
An Overview of the Circadian Clock in the Frame of Chronotherapy: From Bench to Bedside


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
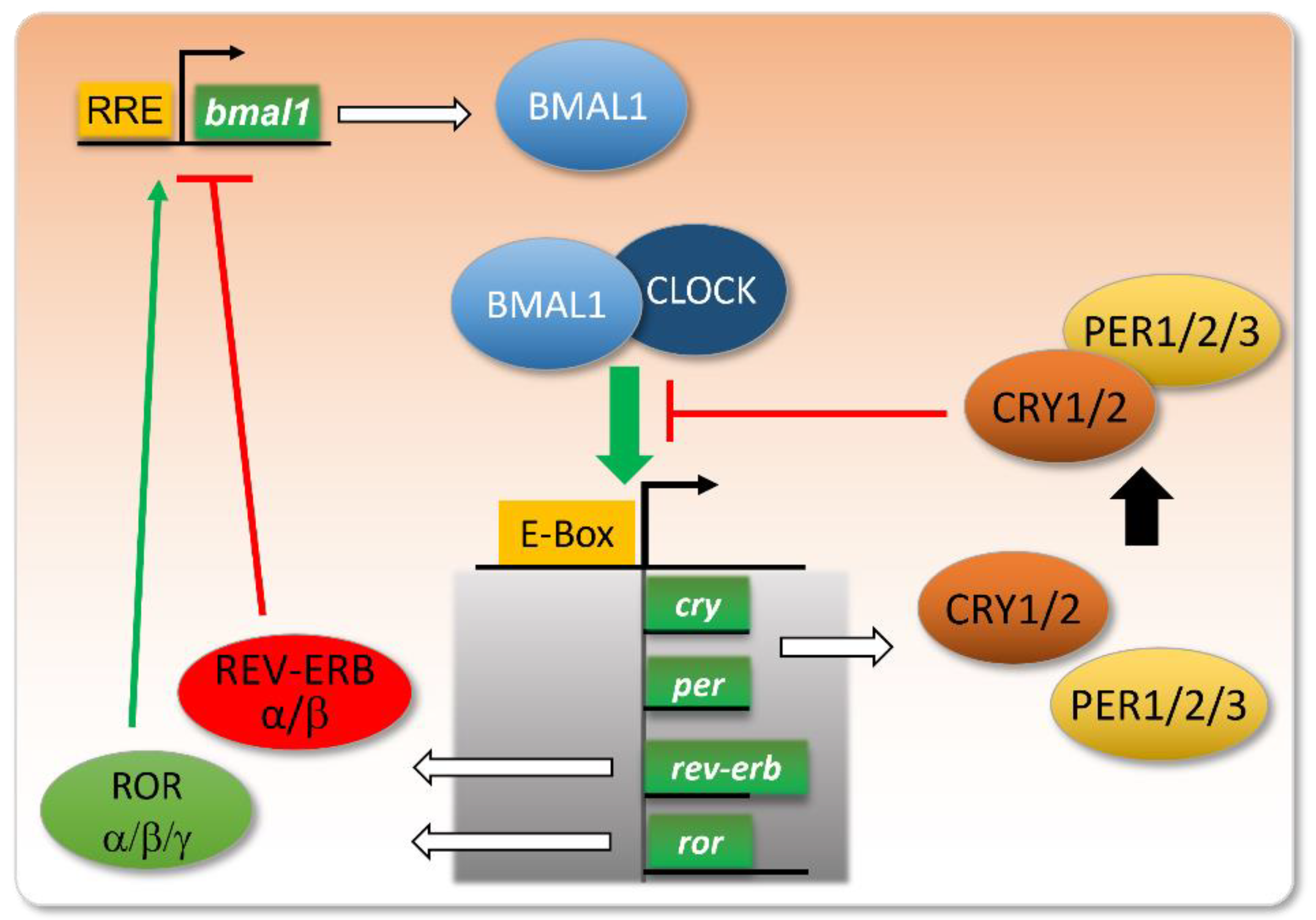
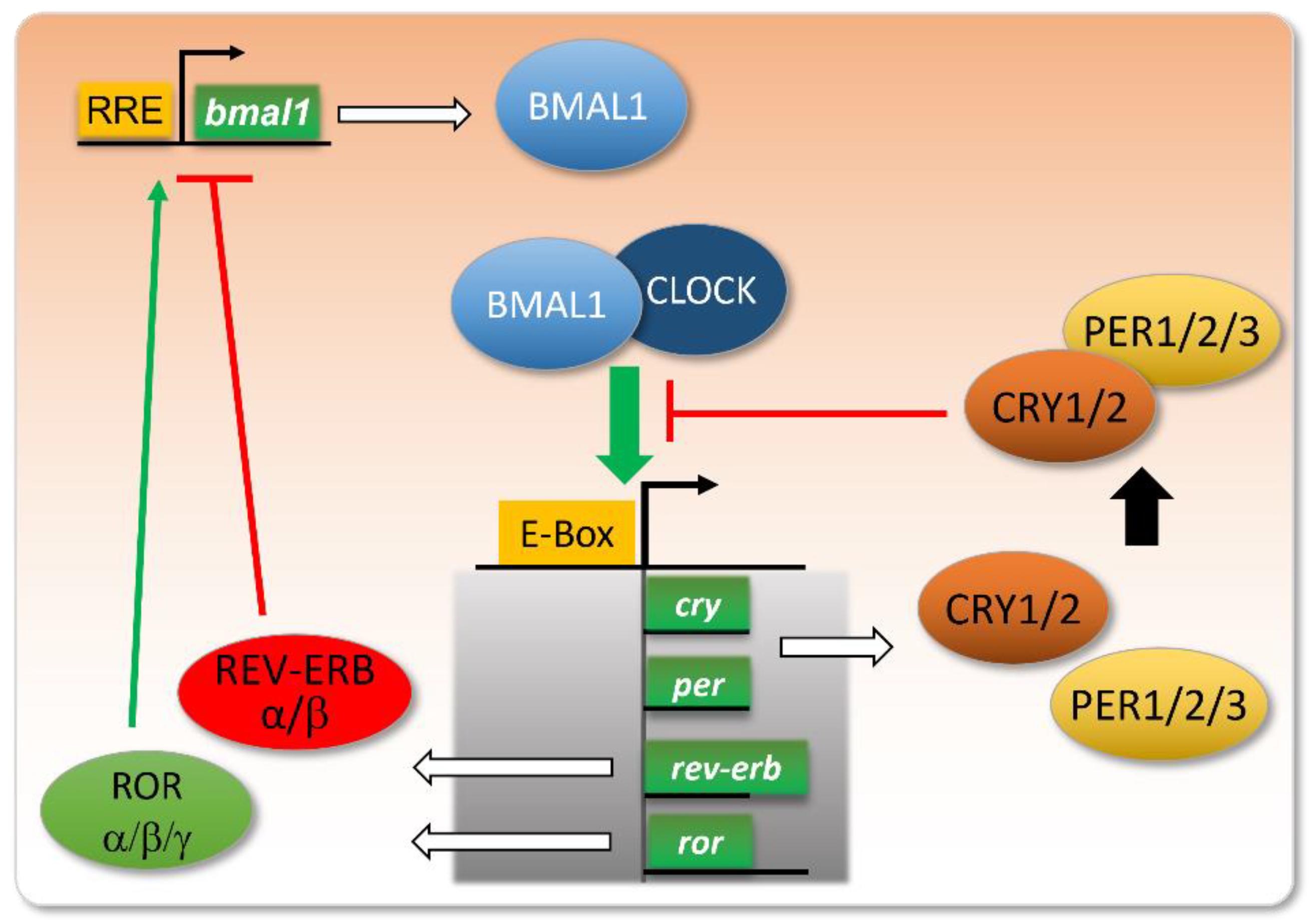
2. A Brief History of Circadian Rhythm Knowledge
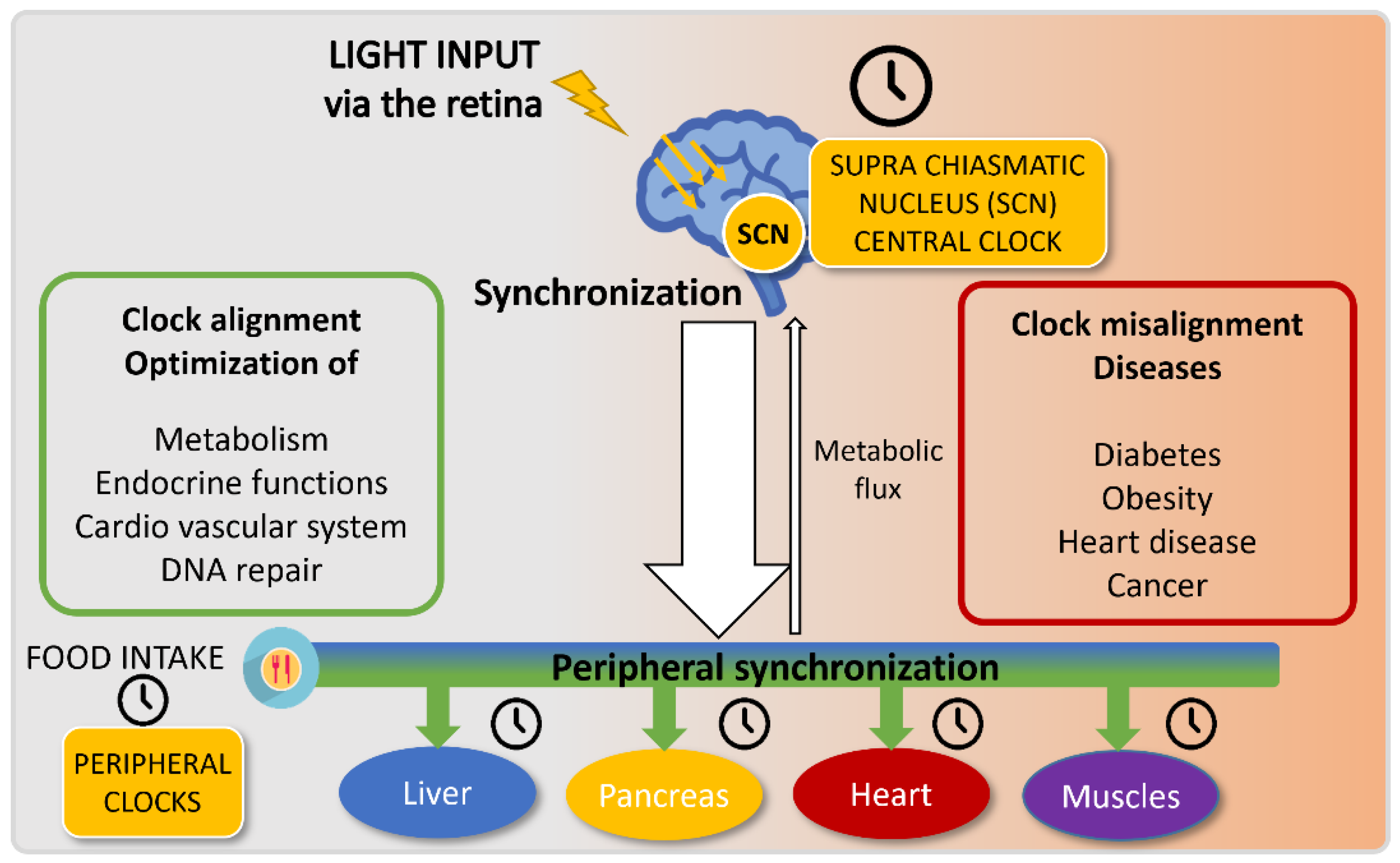
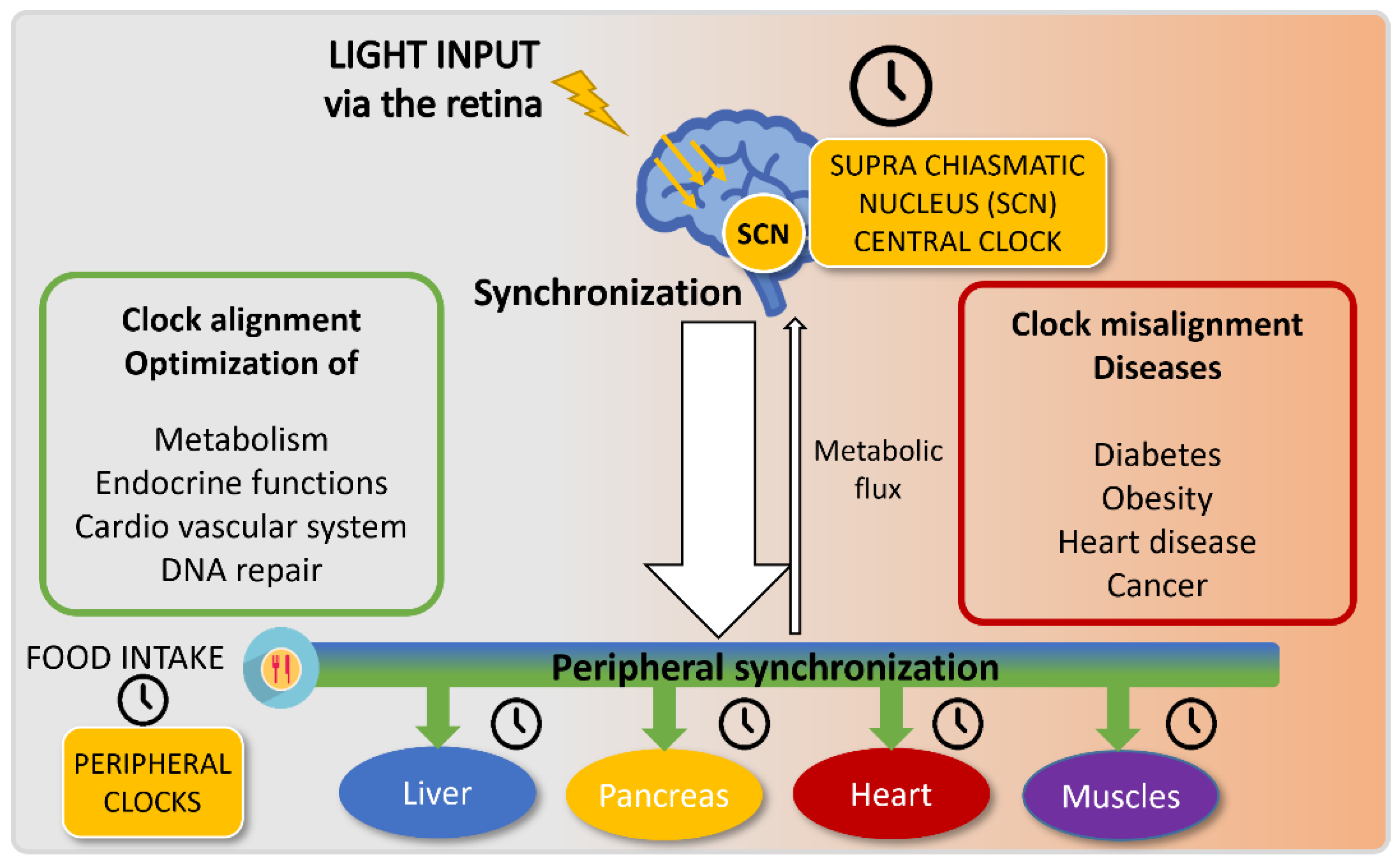
3. Alterations in Day–Night Lifestyle Influence on Human Health: Of the Misdeeds of Human Circadian Misalignment
4. The Molecular Links between the Circadian Clock and Metabolism: The Food Connection
5. A Healthy Way of Life: Get Synchronized with Your Circadian Clock
6. Circadian Medicine and Chronopharmacotherapy
7. Clock, Immunity, and Inflammation
8. Clock and Age-Related Diseases: Molecular Studies Open Therapeutical Perspectives
9. Current Challenges and Perspectives: How to Define the Most Appropriate Timing for Pharmacotherapy
