ischemia: A Narrative Review" Pharmaceutics 15, no. 12: 2697. https://doi.org/10.3390/pharmaceutics15122697
APA StyleMontes, P., Ortíz-Islas, E., Rodríguez-Pérez, C. E., Ruiz-Sánchez, E., Silva-Adaya, D., Pichardo-Rojas, P., & Campos-Peña, V. (2023). Neuroprotective–Neurorestorative Effects Induced by Progesterone on Global Cerebral Ischemia: A Narrative Review. Pharmaceutics, 15(12), 2697. https://doi.org/10.3390/pharmaceutics15122697