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Musiek, E.S.; Holtzman, D.M. Mechanisms Linking Circadian Clocks, Sleep, and Neurodegeneration. Science 2016, 354, 1004–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagannath, A.; Taylor, L.; Wakaf, Z.; Vasudevan, S.R.; Foster, R.G. The Genetics of Circadian Rhythms, Sleep and Health. Hum. Mol. Genet. 2017, 26, R128–R138. [Google Scholar] [CrossRef] [PubMed]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular Mechanisms and Physiological Importance of Circadian Rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Downton, P.; Early, J.O.; Gibbs, J.E. Circadian Rhythms in Adaptive Immunity. Immunology 2020, 161, 268–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherji, A.; Bailey, S.M.; Staels, B.; Baumert, T.F. The Circadian Clock and Liver Function in Health and Disease. J. Hepatol. 2019, 71, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Pickel, L.; Sung, H.-K. Feeding Rhythms and the Circadian Regulation of Metabolism. Front. Nutr. 2020, 7, 39. [Google Scholar] [CrossRef]
- Bass, J.; Takahashi, J.S. Circadian Integration of Metabolism and Energetics. Science 2010, 330, 1349–1354. [Google Scholar] [CrossRef] [Green Version]
- Zehring, W.A.; Wheeler, D.A.; Reddy, P.; Konopka, R.J.; Kyriacou, C.P.; Rosbash, M.; Hall, J.C. P-Element Transformation with Period Locus DNA Restores Rhythmicity to Mutant, Arrhythmic Drosophila Melanogaster. Cell 1984, 39, 369–376. [Google Scholar] [CrossRef]
- Reddy, P.; Zehring, W.A.; Wheeler, D.A.; Pirrotta, V.; Hadfield, C.; Hall, J.C.; Rosbash, M. Molecular Analysis of the Period Locus in Drosophila Melanogaster and Identification of a Transcript Involved in Biological Rhythms. Cell 1984, 38, 701–710. [Google Scholar] [CrossRef]
- Baylies, M.K.; Bargiello, T.A.; Jackson, F.R.; Young, M.W. Changes in Abundance or Structure of the per Gene Product Can Alter Periodicity of the Drosophila Clock. Nature 1987, 326, 390–392. [Google Scholar] [CrossRef]
- De Mairan, J.-J. Observation Botanique. Hist. Acad. R. Sci. 1729, 35–36. Available online: http://www.bibnum.education.fr/sciencesdelavie/biologie/observation-botanique (accessed on 4 May 2022).
- Konopka, R.J.; Benzer, S. Clock Mutants of Drosophila Melanogaster. Proc. Natl. Acad. Sci. USA 1971, 68, 2112–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, J.S. Transcriptional Architecture of the Mammalian Circadian Clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A Circadian Gene Expression Atlas in Mammals: Implications for Biology and Medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J.S.; Reddy, A.B. Circadian Clocks in Human Red Blood Cells. Nature 2011, 469, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Rey, G.; Valekunja, U.K.; Feeney, K.A.; Wulund, L.; Milev, N.B.; Stangherlin, A.; Ansel-Bollepalli, L.; Velagapudi, V.; O’Neill, J.S.; Reddy, A.B. The Pentose Phosphate Pathway Regulates the Circadian Clock. Cell Metab. 2016, 24, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Dibner, C.; Schibler, U.; Albrecht, U. The Mammalian Circadian Timing System: Organization and Coordination of Central and Peripheral Clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [Green Version]
- Kent, B.A.; Rahman, S.A.; St Hilaire, M.A.; Grant, L.K.; Rüger, M.; Czeisler, C.A.; Lockley, S.W. Circadian Lipid and Hepatic Protein Rhythms Shift with a Phase Response Curve Different than Melatonin. Nat. Commun. 2022, 13, 681. [Google Scholar] [CrossRef]
- Hong, F.; Pan, S.; Xu, P.; Xue, T.; Wang, J.; Guo, Y.; Jia, L.; Qiao, X.; Li, L.; Zhai, Y. Melatonin Orchestrates Lipid Homeostasis through the Hepatointestinal Circadian Clock and Microbiota during Constant Light Exposure. Cells 2020, 9, 489. [Google Scholar] [CrossRef] [Green Version]
- Damiola, F.; Le Minh, N.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted Feeding Uncouples Circadian Oscillators in Peripheral Tissues from the Central Pacemaker in the Suprachiasmatic Nucleus. Genes Dev. 2000, 14, 2950–2961. [Google Scholar] [CrossRef] [Green Version]
- Sen, S.; Raingard, H.; Dumont, S.; Kalsbeek, A.; Vuillez, P.; Challet, E. Ultradian Feeding in Mice Not Only Affects the Peripheral Clock in the Liver, but Also the Master Clock in the Brain. Chronobiol. Int. 2017, 34, 17–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, T.M.; Markwald, R.R.; McHill, A.W.; Chinoy, E.D.; Snider, J.A.; Bessman, S.C.; Jung, C.M.; O’Neill, J.S.; Wright, K.P. Effects of Caffeine on the Human Circadian Clock in Vivo and in Vitro. Sci. Transl. Med. 2015, 7, 305ra146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, C.; Roehrs, T.; Shambroom, J.; Roth, T. Caffeine Effects on Sleep Taken 0, 3, or 6 Hours before Going to Bed. J. Clin. Sleep Med. 2013, 9, 1195–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, S.M.; Honn, K.A.; Gaddameedhi, S.; Van Dongen, H.P.A. Shift Work: Disrupted Circadian Rhythms and Sleep-Implications for Health and Well-Being. Curr. Sleep Med. Rep. 2017, 3, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Inokawa, H.; Umemura, Y.; Shimba, A.; Kawakami, E.; Koike, N.; Tsuchiya, Y.; Ohashi, M.; Minami, Y.; Cui, G.; Asahi, T.; et al. Chronic Circadian Misalignment Accelerates Immune Senescence and Abbreviates Lifespan in Mice. Sci. Rep. 2020, 10, 2569. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kuehnle, T.; Pramstaller, P.P.; Ricken, J.; Havel, M.; Guth, A.; Merrow, M. A Marker for the End of Adolescence. Curr. Biol. 2004, 14, R1038–R1039. [Google Scholar] [CrossRef] [Green Version]
- Hida, A.; Kitamura, S.; Ohsawa, Y.; Enomoto, M.; Katayose, Y.; Motomura, Y.; Moriguchi, Y.; Nozaki, K.; Watanabe, M.; Aritake, S.; et al. In Vitro Circadian Period Is Associated with Circadian/Sleep Preference. Sci. Rep. 2013, 3, 2074. [Google Scholar] [CrossRef] [Green Version]
- Taillard, J.; Sagaspe, P.; Philip, P.; Bioulac, S. Sleep Timing, Chronotype and Social Jetlag: Impact on Cognitive Abilities and Psychiatric Disorders. Biochem. Pharm. 2021, 191, 114438. [Google Scholar] [CrossRef]
- Goldin, A.P.; Sigman, M.; Braier, G.; Golombek, D.A.; Leone, M.J. Interplay of Chronotype and School Timing Predicts School Performance. Nat. Hum. Behav. 2020, 4, 387–396. [Google Scholar] [CrossRef]
- Roenneberg, T.; Pilz, L.K.; Zerbini, G.; Winnebeck, E.C. Chronotype and Social Jetlag: A (Self-) Critical Review. Biology 2019, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Islam, Z.; Hu, H.; Akter, S.; Kuwahara, K.; Kochi, T.; Eguchi, M.; Kurotani, K.; Nanri, A.; Kabe, I.; Mizoue, T. Social Jetlag Is Associated with an Increased Likelihood of Having Depressive Symptoms among the Japanese Working Population: The Furukawa Nutrition and Health Study. Sleep 2020, 43, zsz204. [Google Scholar] [CrossRef] [PubMed]
- Makarem, N.; Paul, J.; Giardina, E.-G.V.; Liao, M.; Aggarwal, B. Evening Chronotype Is Associated with Poor Cardiovascular Health and Adverse Health Behaviors in a Diverse Population of Women. Chronobiol. Int. 2020, 37, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Knutson, K.L.; von Schantz, M. Associations between Chronotype, Morbidity and Mortality in the UK Biobank Cohort. Chronobiol. Int. 2018, 35, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Hood, S.; Amir, S. The Aging Clock: Circadian Rhythms and Later Life. J. Clin. Investig. 2017, 127, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Zitting, K.-M.; Chinoy, E.D. Aging and Circadian Rhythms. Sleep Med. Clin. 2015, 10, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and Metabolic Syndrome in Circadian Clock Mutant Mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef] [Green Version]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the Clock Components CLOCK and BMAL1 Leads to Hypoinsulinaemia and Diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Duez, H.; Staels, B. Nuclear Receptors Linking Circadian Rhythms and Cardiometabolic Control. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1529–1534. [Google Scholar] [CrossRef]
- Solt, L.A.; Wang, Y.; Banerjee, S.; Hughes, T.; Kojetin, D.J.; Lundasen, T.; Shin, Y.; Liu, J.; Cameron, M.D.; Noel, R.; et al. Regulation of Circadian Behaviour and Metabolism by Synthetic REV-ERB Agonists. Nature 2012, 485, 62–68. [Google Scholar] [CrossRef]
- Berthier, A.; Vinod, M.; Porez, G.; Steenackers, A.; Alexandre, J.; Yamakawa, N.; Gheeraert, C.; Ploton, M.; Maréchal, X.; Dubois-Chevalier, J.; et al. Combinatorial Regulation of Hepatic Cytoplasmic Signaling and Nuclear Transcriptional Events by the OGT/REV-ERBα Complex. Proc. Natl. Acad. Sci. USA 2018, 115, E11033–E11042. [Google Scholar] [CrossRef] [Green Version]
- Maayan, D.-F.; Chapnik, N.; Froy, O. REV-ERBα Activates the MTOR Signaling Pathway and Promotes Myotubes Differentiation. Biol. Cell 2020, 112, 213–221. [Google Scholar] [CrossRef]
- Ramanathan, C.; Kathale, N.D.; Liu, D.; Lee, C.; Freeman, D.A.; Hogenesch, J.B.; Cao, R.; Liu, A.C. MTOR Signaling Regulates Central and Peripheral Circadian Clock Function. PLoS Genet. 2018, 14, e1007369. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, Y.; Sahar, S.; Astarita, G.; Kaluzova, M.; Sassone-Corsi, P. Circadian Control of the NAD+ Salvage Pathway by CLOCK-SIRT1. Science 2009, 324, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, K.M.; Yoshino, J.; Brace, C.S.; Abrassart, D.; Kobayashi, Y.; Marcheva, B.; Hong, H.-K.; Chong, J.L.; Buhr, E.D.; Lee, C.; et al. Circadian Clock Feedback Cycle through NAMPT-Mediated NAD+ Biosynthesis. Science 2009, 324, 651–654. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Li, S.; Liu, T.; Borjigin, J.; Lin, J.D. Transcriptional Coactivator PGC-1alpha Integrates the Mammalian Clock and Energy Metabolism. Nature 2007, 447, 477–481. [Google Scholar] [CrossRef]
- Cantó, C.; Jiang, L.Q.; Deshmukh, A.S.; Mataki, C.; Coste, A.; Lagouge, M.; Zierath, J.R.; Auwerx, J. Interdependence of AMPK and SIRT1 for Metabolic Adaptation to Fasting and Exercise in Skeletal Muscle. Cell Metab. 2010, 11, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Dyar, K.A.; Lutter, D.; Artati, A.; Ceglia, N.J.; Liu, Y.; Armenta, D.; Jastroch, M.; Schneider, S.; de Mateo, S.; Cervantes, M.; et al. Atlas of Circadian Metabolism Reveals System-Wide Coordination and Communication between Clocks. Cell 2018, 174, 1571–1585.e11. [Google Scholar] [CrossRef] [Green Version]
- Wehrens, S.M.T.; Christou, S.; Isherwood, C.; Middleton, B.; Gibbs, M.A.; Archer, S.N.; Skene, D.J.; Johnston, J.D. Meal Timing Regulates the Human Circadian System. Curr. Biol. 2017, 27, 1768–1775.e3. [Google Scholar] [CrossRef] [Green Version]
- Mukherji, A.; Kobiita, A.; Damara, M.; Misra, N.; Meziane, H.; Champy, M.-F.; Chambon, P. Shifting Eating to the Circadian Rest Phase Misaligns the Peripheral Clocks with the Master SCN Clock and Leads to a Metabolic Syndrome. Proc. Natl. Acad. Sci. USA 2015, 112, E6691–E6698. [Google Scholar] [CrossRef] [Green Version]
- Eckel-Mahan, K.L.; Patel, V.R.; de Mateo, S.; Orozco-Solis, R.; Ceglia, N.J.; Sahar, S.; Dilag-Penilla, S.A.; Dyar, K.A.; Baldi, P.; Sassone-Corsi, P. Reprogramming of the Circadian Clock by Nutritional Challenge. Cell 2013, 155, 1464–1478. [Google Scholar] [CrossRef] [Green Version]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tal, Y.; Chapnik, N.; Froy, O. Non-Obesogenic Doses of Fatty Acids Modulate the Functionality of the Circadian Clock in the Liver. Cell. Mol. Life Sci. 2019, 76, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.J.; et al. Time-Restricted Feeding without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaix, A.; Lin, T.; Le, H.D.; Chang, M.W.; Panda, S. Time-Restricted Feeding Prevents Obesity and Metabolic Syndrome in Mice Lacking a Circadian Clock. Cell Metab. 2019, 29, 303–319.e4. [Google Scholar] [CrossRef]
- Lewis, P.; Oster, H.; Korf, H.W.; Foster, R.G.; Erren, T.C. Food as a Circadian Time Cue-Evidence from Human Studies. Nat. Rev. Endocrinol. 2020, 16, 213–223. [Google Scholar] [CrossRef]
- Di Francesco, A.; Di Germanio, C.; Bernier, M.; de Cabo, R. A Time to Fast. Science 2018, 362, 770–775. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Panda, S. Fasting, Circadian Rhythms, and Time-Restricted Feeding in Healthy Lifespan. Cell Metab. 2016, 23, 1048–1059. [Google Scholar] [CrossRef] [Green Version]
- Ibarz-Blanch, N.; Morales, D.; Calvo, E.; Ros-Medina, L.; Muguerza, B.; Bravo, F.I.; Suárez, M. Role of Chrononutrition in the Antihypertensive Effects of Natural Bioactive Compounds. Nutrients 2022, 14, 1920. [Google Scholar] [CrossRef]
- Mi, Y.; Qi, G.; Fan, R.; Ji, X.; Liu, Z.; Liu, X. EGCG Ameliorates Diet-Induced Metabolic Syndrome Associating with the Circadian Clock. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 1575–1589. [Google Scholar] [CrossRef]
- Shan, Z.; Li, Y.; Zong, G.; Guo, Y.; Li, J.; Manson, J.E.; Hu, F.B.; Willett, W.C.; Schernhammer, E.S.; Bhupathiraju, S.N. Rotating Night Shift Work and Adherence to Unhealthy Lifestyle in Predicting Risk of Type 2 Diabetes: Results from Two Large US Cohorts of Female Nurses. BMJ 2018, 363, k4641. [Google Scholar] [CrossRef] [Green Version]
- Grant, C.L.; Coates, A.M.; Dorrian, J.; Kennaway, D.J.; Wittert, G.A.; Heilbronn, L.K.; Pajcin, M.; Della Vedova, C.; Gupta, C.C.; Banks, S. Timing of Food Intake during Simulated Night Shift Impacts Glucose Metabolism: A Controlled Study. Chronobiol. Int. 2017, 34, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Vetter, C.; Fischer, D.; Matera, J.L.; Roenneberg, T. Aligning Work and Circadian Time in Shift Workers Improves Sleep and Reduces Circadian Disruption. Curr. Biol. 2015, 25, 907–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, J.A.; Artenie, D.Z.; Cyr, M.; Raz, A.; Lee, V. Developing a Light-Based Intervention to Reduce Fatigue and Improve Sleep in Rapidly Rotating Shift Workers. Chronobiol. Int. 2020, 37, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between Light at Night, Melatonin Secretion, Sleep Deprivation, and the Internal Clock: Health Impacts and Mechanisms of Circadian Disruption. Life Sci. 2017, 173, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Gunata, M.; Parlakpinar, H.; Acet, H.A. Melatonin: A Review of Its Potential Functions and Effects on Neurological Diseases. Rev. Neurol. 2020, 176, 148–165. [Google Scholar] [CrossRef]
- Houser, K.W.; Esposito, T. Human-Centric Lighting: Foundational Considerations and a Five-Step Design Process. Front. Neurol. 2021, 12, 630553. [Google Scholar] [CrossRef]
- Stefani, O.; Cajochen, C. Should We Re-Think Regulations and Standards for Lighting at Workplaces? A Practice Review on Existing Lighting Recommendations. Front. Psychiatry 2021, 12, 652161. [Google Scholar] [CrossRef]
- Morse, A.M.; Bender, E. Sleep in Hospitalized Patients. Clocks Sleep 2019, 1, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Bano, M.; Chiaromanni, F.; Corrias, M.; Turco, M.; De Rui, M.; Amodio, P.; Merkel, C.; Gatta, A.; Mazzotta, G.; Costa, R.; et al. The Influence of Environmental Factors on Sleep Quality in Hospitalized Medical Patients. Front. Neurol. 2014, 5, 267. [Google Scholar] [CrossRef] [Green Version]
- Palada, V.; Gilron, I.; Canlon, B.; Svensson, C.I.; Kalso, E. The Circadian Clock at the Intercept of Sleep and Pain. Pain 2019, 161, 894–900. [Google Scholar] [CrossRef]
- Manella, G.; Aviram, R.; Bolshette, N.; Muvkadi, S.; Golik, M.; Smith, D.F.; Asher, G. Hypoxia Induces a Time- and Tissue-Specific Response That Elicits Intertissue Circadian Clock Misalignment. Proc. Natl. Acad. Sci. USA 2020, 117, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Ohdo, S. Chrono-Drug Discovery and Development Based on Circadian Rhythm of Molecular, Cellular and Organ Level. Biol. Pharm. Bull. 2021, 44, 747–761. [Google Scholar] [CrossRef] [PubMed]
- Turek, F.W. Circadian Clocks: Not Your Grandfather’s Clock. Science 2016, 354, 992–993. [Google Scholar] [CrossRef] [PubMed]
- Crnko, S.; Du Pré, B.C.; Sluijter, J.P.G.; Van Laake, L.W. Circadian Rhythms and the Molecular Clock in Cardiovascular Biology and Disease. Nat. Rev. Cardiol. 2019, 16, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.E.; Stone, P.H.; Turi, Z.G.; Rutherford, J.D.; Czeisler, C.A.; Parker, C.; Poole, W.K.; Passamani, E.; Roberts, R.; Robertson, T. Circadian Variation in the Frequency of Onset of Acute Myocardial Infarction. N. Engl. J. Med. 1985, 313, 1315–1322. [Google Scholar] [CrossRef]
- Shi, J.; Tong, R.; Zhou, M.; Gao, Y.; Zhao, Y.; Chen, Y.; Liu, W.; Li, G.; Lu, D.; Meng, G.; et al. Circadian Nuclear Receptor Rev-Erbα Is Expressed by Platelets and Potentiates Platelet Activation and Thrombus Formation. Eur. Heart J. 2022, 43, 2317–2334. [Google Scholar] [CrossRef]
- Krasińska, B.; Paluszkiewicz, L.; Miciak-Lawicka, E.; Krasiński, M.; Rzymski, P.; Tykarski, A.; Krasiński, Z. The Effect of Acetylsalicylic Acid Dosed at Bedtime on the Anti-Aggregation Effect in Patients with Coronary Heart Disease and Arterial Hypertension: A Randomized, Controlled Trial. Cardiol. J. 2019, 26, 727–735. [Google Scholar] [CrossRef] [Green Version]
- Sobiczewski, W.; Wirtwein, M.; Gruchała, M.; Kocić, I. Mortality in Hypertensive Patients with Coronary Heart Disease Depends on Chronopharmacotherapy and Dipping Status. Pharmacol. Rep. 2014, 66, 448–452. [Google Scholar] [CrossRef]
- Smolensky, M.H.; Hermida, R.C.; Geng, Y.-J. Chronotherapy of Cardiac and Vascular Disease: Timing Medications to Circadian Rhythms to Optimize Treatment Effects and Outcomes. Curr. Opin. Pharmacol. 2021, 57, 41–48. [Google Scholar] [CrossRef]
- Durgan, D.J.; Pulinilkunnil, T.; Villegas-Montoya, C.; Garvey, M.E.; Frangogiannis, N.G.; Michael, L.H.; Chow, C.-W.; Dyck, J.R.B.; Young, M.E. Short Communication: Ischemia/Reperfusion Tolerance Is Time-of-Day-Dependent: Mediation by the Cardiomyocyte Circadian Clock. Circ. Res. 2010, 106, 546–550. [Google Scholar] [CrossRef]
- Montaigne, D.; Marechal, X.; Modine, T.; Coisne, A.; Mouton, S.; Fayad, G.; Ninni, S.; Klein, C.; Ortmans, S.; Seunes, C.; et al. Daytime Variation of Perioperative Myocardial Injury in Cardiac Surgery and Its Prevention by Rev-Erbα Antagonism: A Single-Centre Propensity-Matched Cohort Study and a Randomised Study. Lancet 2018, 391, 59–69. [Google Scholar] [CrossRef]
- Montaigne, D.; Staels, B. Time to Check the Clock in Cardiovascular Research and Medicine. Circ. Res. 2018, 123, 648–650. [Google Scholar] [CrossRef]
- Pariollaud, M.; Gibbs, J.E.; Hopwood, T.W.; Brown, S.; Begley, N.; Vonslow, R.; Poolman, T.; Guo, B.; Saer, B.; Jones, D.H.; et al. Circadian Clock Component REV-ERBα Controls Homeostatic Regulation of Pulmonary Inflammation. J. Clin. Investig. 2018, 128, 2281–2296. [Google Scholar] [CrossRef] [Green Version]
- Leone, M.J.; Marpegan, L.; Duhart, J.M.; Golombek, D.A. Role of Proinflammatory Cytokines on Lipopolysaccharide-Induced Phase Shifts in Locomotor Activity Circadian Rhythm. Chronobiol. Int. 2012, 29, 715–723. [Google Scholar] [CrossRef]
- Kim, S.-M.; Neuendorff, N.; Chapkin, R.S.; Earnest, D.J. Role of Inflammatory Signaling in the Differential Effects of Saturated and Poly-Unsaturated Fatty Acids on Peripheral Circadian Clocks. E. Bio. Med. 2016, 7, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.S.; Stangherlin, A.; Nagy, A.D.; Nicoll, M.P.; Efstathiou, S.; O’Neill, J.S.; Reddy, A.B. Cell Autonomous Regulation of Herpes and Influenza Virus Infection by the Circadian Clock. Proc. Natl. Acad. Sci. USA 2016, 113, 10085–10090. [Google Scholar] [CrossRef] [Green Version]
- Heer, C.D.; Sanderson, D.J.; Voth, L.S.; Alhammad, Y.M.O.; Schmidt, M.S.; Trammell, S.A.J.; Perlman, S.; Cohen, M.S.; Fehr, A.R.; Brenner, C. Coronavirus Infection and PARP Expression Dysregulate the NAD Metabolome: An Actionable Component of Innate Immunity. J. Biol. Chem. 2020, 295, 17986–17996. [Google Scholar] [CrossRef]
- Zheng, M.; Schultz, M.B.; Sinclair, D.A. NAD+ in COVID-19 and Viral Infections. Trends. Immunol. 2022, 43, 283–295. [Google Scholar] [CrossRef]
- Burioka, N.; Fukuoka, Y.; Koyanagi, S.; Miyata, M.; Takata, M.; Chikumi, H.; Takane, H.; Watanabe, M.; Endo, M.; Sako, T.; et al. Asthma: Chronopharmacotherapy and the Molecular Clock. Adv. Drug. Deliv. Rev. 2010, 62, 946–955. [Google Scholar] [CrossRef]
- Giri, A.; Wang, Q.; Rahman, I.; Sundar, I.K. Circadian Molecular Clock Disruption in Chronic Pulmonary Diseases. Trends Mol. Med. 2022, 28, 513–527. [Google Scholar] [CrossRef]
- Wang, S.; Lin, Y.; Yuan, X.; Li, F.; Guo, L.; Wu, B. REV-ERBα Integrates Colon Clock with Experimental Colitis through Regulation of NF-ΚB/NLRP3 Axis. Nat. Commun. 2018, 9, 4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.; Peigneux, P.; Cajochen, C.; Collette, F. Adapting Test Timing to the Sleep-Wake Schedule: Effects on Diurnal Neurobehavioral Performance Changes in Young Evening and Older Morning Chronotypes. Chronobiol. Int. 2012, 29, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Zeitzer, J.M.; Daniels, J.E.; Duffy, J.F.; Klerman, E.B.; Shanahan, T.L.; Dijk, D.J.; Czeisler, C.A. Do Plasma Melatonin Concentrations Decline with Age? Am. J. Med. 1999, 107, 432–436. [Google Scholar] [CrossRef]
- Hoyt, K.R.; Obrietan, K. Circadian Clocks, Cognition, and Alzheimer’s Disease: Synaptic Mechanisms, Signaling Effectors, and Chronotherapeutics. Mol. Neurodegener. 2022, 17, 35. [Google Scholar] [CrossRef]
- Wirianto, M.; Wang, C.-Y.; Kim, E.; Koike, N.; Gomez-Gutierrez, R.; Nohara, K.; Escobedo, G.; Choi, J.M.; Han, C.; Yagita, K.; et al. The Clock Modulator Nobiletin Mitigates Astrogliosis-Associated Neuroinflammation and Disease Hallmarks in an Alzheimer’s Disease Model. FASEB J. 2022, 36, e22186. [Google Scholar] [CrossRef]
- Harper, D.G.; Volicer, L.; Stopa, E.G.; McKee, A.C.; Nitta, M.; Satlin, A. Disturbance of Endogenous Circadian Rhythm in Aging and Alzheimer Disease. Am. J. Geriatr. Psychiatry 2005, 13, 359–368. [Google Scholar] [CrossRef]
- Gubin, D.G.; Malishevskaya, T.N.; Astakhov, Y.S.; Astakhov, S.Y.; Cornelissen, G.; Kuznetsov, V.A.; Weinert, D. Progressive Retinal Ganglion Cell Loss in Primary Open-Angle Glaucoma Is Associated with Temperature Circadian Rhythm Phase Delay and Compromised Sleep. Chronobiol. Int. 2019, 36, 564–577. [Google Scholar] [CrossRef]
- Gubin, D.; Neroev, V.; Malishevskaya, T.; Cornelissen, G.; Astakhov, S.Y.; Kolomeichuk, S.; Yuzhakova, N.; Kabitskaya, Y.; Weinert, D.C. Melatonin mitigates disrupted circadian rhythms, lowers intraocular pressure, and improves retinal ganglion cells function in glaucoma. J. Pineal Res. 2021, 70, e12730. [Google Scholar] [CrossRef]
- Kim, J.; Park, I.; Jang, S.; Choi, M.; Kim, D.; Sun, W.; Choe, Y.; Choi, J.-W.; Moon, C.; Park, S.H.; et al. Pharmacological Rescue with SR8278, a Circadian Nuclear Receptor REV-ERBα Antagonist as a Therapy for Mood Disorders in Parkinson’s Disease. Neurotherapeutics 2022, 19, 592–607. [Google Scholar] [CrossRef]
- Froy, O. Circadian Rhythms, Nutrition and Implications for Longevity in Urban Environments. Proc. Nutr. Soc. 2018, 77, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Rodríguez, V.A.; Rijo-Ferreira, F.; Green, C.B.; Takahashi, J.S. Importance of Circadian Timing for Aging and Longevity. Nat. Commun. 2021, 12, 2862. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-C.; Guarente, L. SIRT1 Mediates Central Circadian Control in the SCN by a Mechanism That Decays with Aging. Cell 2013, 153, 1448–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, E.S.; Song, S.B. Possible Adverse Effects of High-Dose Nicotinamide: Mechanisms and Safety Assessment. Biomolecules 2020, 10, 687. [Google Scholar] [CrossRef]
- Guenthner, C.J.; Luitje, M.E.; Pyle, L.A.; Molyneux, P.C.; Yu, J.K.; Li, A.S.; Leise, T.L.; Harrington, M.E. Circadian Rhythms of Per2::Luc in Individual Primary Mouse Hepatocytes and Cultures. PLoS ONE 2014, 9, e87573. [Google Scholar] [CrossRef] [Green Version]
- Woller, A.; Duez, H.; Staels, B.; Lefranc, M. A Mathematical Model of the Liver Circadian Clock Linking Feeding and Fasting Cycles to Clock Function. Cell Rep. 2016, 17, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Lamia, K.A.; Sachdeva, U.M.; DiTacchio, L.; Williams, E.C.; Alvarez, J.G.; Egan, D.F.; Vasquez, D.S.; Juguilon, H.; Panda, S.; Shaw, R.J.; et al. AMPK Regulates the Circadian Clock by Cryptochrome Phosphorylation and Degradation. Science 2009, 326, 437–440. [Google Scholar] [CrossRef] [Green Version]
- Furlan, A.; Jacquier, M.; Woller, A.; Héliot, L.; Duez, H.; Staels, B.; Lefranc, M. Mathematical Models Converge on PGC1α as the Key Metabolic Integrator of SIRT1 and AMPK Regulation of the Circadian Clock. Proc. Natl. Acad. Sci. USA 2019, 116, 13171–13172. [Google Scholar] [CrossRef] [Green Version]
- Foteinou, P.T.; Venkataraman, A.; Francey, L.J.; Anafi, R.C.; Hogenesch, J.B.; Doyle, F.J. Computational and Experimental Insights into the Circadian Effects of SIRT1. Proc. Natl. Acad. Sci. USA 2018, 115, 11643–11648. [Google Scholar] [CrossRef] [Green Version]
- Sadria, M.; Layton, A.T. Aging Affects Circadian Clock and Metabolism and Modulates Timing of Medication. iScience 2021, 24, 102245. [Google Scholar] [CrossRef]
- Gadacha, W.; Ben-Attia, M.; Bonnefont-Rousselot, D.; Aouani, E.; Ghanem-Boughanmi, N.; Touitou, Y. Resveratrol Opposite Effects on Rat Tissue Lipoperoxidation: Pro-Oxidant during Day-Time and Antioxidant at Night. Redox. Rep. 2009, 14, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Lévi, F.; Zidani, R.; Misset, J.L. Randomised Multicentre Trial of Chronotherapy with Oxaliplatin, Fluorouracil, and Folinic Acid in Metastatic Colorectal Cancer. International Organization for Cancer Chronotherapy. Lancet 1997, 350, 681–686. [Google Scholar] [CrossRef]
- Okyar, A.; Kumar, S.A.; Filipski, E.; Piccolo, E.; Ozturk, N.; Xandri-Monje, H.; Pala, Z.; Abraham, K.; de Gomes, A.R.G.J.; Orman, M.N.; et al. Sex-, Feeding-, and Circadian Time-Dependency of P-Glycoprotein Expression and Activity-Implications for Mechanistic Pharmacokinetics Modeling. Sci. Rep. 2019, 9, 10505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Huang, Q.; Hu, X.; Zhang, S.; Jiang, Y.; Yao, G.; Hu, K.; Xu, X.; Liang, B.; Wu, Q.; et al. Disrupting Circadian Rhythm via the PER1-HK2 Axis Reverses Trastuzumab Resistance in Gastric Cancer. Cancer Res. 2022, 82, 1503–1517. [Google Scholar] [CrossRef] [PubMed]
- Shiromizu, S.; Yamauchi, T.; Kusunose, N.; Matsunaga, N.; Koyanagi, S.; Ohdo, S. Dosing Time-Dependent Changes in the Anti-Tumor Effect of XCT Inhibitor Erastin in Human Breast Cancer Xenograft Mice. Biol. Pharm. Bull. 2019, 42, 1921–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, J.; Martinelli, J.; Aboumanify, O.; Ballesta, A.; Relógio, A. A Mathematical Model of the Circadian Clock and Drug Pharmacology to Optimize Irinotecan Administration Timing in Colorectal Cancer. Comput. Struct. Biotechnol. J. 2021, 19, 5170–5183. [Google Scholar] [CrossRef] [PubMed]
- Cederroth, C.R.; Albrecht, U.; Bass, J.; Brown, S.A.; Dyhrfjeld-Johnsen, J.; Gachon, F.; Green, C.B.; Hastings, M.H.; Helfrich-Förster, C.; Hogenesch, J.B.; et al. Medicine in the Fourth Dimension. Cell Metab. 2019, 30, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.J.W.; Innominato, P.F.; Lévi, F.; Ballesta, A. Optimizing Circadian Drug Infusion Schedules towards Personalized Cancer Chronotherapy. PLoS Comput. Biol. 2020, 16, e1007218. [Google Scholar] [CrossRef] [Green Version]
- Innominato, P.F.; Ballesta, A.; Huang, Q.; Focan, C.; Chollet, P.; Karaboué, A.; Giacchetti, S.; Bouchahda, M.; Adam, R.; Garufi, C.; et al. Sex-Dependent Least Toxic Timing of Irinotecan Combined with Chronomodulated Chemotherapy for Metastatic Colorectal Cancer: Randomized Multicenter EORTC 05011 Trial. Cancer Med. 2020, 9, 4148–4159. [Google Scholar] [CrossRef] [Green Version]
- Marjot, T.; Ray, D.W.; Tomlinson, J.W. Is It Time for Chronopharmacology in NASH? J. Hepatol. 2022, 76, 1215–1224. [Google Scholar] [CrossRef]
- Hesse, J.; Malhan, D.; Yalҫin, M.; Aboumanify, O.; Basti, A.; Relógio, A. An Optimal Time for Treatment—Predicting Circadian Time by Machine Learning and Mathematical Modelling. Cancers 2020, 12, 3103. [Google Scholar] [CrossRef]
- Philip, A.K.; Philip, B. Chronopharmaceuticals: Hype or Future of Pharmaceutics. Curr. Pharm. Des. 2011, 17, 1512–1516. [Google Scholar] [CrossRef] [PubMed]
- Taranalli, S.S.; Dandagi, P.M.; Mastiholimath, V.S. Development of Hollow/Porous Floating Beads of Metoprolol for Pulsatile Drug Delivery. Eur. J. Drug Metab. Pharm. 2015, 40, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, A.Z.; Zyad, H.; Mohammed, H.; Ihsan, K.; Alrawi, L.; Abdullah, M.; Akram, O. Recent Advances in Remotely Controlled Pulsatile Drug Delivery Systems. J. Adv. Pharm. Technol. Res. 2022, 13, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Klerman, E.B.; Rahman, S.A.; St Hilaire, M.A. What Time Is It? A Tale of Three Clocks, with Implications for Personalized Medicine. J. Pineal Res. 2020, 68, e12646. [Google Scholar] [CrossRef] [PubMed]
- Braun, R.; Kath, W.L.; Iwanaszko, M.; Kula-Eversole, E.; Abbott, S.M.; Reid, K.J.; Zee, P.C.; Allada, R. Universal Method for Robust Detection of Circadian State from Gene Expression. Proc. Natl. Acad. Sci. USA 2018, 115, E9247–E9256. [Google Scholar] [CrossRef] [Green Version]
- Wittenbrink, N.; Ananthasubramaniam, B.; Münch, M.; Koller, B.; Maier, B.; Weschke, C.; Bes, F.; de Zeeuw, J.; Nowozin, C.; Wahnschaffe, A.; et al. High-Accuracy Determination of Internal Circadian Time from a Single Blood Sample. J. Clin. Investig. 2018, 128, 3826–3839. [Google Scholar] [CrossRef]
- Wu, G.; Ruben, M.D.; Francey, L.J.; Smith, D.F.; Sherrill, J.D.; Oblong, J.E.; Mills, K.J.; Hogenesch, J.B. A Population-Based Gene Expression Signature of Molecular Clock Phase from a Single Epidermal Sample. Genome Med. 2020, 12, 73. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vandenberghe, A.; Lefranc, M.; Furlan, A. An Overview of the Circadian Clock in the Frame of Chronotherapy: From Bench to Bedside. Pharmaceutics 2022, 14, 1424. https://doi.org/10.3390/pharmaceutics14071424
Vandenberghe A, Lefranc M, Furlan A. An Overview of the Circadian Clock in the Frame of Chronotherapy: From Bench to Bedside. Pharmaceutics. 2022; 14(7):1424. https://doi.org/10.3390/pharmaceutics14071424
Chicago/Turabian StyleVandenberghe, Alan, Marc Lefranc, and Alessandro Furlan. 2022. "An Overview of the Circadian Clock in the Frame of Chronotherapy: From Bench to Bedside" Pharmaceutics 14, no. 7: 1424. https://doi.org/10.3390/pharmaceutics14071424
APA StyleVandenberghe, A., Lefranc, M., & Furlan, A. (2022). An Overview of the Circadian Clock in the Frame of Chronotherapy: From Bench to Bedside. Pharmaceutics, 14(7), 1424. https://doi.org/10.3390/pharmaceutics14071424